
Time Course of Reactive Brain Activities during a Stroop Color-Word Task: Evidence of Specific Facilitation and Interference Effects


Abstract
:1. Introduction
2. Methods
2.1. Participants
2.2. Apparatus and Task Procedure
2.3. Behavioral Performance
2.4. EEG
3. Results
3.1. Behavioral Results
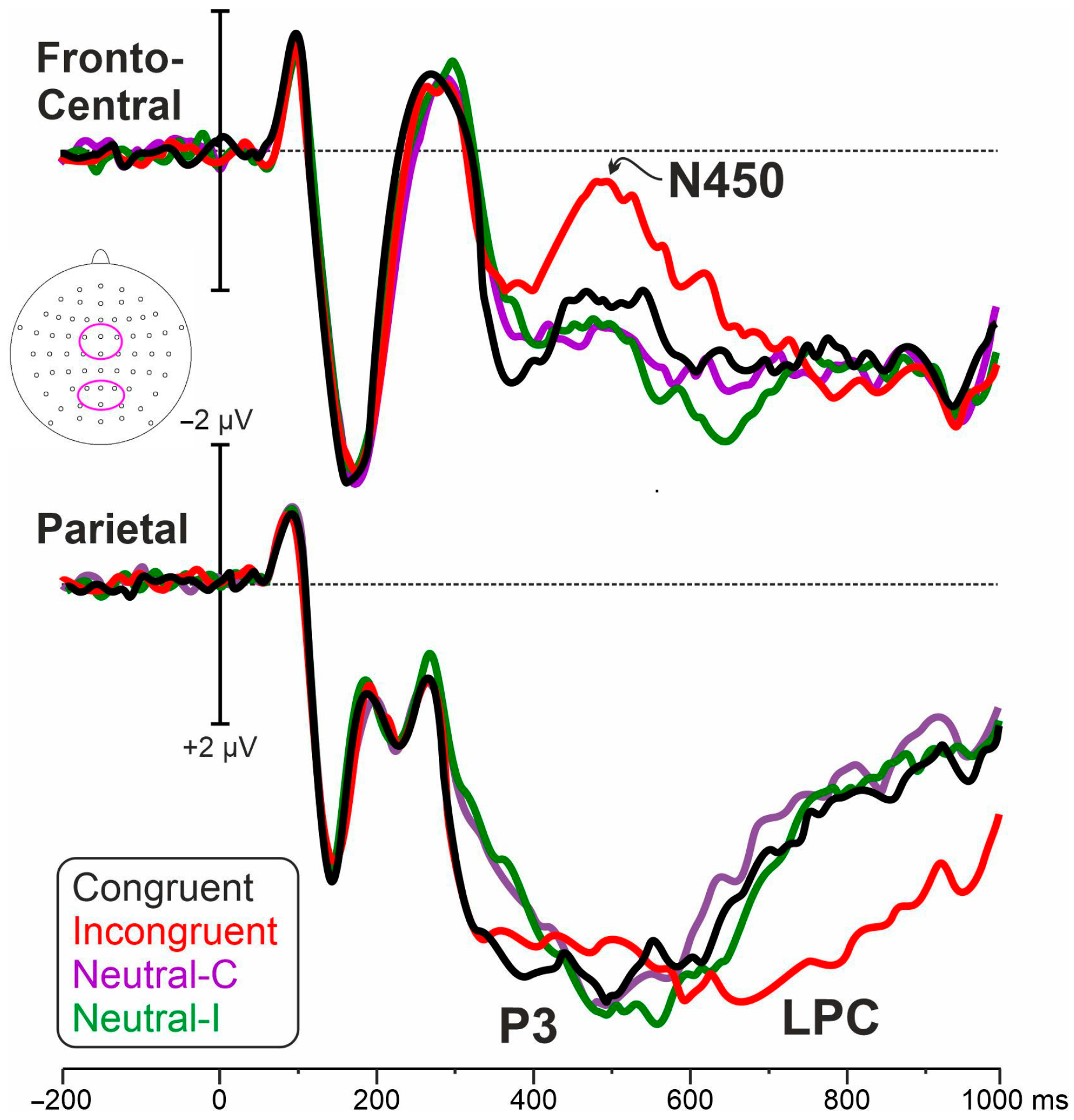
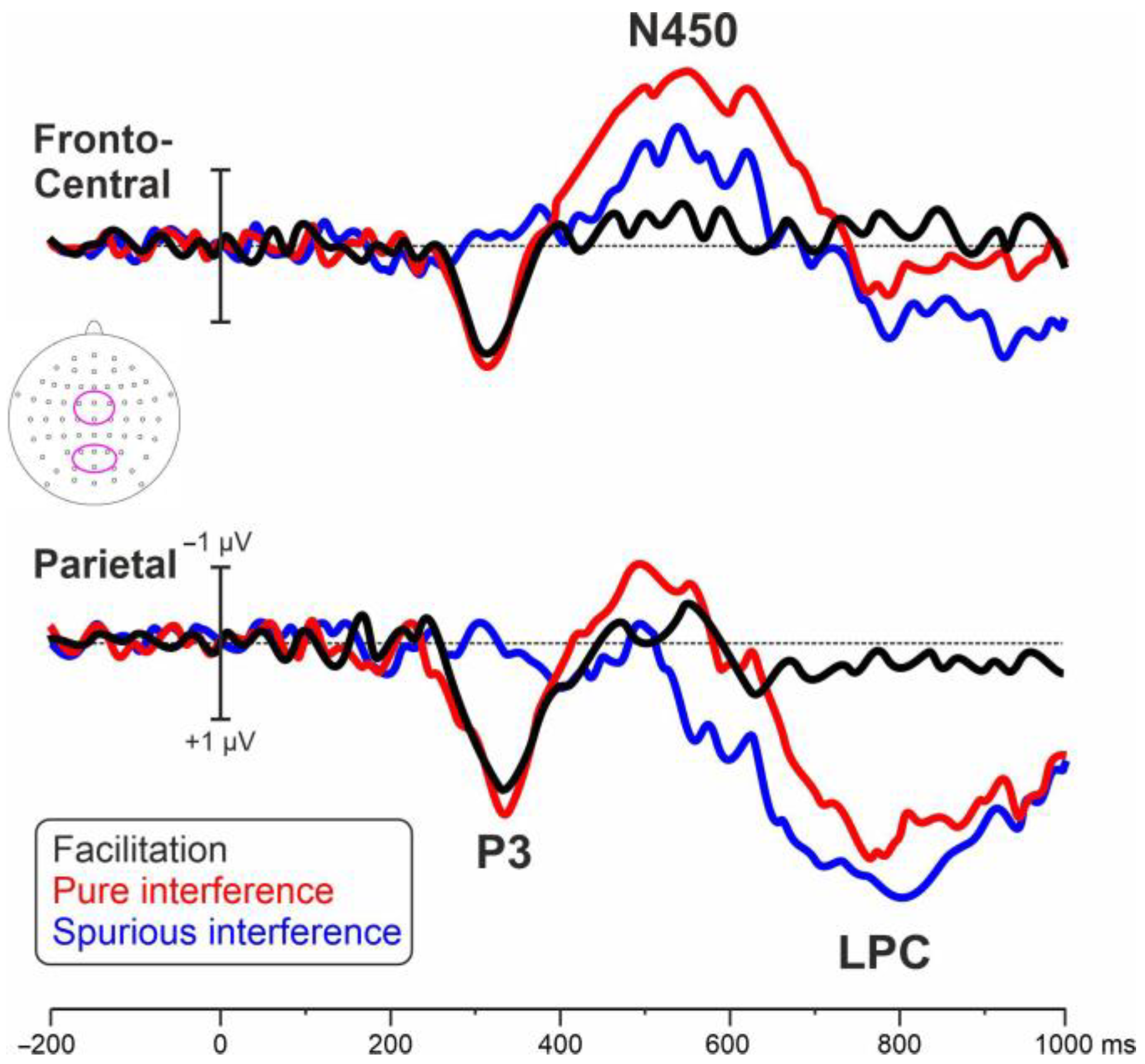
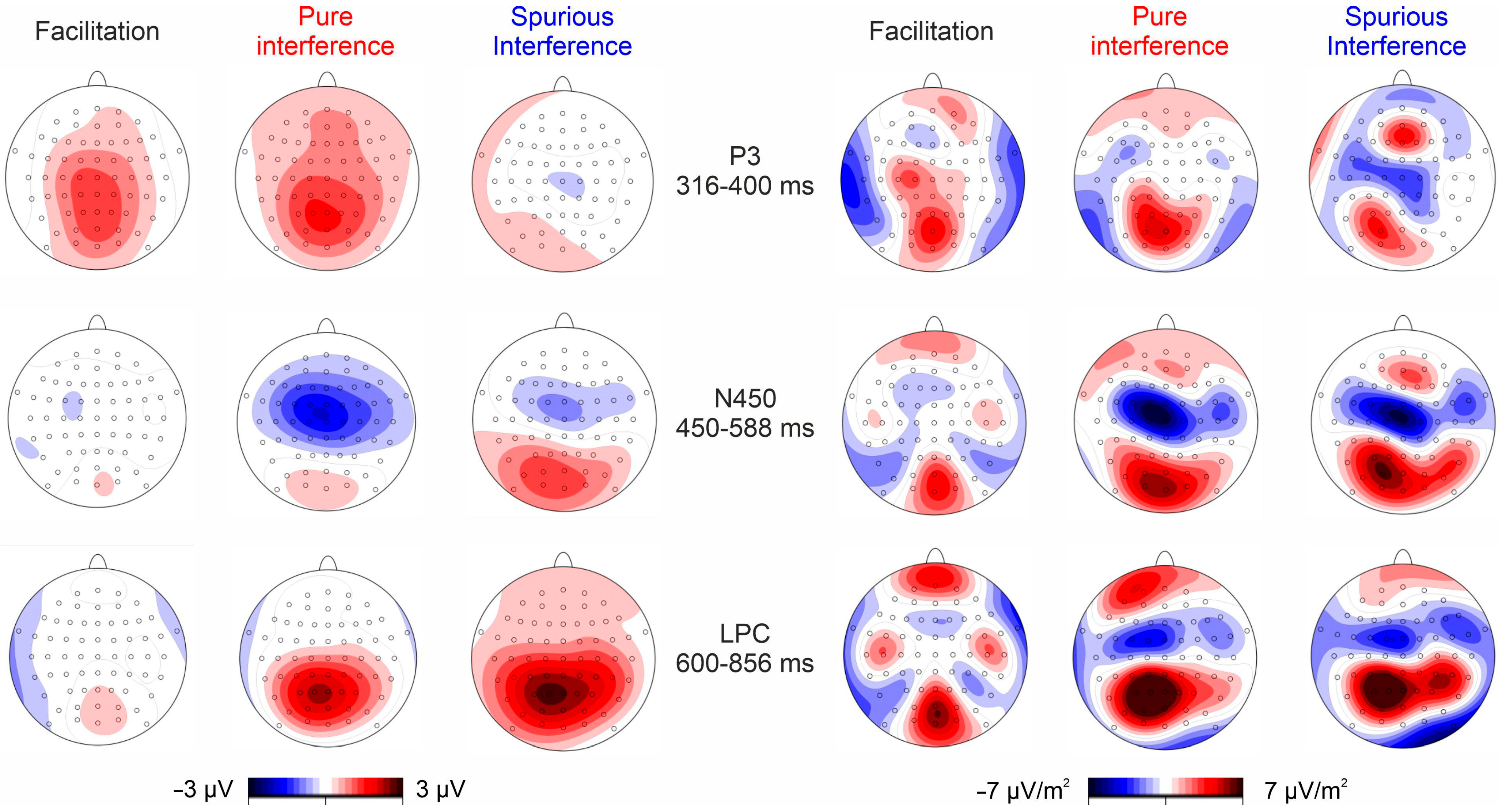
3.2. Electrophysiological Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Miller, J.O.; Low, K. Motor processes in simple, go/no-go, and choice reaction time tasks: A psychophysiological analysis. J. Exp. Psychol. Hum. Percept. Perform. 2001, 27, 266–289. [Google Scholar] [CrossRef]
- Stroop, J.R. Studies of interference in serial verbal reactions. J. Exp. Psychol. 1935, 18, 643. [Google Scholar] [CrossRef]
- Besner, D.; Stolz, J.A.; Boutilier, C. The Stroop effect and the myth of automaticity. Psychon. Bull. Rev. 1997, 4, 221–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Archibald, S.J.; Kerns, K.A. Identification and description of new tests of executive functioning in children. Child Neuropsychol. 1999, 5, 115–129. [Google Scholar] [CrossRef]
- Strauss, E.; Sherman, E.M.; Spreen, O. A Compendium of Neuropsychological Tests: Administration, Norms, and Commentary, 3rd ed.; Oxford University Press: New York, NY, USA, 2006; pp. 493–497. [Google Scholar]
- Epp, A.M.; Dobson, K.S.; Dozois, D.J.; Frewen, P.A. A systematic meta-analysis of the Stroop task in depression. Clin. Psychol. Rev. 2012, 32, 316–328. [Google Scholar] [CrossRef]
- Henik, A.; Salo, R. Schizophrenia and the stroop effect. Behav. Cogn. Neurosci. Rev. 2004, 3, 42–59. [Google Scholar] [CrossRef] [Green Version]
- Stuss, D.T.; Floden, D.; Alexander, M.P.; Levine, B.; Katz, D. Stroop performance in focal lesion patients: Dissociation of processes and frontal lobe lesion location. Neuropsychologia 2001, 39, 771–786. [Google Scholar] [CrossRef]
