Antioxidant and Hypolipidemic Activity of Açai Fruit Makes It a Valuable Functional Food


 ,
,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Cell Selection and Culture
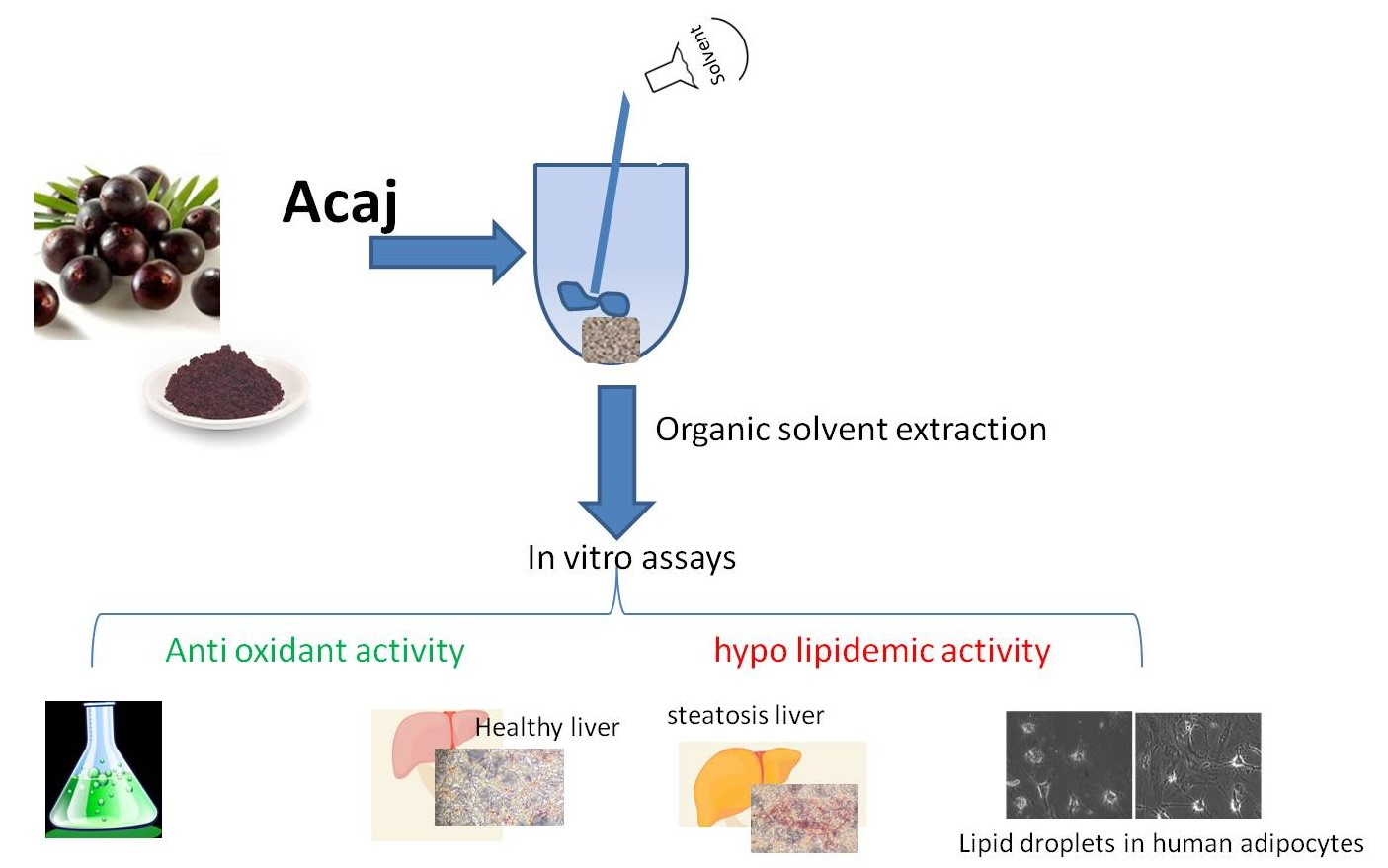
2.3. Açai Extracts Preparation
2.4. HPLC Analyses
2.5. 2′,7′-dihydrodichlorofluorescein Diacetate Assay (DCFDA)
2.6. Cell Viability Assay
2.7. Lipid Peroxidation Assay
2.8. In Vitro Steatosis and Determination of Intracellular Lipid Content
2.9. Fat Deposition in Hypoxic Condition Using Adipocytes In Vitro Model
2.10. Western Blotting Analyses
2.11. Statistical Analysis
3. Results
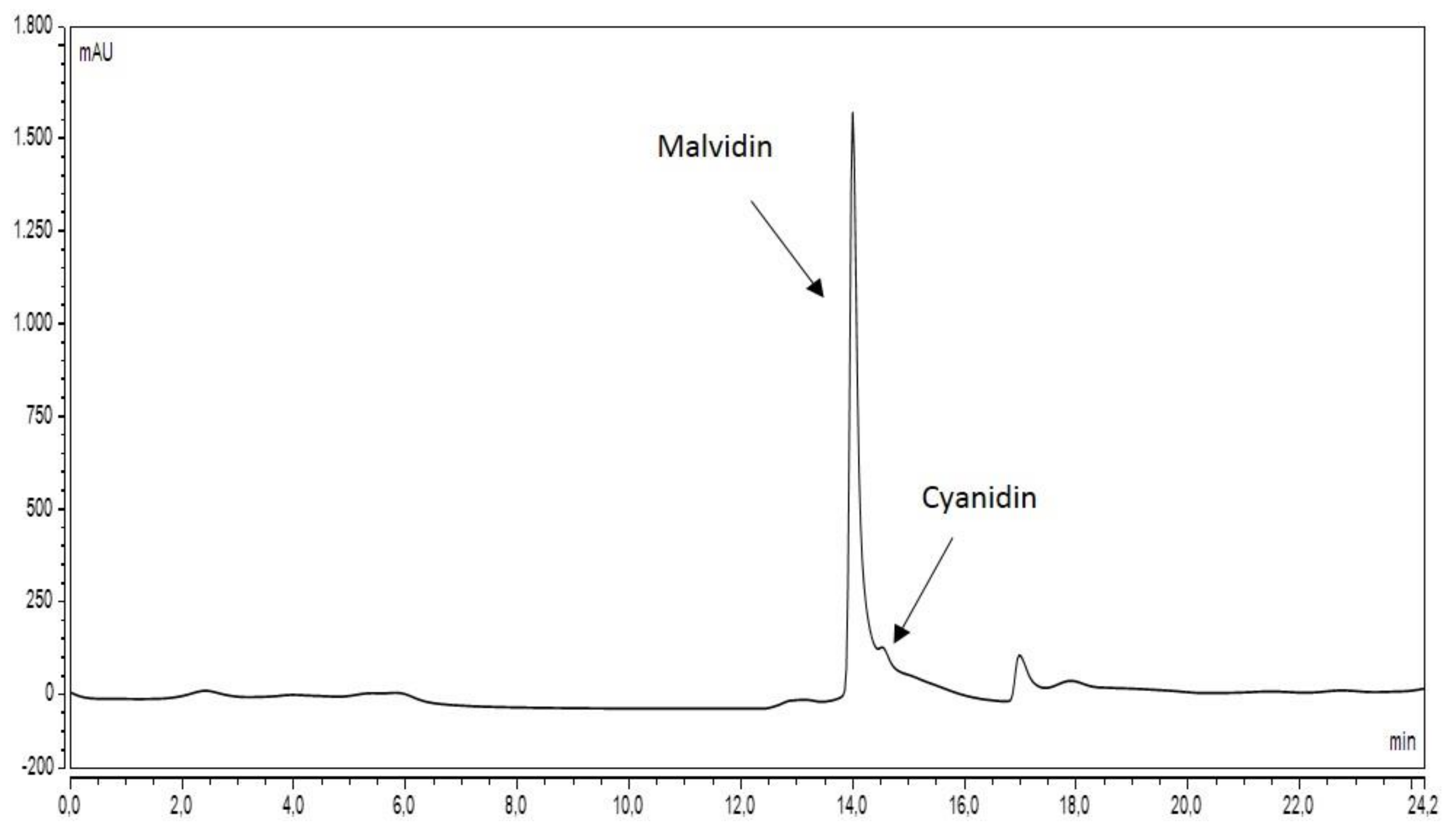
3.1. HPLC Analyses
3.2. Antioxidant Activity of Açai Extract on HaCaT Cells
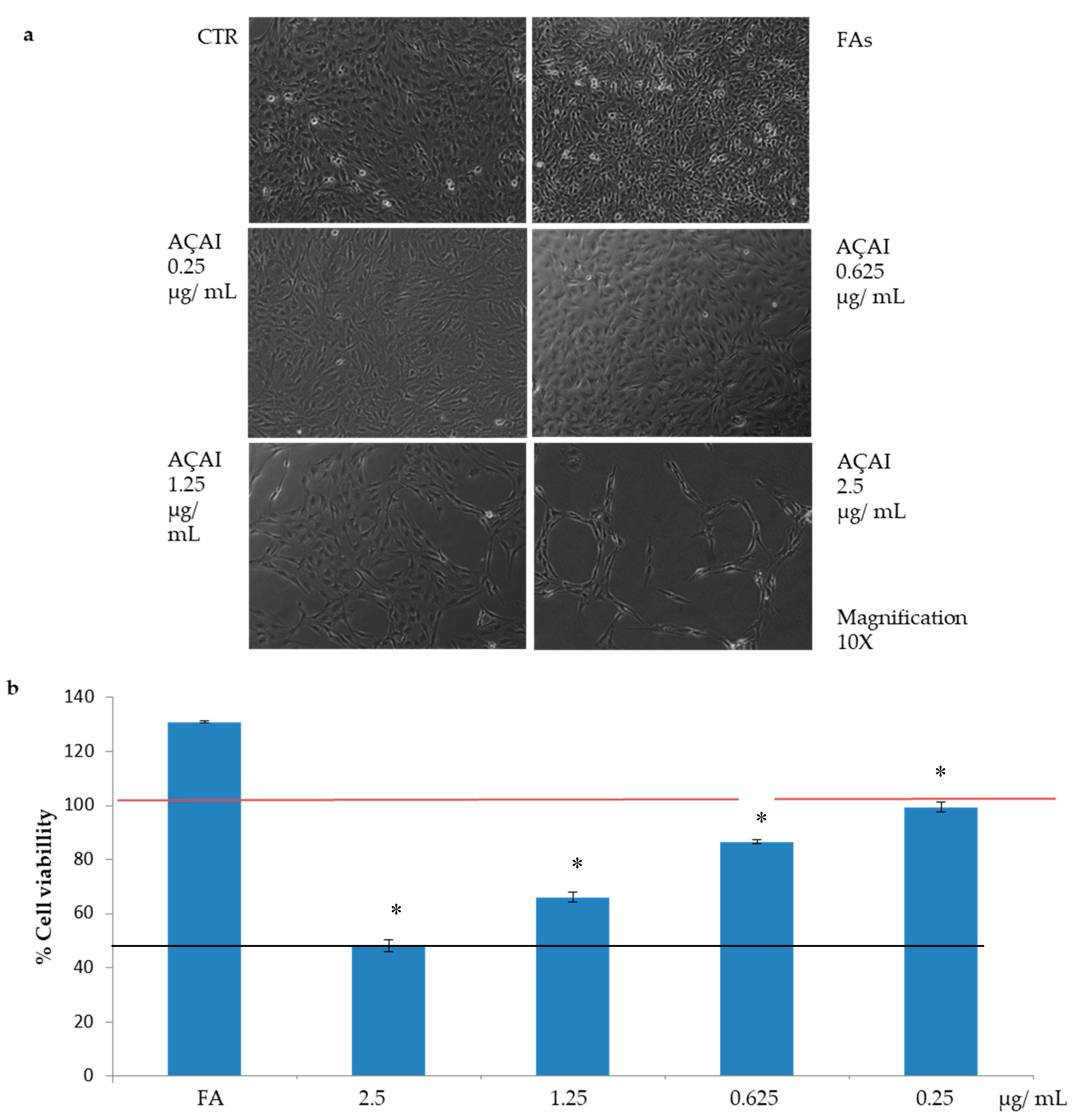
3.2.1. Effects of Açai Extract on HepG2 Cell Viability
3.2.2. Effects of Açai Extract on Lipid Peroxidation in HepG2 Cells
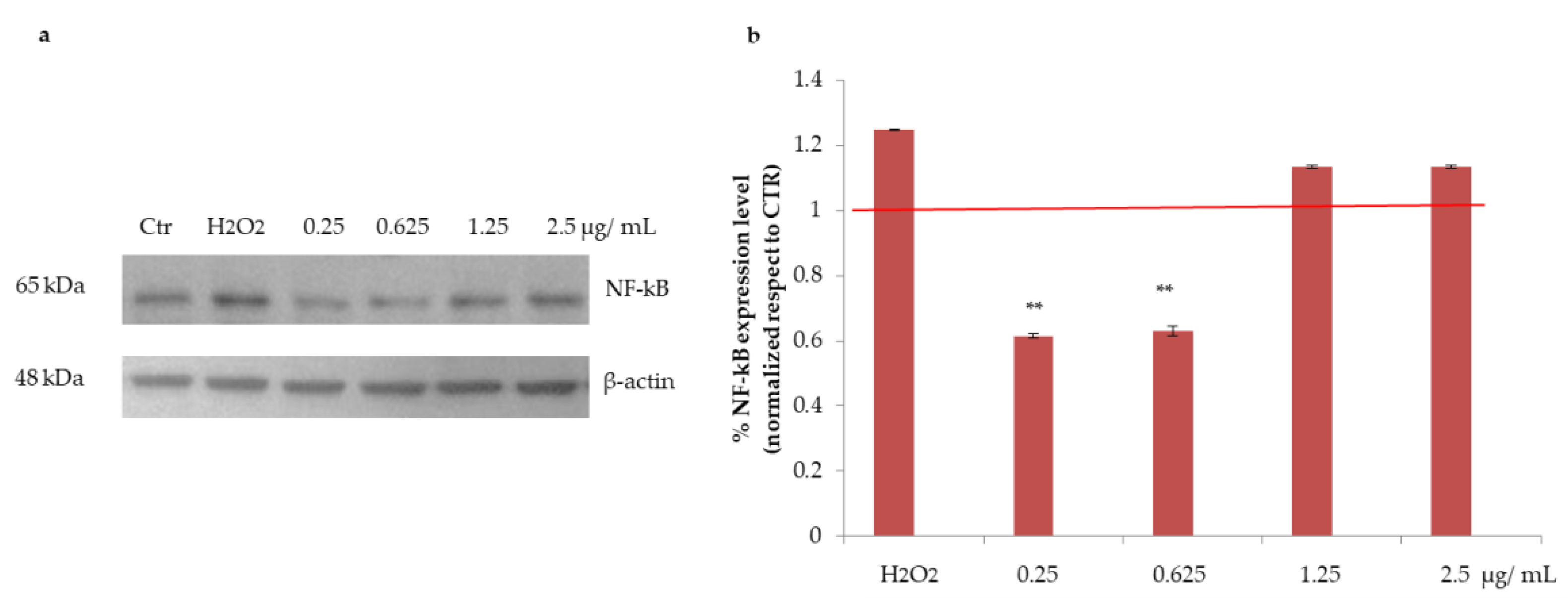
3.2.3. Effects of Açai Extract on NF-kB Protein Expression in HepG2 Cells
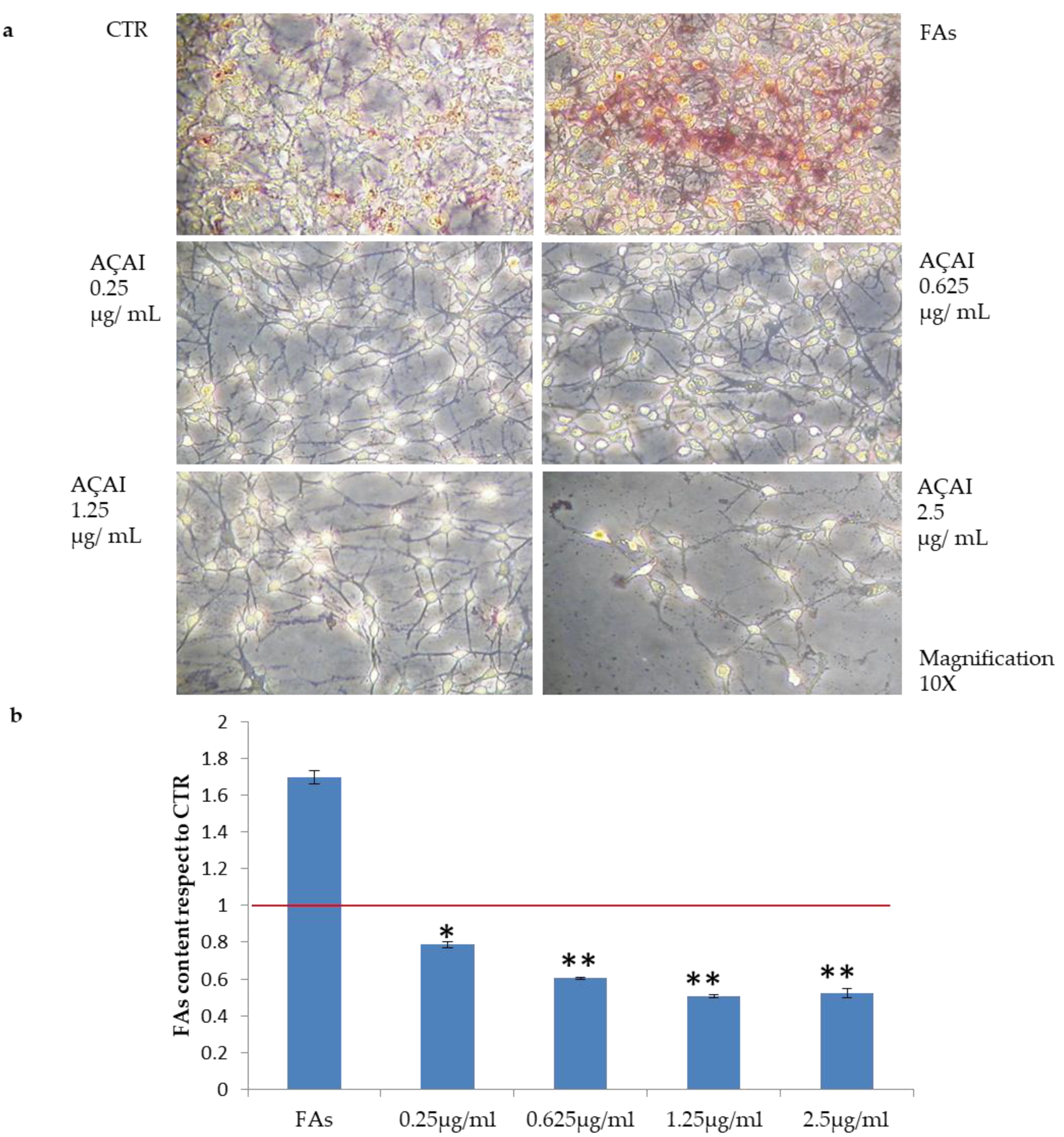
3.2.4. Quali-Quantitative Analyses of Fat Levels in HepG2 and Adipocytes Cells Treated with Açai Extract
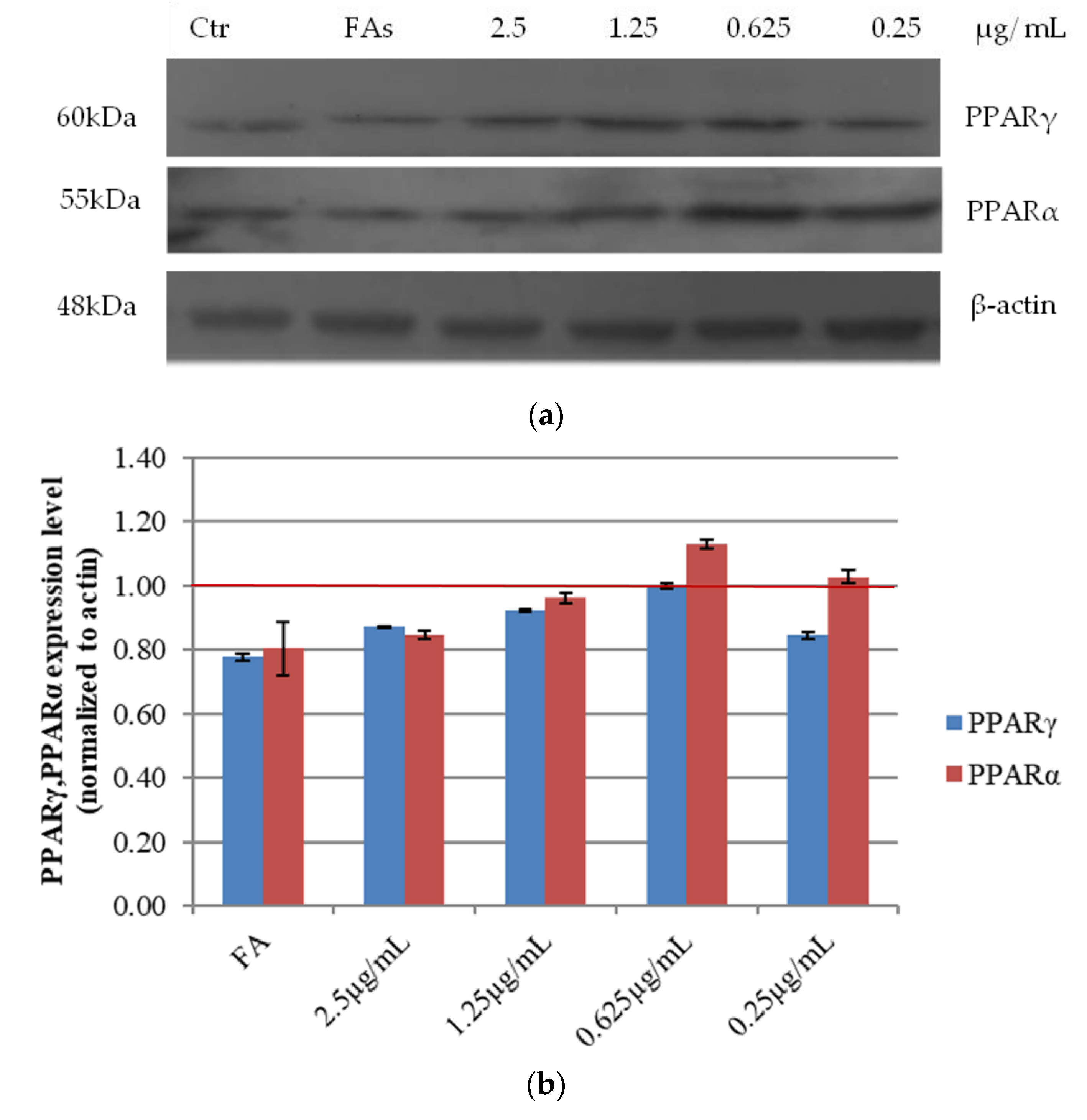
3.2.5. Effects of Açai Extract on PPARs Protein Expression in HepG2 Cells
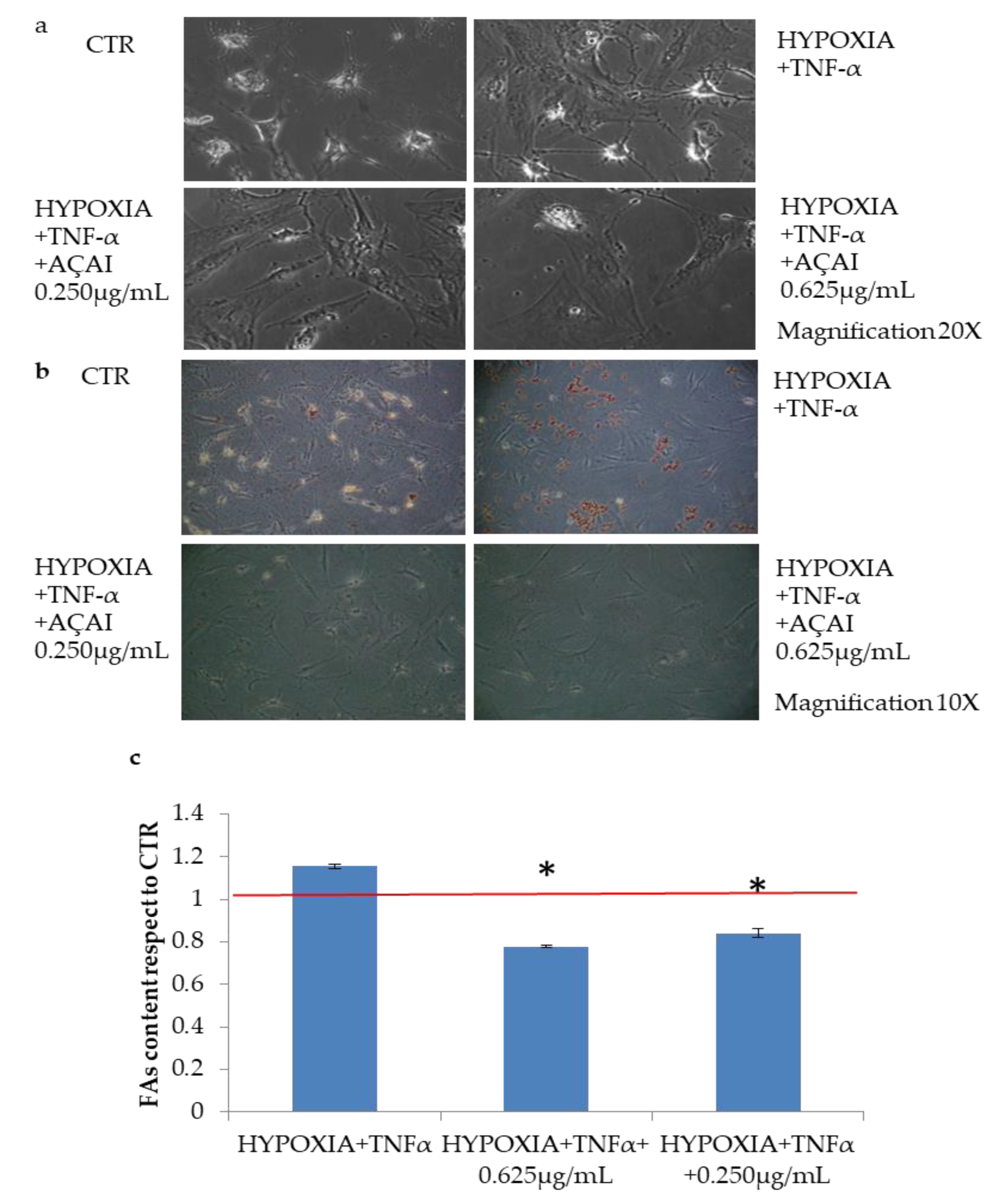
3.2.6. Quali-Quantitative Analyses of Fat Levels in Adipocytes Cells Treated with Açai Extract
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Research Ethics
Conflicts of Interest
References
- Muñiz-Miret, N.V.R.; Hiraoka, M.; Montagnini, F.; Mendelsohn, R.O. The economic value of managing the açaí palm (Euterpe oleracea Mart.) in the floodplains of the Amazon estuary, Pará, Brazil. Forest Ecol. Manag. 1996, 87, 163–173. [Google Scholar] [CrossRef]
