
Neurodevelopmental Model Explaining Associations between Sex Hormones, Personality, and Eating Pathology
Abstract
:1. Introduction
2. Material and Methods
Literature Search and Criteria
3. Discussion
3.1. Association between Sex Hormones and Eating Pathology
3.1.1. Androgen
Animal Models
Human Models
3.1.2. Ovarian Hormones
Animal Models
Human Models
3.2. Association between Personality and Eating Pathology
3.2.1. Perfectionism
3.2.2. Obsessive-Compulsiveness
3.2.3. Impulsivity
3.2.4. Sensation Seeking
3.3. Association between Sex Hormones and Personality Related to Eating Pathology
3.3.1. Androgen
3.3.2. Ovarian Hormones
3.4. Personality as Mediator or Moderator of Sex Hormones and Eating Pathology
3.5. Neural Representation of Personalities and Corresponding Hormone Receptor Density
3.5.1. Perfectionism
3.5.2. Obsessive-Compulsiveness
3.5.3. Impulsivity
3.5.4. Sensation Seeking
3.6. Brain Sex Hormone Receptor Density, Personality, and ED Symptomology
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Inclusion and Diversity Statement
Acknowledgments
Conflicts of Interest
References
- Beumont, P.J.V. Anorexia nervosa in male subjects. Psychother. Psychosom. 1970, 18, 365–371. [Google Scholar] [CrossRef]
- Culbert, K.M.; Sisk, C.L.; Klump, K.L. A narrative review of sex differences in eating disorders: Is there a biological basis? Clin. Ther. 2021, 43, 95–111. [Google Scholar] [CrossRef] [PubMed]
- Lavender, J.M.; Mitchell, J.E. Eating Disorders and Their Relationship to Impulsivity. Curr. Treat Options Psychiatry 2015, 2, 394–401. [Google Scholar] [CrossRef]
- Farstad, S.M.; McGeown, L.M.; von Ranson, K.M. Eating disorders and personality, 2004–2016: A systematic review and meta-analysis. Clin. Psychol. Rev. 2016, 46, 91–105. [Google Scholar] [CrossRef]
- Gray, J.A. The psychophysiological basis of introversion-extraversion. Behav. Res. Ther. 1970, 8, 249–266. [Google Scholar] [CrossRef]
- McNaughton, N.; Corr, P.J. A two-dimensional neuropsychology of defense: Fear/anxiety and defensive distance. Neurosci. Biobehav. Rev. 2004, 28, 285–305. [Google Scholar] [CrossRef]
- Caseras, X.; Àvila, C.; Torrubia, R. The measurement of individual differences in Behavioural Inhibition and Behavioural Activation Systems: A comparison of personality scales. Personal. Individ. Differ. 2003, 34, 999–1013. [Google Scholar] [CrossRef]
- Newman, J.P.; MacCoon, D.G.; Vaughn, L.J.; Sadeh, N. Validating a distinction between primary and secondary psychopathy with measures of gray’s bis and bas constructs. J. Abnorm. Psychol. 2005, 114, 319–323. [Google Scholar] [CrossRef]
- Määttänen, I.; Jokela, M.; Hintsa, T.; Firtser, S.; Kähönen, M.; Jula, A.; Raitakari, O.T.; Keltikangas-Järvinen, L. Testosterone and temperament traits in men: Longitudinal analysis. Psychoneuroendocrinology 2013, 38, 2243–2248. [Google Scholar] [CrossRef]
- Hines, M. Gender development and the human brain. Ann. Rev. Neurosci. 2011, 34, 69–88. [Google Scholar] [CrossRef] [PubMed]
- Zucker, I. Hormonal determinants of sex differences in saccharin preference, food intake and body weight. Physiol. Behav. 1969, 4, 595–602. [Google Scholar] [CrossRef]
- Bell, D.D.; Zucker, I. Sex differences in body weight and eating: Organization and activation by gonadal hormones in the rat. Physiol. Behav. 1971, 7, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Culbert, K.M.; Sinclair, E.B.; Hildebrandt, B.A.; Klump, K.L.; Sisk, C.L. Perinatal testosterone contributes to mid-to-post pubertal sex differences in risk for binge eating in male and female rats. J. Abnorm. Psychol. 2018, 127, 239–250. [Google Scholar] [CrossRef]
- Phoenix, C.H.; Goy, R.W.; Gerall, A.A.; Young, W.C. Organizing action of prenatally administered testosterone propionate on the tissues mediating mating behavior in the female guinea pig. Endocrinology 1959, 65, 369–382. [Google Scholar] [CrossRef] [PubMed]
- Arnold, A.P. The organizational–activational hypothesis as the foundation for a unified theory of sexual differentiation of all mammalian tissues. Horm. Behav. 2009, 55, 570–578. [Google Scholar] [CrossRef]
- Asarian, L.; Geary, N. Sex differences in the physiology of eating. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2013, 305, R1215–R1267. [Google Scholar] [CrossRef]