- Cohen, J.D.; Dunbar, K.; McClelland, J.L. On the control of automatic processes: A parallel distributed processing account of the Stroop effect. Psychol. Rev. 1990, 97, 332. [Google Scholar] [CrossRef] [Green Version]
- Herd, S.A.; Banich, M.T.; O’reilly, R.C. Neural mechanisms of cognitive control: An integrative model of Stroop task performance and fMRI data. J. Cogn. Neurosci. 2006, 18, 22–32. [Google Scholar] [CrossRef]
- Lansbergen, M.M.; Kenemans, J.L. Stroop interference and the timing of selective response activation. Clin. Neurophysiol. 2008, 119, 2247–2254. [Google Scholar] [CrossRef]
- MacLeod, C.M. Half a century of research on the Stroop effect: An integrative review. Psychol. Bull. 1991, 109, 163. [Google Scholar] [CrossRef] [PubMed]
- Ergen, M.; Saban, S.; Kirmizi-Alsan, E.; Uslu, A.; Keskin-Ergen, Y.; Demiralp, T. Time–frequency analysis of the event-related potentials associated with the Stroop test. Int. J. Psychophysiol. 2014, 94, 463–472. [Google Scholar] [CrossRef]
- Aine, C.J.; Harter, M.R. Hemipsheric differences in event-related potentials to Stroop stimuli. Ann. N. Y. Acad. Sci. 1984, 425, 154–156. [Google Scholar] [CrossRef] [PubMed]
- West, R.; Jakubek, K.; Wymbs, N.; Perry, M.; Moore, K. Neural correlates of conflict processing. Exp. Brain Res. 2005, 167, 38–48. [Google Scholar] [CrossRef]
- Mager, R.; Bullinger, A.H.; Brand, S.; Schmidlin, M.; Schärli, H.; Müller-Spahn, F.; Falkenstein, M. Age-related changes in cognitive conflict processing: An event-related potential study. Neurobiol. Aging. 2007, 28, 1925–1935. [Google Scholar] [CrossRef] [PubMed]
- West, R.; Alain, C. Event-related neural activity associated with the Stroop task. Cogn. Brain Res. 1999, 8, 157–164. [Google Scholar] [CrossRef]