- Schauss, A.G.; Wu, X.; Prior, R.L.; Ou, B.; Patel, D.; Huang, D.; Kababick, J.P. Phytochemical and nutrient composition of the freeze-dried amazonian palm berry, Euterpe oleraceae Mart. (açai). J. Agric. Food Chem. 2006, 54, 8598–8603. [Google Scholar] [CrossRef] [PubMed]
- Petruk, G.; Illiano, A.; Del Giudice, R.; Raiola, A.; Amoresano, A.; Rigano, M.M.; Piccoli, R.; Monti, D.M. Malvidin and cyanidin derivatives from açai fruit (Euterpe oleracea Mart.) counteract UV-A-induced oxidative stress in immortalized fibroblasts. J. Photochem. Photobiol. B 2017, 172, 42–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, J.; Xie, C.; Li, Z.; Nagarajan, S.; Schauss, A.G.; Wu, T.; Wu, X. Flavonoids from açai (Euterpe oleracea Mart.) pulp and their antioxidant and anti-inflammatory activities. Food Chem. 2011, 128, 152–157. [Google Scholar] [CrossRef] [PubMed]
- Van De Wier, B.; Koek, G.H.; Bast, A.; Haenen, G.R. The potential of flavonoids in the treatment of non-alcoholic fatty liver disease. Crit. Rev. Food Sci. 2017, 57, 834–855. [Google Scholar] [CrossRef] [PubMed]
- Valenti, L.; Riso, P.; Mazzocchi, A.; Porrini, M.; Fargion, S.; Carlo, A. Dietary anthocyanins as nutritional therapy for nonalcoholic fatty liver disease. Oxid. Med. Cell Longev. 2013, 2013, 145421. [Google Scholar] [CrossRef] [Green Version]
- Heinrich, M.; Dhanji, T.; Casselman, I. Açai (Euterpe oleracea Mart.)—A phytochemical and pharmacological assessment of the species’ health claims. Phytochem. Lett. 2011, 4, 10–21. [Google Scholar] [CrossRef] [Green Version]
- Sudo, R.T.; Neto, M.L.; Monteiro, C.E.; Amaral, R.V.; Resende, Â.C.; Souza, P.J.; Zapata-Sudo, G.; Moura, R.S. Antinociceptive effects of hydroalcoholic extract from Euterpe oleracea Mart. (Açaí) in a rodent model of acute and neuropathic pain. BMC Complement. Altern. Med. 2015, 15, 208. [Google Scholar] [CrossRef] [Green Version]
- Martinez, R.M.; Guimarães, D.A.B.; Berniz, C.R.; Abreu, J.P.; Rocha, A.P.M.D.; Moura, R.S.; Resende, A.C.; Teodoro, A.J. Açai (Euterpe oleracea Mart.) Seed Extract Induces Cell Cycle Arrest and Apoptosis in Human Lung Carcinoma Cells. AJ. Foods 2018, 7, 26. [Google Scholar] [CrossRef] [Green Version]
- de Souza, M.O.; Silva, M.; Silva, M.E.; de Paula Oliveira, R.; Pedrosa, M.L. Diet supplementation with acai (Euterpe oleracea Mart.) pulp improves biomarkers of oxidative stress and the serum lipid profile in rats. Nutrition 2010, 26, 804–810. [Google Scholar] [CrossRef]
- Pereira, R.R.; de Abreu, I.C.; Guerra, J.F.; Lage, N.N.; Lopes, J.M.; Silva, M.; de Lima, W.G.; Silva, M.E.; Pedrosa, M.L. Açai (Euterpe oleracea Mart.) Upregulates Paraoxonase 1 Gene Expression and Activity with Concomitant Reduction of Hepatic Steatosis in High-Fat Diet-Fed Rats. Oxid. Med. Cell Longev. 2016, 2016, 8379105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Oliveira, P.R.; da Costa, C.A.; de Bem, G.F.; Cordeiro, V.S.; Santos, I.B.; de Carvalho, L.C.; da Conceição, E.P.; Lisboa, P.C.; Ognibene, D.T.; Sousa, P.J.; et al. Euterpe oleracea Mart.