- Duncan-Johnson, C.C.; Kopell, B.S. The Stroop effect: Brain potentials localize the source of interference. Science 1981, 214, 938–940. [Google Scholar] [CrossRef]
- Ilan, A.B.; Polich, J. P300 and response time from a manual Stroop task. Clin. Neurophysiol. 1999, 110, 367–373. [Google Scholar]
- Liotti, M.; Woldorff, M.G.; Perez, R.; Mayberg, H.S. An ERP study of the temporal course of the Stroop color-word interference effect. Neuropsychologia 2000, 38, 701–711. [Google Scholar] [CrossRef]
- Parris, B.A.; Wadsley, M.G.; Hasshim, N.; Benattayallah, A.; Augustinova, M.; Ferrand, L. An fMRI study of response and semantic conflict in the stroop task. Front. Psychol. 2019, 10, 2426. [Google Scholar] [CrossRef]
- Polich, J. Updating P300: An integrative theory of P3a and P3b. Clin. Neurophysiol. 2007, 118, 2128–2148. [Google Scholar] [CrossRef] [Green Version]
- Silton, R.L.; Heller, W.; Towers, D.N.; Engels, A.S.; Spielberg, J.M.; Edgar, J.C.; Sass, S.M.; Stewart, J.L.; Sutton, B.P.; Banich, M.T.; et al. The time course of activity in dorsolateral prefrontal cortex and anterior cingulate cortex during top-down attentional control. NeuroImage 2010, 50, 1292–1302. [Google Scholar] [CrossRef]
- D’altrozzo, J.; Wioland, N.; Kotchoubey, B. The N400 and late positive complex (LPC) effects reflect controlled rather than automatic mechanisms of sentence processing. Brain Sci. 2012, 2, 267–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heidlmayr, K.; Kihlstedt, M.; Isel, F. A review on the electroencephalography markers of Stroop executive control processes. Brain Cogn. 2020, 146, 105637. [Google Scholar] [CrossRef] [PubMed]
- Hanslmayr, S.; Pastötter, B.; Bäuml, K.H.; Gruber, S.; Wimber, M.; Klimesch, W. The electrophysiological dynamics of interference during the Stroop task. J. Cogn. Neurosci. 2008, 20, 215–225. [Google Scholar] [CrossRef]
- Coderre, E.; Conklin, K.; van Heuven, W.J. Electrophysiological measures of conflict detection and resolution in the Stroop task. Brain Res. 2011, 1413, 51–59. [Google Scholar] [CrossRef]
- Heidlmayr, K.; Hemforth, B.; Moutier, S.; Isel, F. Neurodynamics of executive control processes in bilinguals: Evidence from ERP and source reconstruction analyses. Front. Psychol. 2015, 6, 821. [Google Scholar] [CrossRef] [Green Version]
- Naylor, L.J.; Stanley, E.M.; Wicha, N.Y. Cognitive and electrophysiological correlates of the bilingual Stroop effect. Front. Psychol. 2012, 3, 81. [Google Scholar] [CrossRef] [Green Version]
- West, R. The effects of aging on controlled attention and conflict processing in the Stroop task. J. Cogn. Neurosci. 2004, 16, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Appelbaum, L.G.; Meyerhoff, K.L.; Woldorff, M.G. Priming and backward influences in the human brain: Processing interactions during the stroop interference effect. Cereb. Cortex 2009, 19, 2508–2521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- West, R. Neural correlates of cognitive control and conflict detection in the Stroop and digit-location tasks. Neuropsychologia 2003, 41, 1122–1135. [Google Scholar] [CrossRef]
- Ehlis, A.C.; Herrmann, M.J.; Wagener, A.; Fallgatter, A.J. Multi-channel near-infrared spectroscopy detects specific inferior-frontal activation during incongruent Stroop trials. Biol. Psychol. 2005, 69, 315–331. [Google Scholar] [CrossRef]
- Burt, J.S. Why do non-color words interfere with color naming? J. Exp. Psychol. Hum. Percep. Perform. 2002, 28, 1019. [Google Scholar] [CrossRef]
- Faul, F.; Erdfelder, E.; Buchner, A.; Lang, A.G. Statistical power analyses using G* Power 3.1: Tests for correlation and regression analyses. Behav. Res. Meth. 2009, 41, 1149–1160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsieh, S.S.; Huang, C.J.; Wu, C.T.; Chang, Y.K.; Hung, T.M. Acute exercise facilitates the N450 inhibition marker and P3 attention marker during stroop test in young and older adults. J. Clin. Med. 2018, 7, 391. [Google Scholar] [CrossRef] [Green Version]