-Derived Polyphenols Protect Mice from Diet-Induced Obesity and Fatty Liver by Regulating Hepatic Lipogenesis and Cholesterol Excretion. PLoS ONE 2015, 10, e0143721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gironés-Vilaplana, A.; Mena, P.; Moreno, D.A.; García-Viguera, C. Evaluation of sensorial, phytochemical and biological properties of new isotonic beverages enriched with lemon and berries during shelf life. J. Sci. Food Agric. 2014, 94, 1090–1100. [Google Scholar] [CrossRef]
- Neuschwander-Tetri, B.A. and Caldwell, S.H. Nonalcoholic steatohepatitis: Summary of an AASLD Single Topic Conference. Hepatology 2003, 37, 1202–1219. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Sánchez, J.; Chánez-Cárdenas, M.E. The use of cobalt chloride as a chemical hypoxia model. J. Appl. Toxicol. 2019, 39, 556–570. [Google Scholar] [CrossRef]
- Marques, E.S.; Tsuboy, M.S.F.; Carvalho, J.C.T.; Rosa, P.C.P.; Perazzo, F.F.; Gaivão, I.O.M.; Maistro, E.L. First cytotoxic, genotoxic, and antigenotoxic assessment of Euterpe oleracea fruit oil (açaí) in cultured human cells. Genet. Mol. Res. 2017, 16. [Google Scholar] [CrossRef]
- Stellavato, A.; Pirozzi, A.V.A.; de Novellis, F.; Scognamiglio, I.; Vassallo, V.; Giori, A.M.; De Rosa, M.; Schiraldi, C. In vitro assessment of nutraceutical compounds and novel nutraceutical formulations in a liver-steatosis-based model. Lipids Health Dis. 2018, 17, 24. [Google Scholar] [CrossRef] [Green Version]
- Stellavato, A.; La Noce, M.; Corsuto, L.; Pirozzi, A.V.A.; De Rosa, M.; Papaccio, G.; Schiraldi, C.; Tirino, V. Hybrid Complexes of High and Low Molecular Weight Hyaluronans Highly Enhance HASCs Differentiation: Implication for Facial Bioremodelling. Cell Physiol. Biochem. 2017, 44, 1078–1092. [Google Scholar] [CrossRef] [Green Version]
- Pirozzi, A.V.A.; Stellavato, A.; La Gatta, A.; Lamberti, M.; Schiraldi, C. Mancozeb, a fungicide routinely used in agriculture, worsens nonalcoholic fatty liver disease in the human HepG2 cell model. Toxicol. Lett. 2016, 249. [Google Scholar] [CrossRef]
- Stellavato, A.; Lamberti, M.; Pirozzi, A.V.; de Novellis, F.; Schiraldi, C. Myclobutanil worsens nonalcoholic fatty liver disease: An in vitro study of toxicity and apoptosis on HepG2 cells. Toxicol. Lett. 2016, 262, 100–104. [Google Scholar] [CrossRef]
- Grygiel-Górniak, B. Peroxisome proliferator-activated receptors and their ligands: Nutritional and clinical implications. Nutr. J. 2011, 13, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, U.J.; Choi, M.S. Obesity and its metabolic complications: The role of adipokines and the relationship between obesity, inflammation, insulin resistance, dyslipidemia and nonalcoholic fatty liver disease. Int. J. Mol. Sci. 2014, 15, 6184–6223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Penumetcha, M.; Santanam, N. Nutraceuticals as Ligands of PPARγ. PPAR Res. 2012, 2012, 858352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calabrese, E.; Mattson, M.P.; Calabrese, V. Resveratrol commonly displays hormesis: Occurrence and biomedical significance. Hum. Exp. Toxicol. 2010, 29, 980–1015. [Google Scholar] [CrossRef]
- Galiniak, S.; Aebisher, D.; Bartusik-Aebisher, D. Health benefits of resveratrol administration. Acta Biochim. Pol. 2019, 66, 13–21. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Lang, H.; Chen, K.; Zhang, Y.; Gao, Y.; Ran, L.; Yi, L.; Mi, M.; Zhang, Q. Resveratrol protects against nonalcoholic fatty liver disease by improving lipid metabolism and redox homeostasis via the PPARα pathway. Appl. Physiol. Nutr. Metab. 2019. [Google Scholar] [CrossRef]