- Veale, J.F. Edinburgh handedness inventory–short form: A revised version based on confirmatory factor analysis. Laterality Asymmetries Body Brain Cogn. 2014, 19, 164–177. [Google Scholar] [CrossRef] [PubMed]
- Jung, T.P.; Makeig, S.; Humphries, C.; Lee, T.W.; Mckeown, M.J.; Iragui, V.; Sejnowski, T.J. Removing electroencephalographic artifacts by blind source separation. Psychophysiology 2000, 37, 163–178. [Google Scholar] [CrossRef] [PubMed]
- Luck, S.J.; Gaspelin, N. How to get statistically significant effects in any ERP experiment (and why you shouldn’t). Psychophysiology 2017, 54, 146–157. [Google Scholar] [CrossRef] [Green Version]
- Badzakova-Trajkov, G.; Barnett, K.J.; Waldie, K.E.; Kirk, I.J. An ERP investigation of the Stroop task: The role of the cingulate in attentional allocation and conflict resolution. Brain Res. 2009, 1253, 139–148. [Google Scholar] [CrossRef]
- Szucs, D.; Soltész, F. Event-related potentials dissociate facilitation and interference effects in the numerical Stroop paradigm. Neuropsychologia 2007, 45, 3190–3202. [Google Scholar] [CrossRef] [PubMed]
- Lindsay, D.S.; Jacoby, L.L. Stroop process dissociations: The relationship between facilitation and interference. J. Exp. Psychol. Hum. Percep. Perform. 1994, 20, 219. [Google Scholar] [CrossRef] [PubMed]
- Macleod, C.M. Training on integrated versus separated Stroop tasks: The progression of interference and facilitation. Mem. Cogn. 1998, 26, 201–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacLeod, C.M.; MacDonald, P.A. Interdimensional interference in the Stroop effect: Uncovering the cognitive and neural anatomy of attention. Trends Cogn. Sci. 2000, 4, 383–391. [Google Scholar] [CrossRef]