- Rimando, A.M.; Nagmani, R.; Feller, D.R.; Yokoyama, W. Pterostilbene, a new agonist for the peroxisome proliferator-activated receptor alpha-isoform, lowers plasma lipoproteins and cholesterol in hypercholesterolemic hamsters. J. Agric. Food Chem. 2005, 53, 3403–3407. [Google Scholar] [CrossRef]
- Shahidi, F.; Yeo, J. Bioactivities of Phenolics by Focusing on Suppression of Chronic Diseases: A Review. Int. J. Mol. Sci. 2018, 19, 1573. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Wei, L.; Zhao, C.; Li, J.; Liu, Z.; Zhang, M.; Wang, Y. Resveratrol Maintains Lipid Metabolism Homeostasis via One of the Mechanisms Associated with the Key Circadian Regulator Bmal1. Molecules 2019, 24, 2916. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, M.M.F.; Reis, L.L.T.; Lopes, J.M.M.; Lage, N.N.; Guerra, J.F.D.C.; Zago, H.P.; Bonomo, L.F.; Pereira, R.R.; Lima, W.G.; Silva, M.E.; et al. Açai improves non-alcoholic fatty liver disease (NAFLD) induced by fructose. Nutr. Hosp. 2018, 35, 318–325. [Google Scholar] [CrossRef]
- Baur, J.A.; Pearson, K.J.; Price, N.L.; Jamieson, H.A.; Lerin, C.; Kalra, A.; Pistell, P.J. Resveratrol improves health and survival of mice on a high-calorie diet. Nature 2016, 444, 337–342. [Google Scholar] [CrossRef]
- Baur, J.A.; Sinclair, D.A. Therapeutic potential of resveratrol: The in vivo evidence. Nat. Rev. Drug Discov. 2006, 5, 493–506. [Google Scholar] [CrossRef] [PubMed]