- Botvinick, M.M.; Braver, T.S.; Barch, D.M.; Carter, C.S.; Cohen, J.D. Conflict monitoring and cognitive control. Psychol. Rev. 2001, 108, 624. [Google Scholar] [CrossRef] [PubMed]
- Brown, T.L. The Relationship Between Stroop Interference and Facilitation Effects: Statistical Artifacts, Baselines, and a Reassessment. J. Exp. Psychol. Hum. Percep. Perform. 2011, 37, 85–99. [Google Scholar] [CrossRef]
- Atkinson, C.M.; Drysdale, K.; Fulham, W.R. Event-related potentials to Stroop and reverse Stroop stimuli. Int. J. Psychophysiol. 2003, 47, 1–21. [Google Scholar] [CrossRef]
- Gonzalez-Rosa, J.J.; Inuggi, A.; Blasi, V.; Cursi, M.; Annovazzi, P.; Comi, G.; Falini, A.; Leocani, L. Response competition and response inhibition during different choice-discrimination tasks: Evidence from ERP measured inside MRI scanner. Int. J. Psychophysiol. 2013, 89, 37–47. [Google Scholar] [CrossRef]
- Mecklinger, A.; Ullsperger, P. P3 varies with stimulus categorization rather than probability. Electroencephalogr. Clin. Neurophysiol. 1993, 86, 395–407. [Google Scholar] [CrossRef]
- Kok, A. On the utility of P3 amplitude as a measure of processing capacity. Psychophysiology 2001, 38, 557–577. [Google Scholar] [CrossRef] [Green Version]
- Soltani, M.; Knight, R.T. Neural origins of the P300. Crit. Rev. Neurobiol. 2000, 14, 199–224. [Google Scholar] [CrossRef] [Green Version]
- Kopp, B.; Mattler, U.; Rist, F. Selective attention and response competition in schizophrenic patients. Psychiatry Res. 1994, 53, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Nieuwenhuis, S.; Yeung, N.; Van Den Wildenberg, W.; Ridderinkhof, K.R. Electrophysiological correlates of anterior cingulate function in a go/no-go task: Effects of response conflict and trial type frequency. Cogn. Affect. Behav. Neurosci. 