- Pagliassotti, M.J. Endoplasmic reticulum stress in nonalcoholic fatty liver disease. Annu. Rev. Nutr. 2012, 32, 17–33. [Google Scholar] [CrossRef] [PubMed]
- Lagouge, M.; Argmann, C.; Gerhart-Hines, Z.; Meziane, H.; Lerin, C.; Daussin, F.; Geny, B. Resveratrol improves mitochondrial function and protects against metabolic disease by activating SIRT1 and PGC-1α. Cell 2006, 127, 1109–1122. [Google Scholar] [CrossRef] [PubMed]
- Levy, A.; Hasson, P. Correlation between In-vitro and In-vivo Studies based on Pharmacokinetic Considerations. AJBSR 2020, 8, 001236. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time | Storage Conditions | Ethanol | Isopropanol | SA Treatment | ||
|---|---|---|---|---|---|---|
| H2O | PBS | H2O | PBS | |||
| Time 0 | 118 ± 29 | 116 ± 20 | 117 ± 16 | 119 ± 21 | 155 ± 5 * | |
| 72 h | −20 °C | 135 ± 47 | 166 ± 28 | 111 ± 8 a | 121 ± 14 a | 147 ± 1 ** |
| +4 °C | 93 ± 12 a | 108 ± 14 | 123 ± 17 | 82 ± 13 a | ||
| r.t. light | 106 ± 15 | 94 ± 12 a | 106 ± 38 | 89 ± 15 b | ||
| r.t. dark | 130 ± 45 | 98 ± 3 a | 102 ± 7 b | 87 ± 15 b | ||
| 37 °C | 137 ± 2 | 103 ± 31 | 78 ± 18 a | 101 ± 33 | ||
| 7 days | −20 °C | 77 ± 73 | 84 ± 36 a | 136 ± 20 | 93 ± 23 a | 150 ± 8 * |
| +4 °C | 216 ± 99 | 216 ± 37 a | 123 ± 18 | 109 ± 3 b | ||
| r.t. light | 40 ± 44 | 169 ± 31 | 164 ± 8 a | 89 ± 39 | ||
| r.t. dark | 50 ± 57 | 142 ± 23 | 124 ± 4 a | 96 ± 13 a | ||
| 37 °C | 78 ± 39 | 208 ± 53 | 140 ± 36 | 105 ± 23 | ||
| 14 days | −20 °C | 158 ± 18 | 136 ± 11 | 70 ± 7 a | 59 ± 13 a | 147 ± 16 * |
| +4 °C | 114 ± 30 | 116 ± 7 a | 71 ± 2 c | 77 ± 17 a | ||
| r.t. light | 126 ± 15 | 147 ± 10 | 60 ± 37 a | 63 ± 13 c | ||