2003, 3, 17–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perri, R.L.; Bianco, V.; Facco, E.; Di Russo, F. Now you see one letter, now you see meaningless symbols: Perceptual and semantic hypnotic suggestions reduce Stroop errors through different neurocognitive mechanisms. Front. Neurosci. 2021, 14, 600083. [Google Scholar] [CrossRef] [PubMed]
- Bianco, V.; Berchicci, M.; Mussini, E.; Perri, R.L.; Quinzi, F.; Di Russo, F. Electrophysiological Evidence of Anticipatory Cognitive Control in the Stroop Task. Brain Sci. 2021, 11, 783. [Google Scholar] [CrossRef]
- Bianco, V.; Berchicci, M.; Quinzi, F.; Perri, R.L.; Spinelli, D.; Di Russo, F. Females are more proactive, males are more reactive: Neural basis of the gender-related speed/accuracy trade-off in visuo-motor tasks. Brain Struct. Funct. 2020, 225, 187–201. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| RT ± SD | CE% ± SD | |
|---|---|---|
| Incongruent | 576 ± 57 | 8.4 ± 6.4 |
| Neutral-I | 522 ± 36 | 6.0 ± 4.33 |
| Congruent | 504 ± 26 | 4.1 ± 3.6 |
| Neutral-C | 505 ± 35 | 6.1 ± 4.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Russo, F.; Bianco, V. Time Course of Reactive Brain Activities during a Stroop Color-Word Task: Evidence of Specific Facilitation and Interference Effects. Brain Sci. 2023, 13, 982. https://doi.org/10.3390/brainsci13070982
Di Russo F, Bianco V. Time Course of Reactive Brain Activities during a Stroop Color-Word Task: Evidence of Specific Facilitation and Interference Effects. Brain Sciences. 2023; 13(7):982. https://doi.org/10.3390/brainsci13070982
Chicago/Turabian StyleDi Russo, Francesco, and Valentina Bianco. 2023. "Time Course of Reactive Brain Activities during a Stroop Color-Word Task: Evidence of Specific Facilitation and Interference Effects" Brain Sciences 13, no. 7: 982. https://doi.org/10.3390/brainsci13070982