| r.t. dark | 114 ± 24 | 109 ± 5 b | 67 ± 18 c | 73 ± 9 c | ||
| 37 °C | 143 ± 1 | 94 ± 11 c | 125 ± 7 | 84 ± 19 a | ||
| 21 days | −20 °C | 144 ± 23 | 168 ± 9 c | 151 ± 5 | 156 ± 42 | 140 ± 5 * |
| +4 °C | 57 ± 22 a | 194 ± 33 a | 181 ± 15 c | 170 ± 7 c | ||
| r.t. light | 198 ± 31 | 162 ± 12 c | 161 ± 11 b | 146 ± 14 | ||
| r.t. dark | 114 ± 3 a | 147 ± 10 | 150 ± 4 a | 147 ± 5 | ||
| 37 °C | 159 ± 5 a | 149 ± 30 | 165 ± 30 | 137 ± 18 | ||
| 70 days | −20 °C | 147 ± 14 | 141 ± 1 a | 135 ± 10 | 128 ± 16 | 149 ± 4 * |
| +4 °C | 148 ± 17 | 158 ± 4 a | 149 ± 10 | 130 ± 22 | ||
| r.t. light | 126 ± 4 b | 145 ± 5 | 121 ± 26 | 157 ± 67 | ||
| r.t. dark | 129 ± 6 a | 118 ± 12 a | 133 ± 42 | 138 ± 50 | ||
| 37 °C | ND | ND | ND | ND | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pirozzi, A.V.A.; Imbimbo, P.; D’Agostino, A.; Tirino, V.; Finamore, R.; Monti, D.M.; Piccoli, R.; Schiraldi, C. Antioxidant and Hypolipidemic Activity of Açai Fruit Makes It a Valuable Functional Food. Antioxidants 2021, 10, 40. https://doi.org/10.3390/antiox10010040
Pirozzi AVA, Imbimbo P, D’Agostino A, Tirino V, Finamore R, Monti DM, Piccoli R, Schiraldi C. Antioxidant and Hypolipidemic Activity of Açai Fruit Makes It a Valuable Functional Food. Antioxidants. 2021; 10(1):40. https://doi.org/10.3390/antiox10010040
Chicago/Turabian StylePirozzi, Anna Virginia Adriana, Paola Imbimbo, Antonella D’Agostino, Virginia Tirino, Rosario Finamore, Daria Maria Monti, Renata Piccoli, and Chiara Schiraldi. 2021. "Antioxidant and Hypolipidemic Activity of Açai Fruit Makes It a Valuable Functional Food" Antioxidants 10, no. 1: 40. https://doi.org/10.3390/antiox10010040