- Ryan, B.C.; Vandenbergh, J.G. Intrauterine position effects. Neurosci. Biobehav. Rev. 2002, 26, 665–678. [Google Scholar] [CrossRef]
- Lutchmaya, S.; Baron-Cohen, S.; Raggatt, P.; Knickmeyer, R.; Manning, J.T. 2nd to 4th digit ratios, fetal testosterone and estradiol. Early Hum. Dev. 2004, 77, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Oinonen, K.A.; Bird, J.L. Age at menarche and digit ratio (2D, 4D): Relationships with body dissatisfaction, drive for thinness, and bulimia symptoms in women. Body Image 2012, 9, 302–306. [Google Scholar] [CrossRef] [PubMed]
- Quinton, S.J.; Smith, A.R.; Joiner, T. The 2nd to 4th digit ratio (2D, 4D) and eating disorder diagnosis in women. Personal. Individ. Differ. 2011, 51, 402–405. [Google Scholar] [CrossRef] [PubMed]
- Lydecker, J.A.; Pisetsky, E.M.; Mitchell, K.S.; Thornton, L.M.; Kendler, K.S.; Reichborn-Kjennerud, T.; Lichtenstein, P.; Bulik, C.M.; Mazzeo, S.E. Association between co-twin sex and eating disorders in opposite sex twin pairs: Evaluations in North American, Norwegian, and Swedish samples. J. Psychosom. Res. 2012, 72, 73–77. [Google Scholar] [CrossRef] [PubMed]
- Raevuori, A.; Kaprio, J.; Hoek, H.W.; Sihvola, E.; Rissanen, A.; Keski-Rahkonen, A. Anorexia and bulimia nervosa in same-sex and opposite-sex twins: Lack of association with twin type in a nationwide study of finnish twins. Am. J. Psychiatr. 2008, 165, 1604–1610. [Google Scholar] [CrossRef] [PubMed]
- Mikhail, M.E.; Keel, P.K.; Burt, S.A.; Sisk, C.L.; Neale, M.; Boker, S.; Klump, K.L. Trait negative affect interacts with ovarian hormones to predict risk for emotional eating. Clin. Psychol. Sci. J. Assoc. Psychol Sci. 2021, 9, 114–128. [Google Scholar] [CrossRef] [PubMed]
- Lemaire, A.; Ardaens, K.; Lepretre, J.; Racadot, A.; Buvat-Herbaut, M.; Buvat, J. Gonadal hormones in male anorexia nervosa. Int. J. Eat. Disord. 1983, 2, 135–144. [Google Scholar] [CrossRef]
- Sundblad, C.; Bergman, L.; Eriksson, E. High levels of free testosterone in women with bulimia nervosa. Acta Psychiatr. Scand. 1994, 90, 397–398. [Google Scholar] [CrossRef] [PubMed]
- Culbert, K.M.; Burt, S.A.; Sisk, C.L.; Nigg, J.T.; Klump, K.L. The effects of circulating testosterone and pubertal maturation on risk for disordered eating symptoms in adolescent males. Psychol. Med. 2014, 44, 2271–2286. [Google Scholar] [CrossRef]
- Klump, K.L.; Culbert, K.M.; Sisk, C.L. Sex differences in binge eating: Gonadal hormone effects across development. Annu. Rev. Clin. Psychol. 2017, 13, 183–207. [Google Scholar] [CrossRef] [PubMed]
- Morgan, J.F.; McCluskey, S.E.; Brunton, J.N.; Hubert Lacey, J. Polycystic ovarian morphology and bulimia nervosa: A 9-year follow-up study. Fertil. Steril. 2002, 77, 928–931. [Google Scholar] [CrossRef]
- Culbert, K.M.; Shope, M.M.; Sisk, C.L.; Klump, K.L. Low testosterone is associated with dysregulated eating symptoms in young adult men. Int. J. Eat. Disord. 2020, 53, 1469–1479. [Google Scholar] [CrossRef]
- Klump, K.L.; Gobrogge, K.L.; Perkins, P.S.; Thorne, D.; Sisk, C.L.; Breedlove, S.M. Preliminary evidence that gonadal hormones organize and activate disordered eating. Psychol. Med. 2006, 36, 539–546. [Google Scholar] [CrossRef]
- Bonavera, J.J.; Dube, M.G.; Kalra, P.S.; Kalra, S.P. Anorectic effects of estrogen may be mediated by decreased neuropeptide-Y release in the hypothalamic paraventricular nucleus. Endocrinology 1994, 134, 2367–2370. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Xu, P.; Oyola, M.G.; Xia, Y.; Yan, X.; Saito, K.; Zou, F.; Wang, C.; Yang, Y.; Hinton, A., Jr.; et al. Estrogens stimulate serotonin neurons to inhibit binge-like eating in mice. J. Clin. Invest. 2014, 124, 4351–4362. [Google Scholar] [CrossRef] [PubMed]
- Edler, C.; Lipson, S.F.; Keel, P.K. Ovarian hormones and binge eating in bulimia nervosa. Psychol. Med. 2007, 37, 131–141. [Google Scholar] [CrossRef] [PubMed]
- Klump, K.L.; Suisman, J.L.; Culbert, K.M.; Kashy, D.A.; Keel, P.K.; Sisk, C.L. The effects of ovariectomy on binge eating proneness in adult female rats. Horm. Behav. 2011, 59, 585–593. [Google Scholar] [CrossRef]
- Mystkowski, P.; Seeley, R.J.; Hahn, T.M.; Baskin, D.G.; Havel, P.J.; Matsumoto, A.M.; Wilkinson, C.W.; Peacock-Kinzig, K.; Blake, K.A.; Schwartz, M.W. Hypothalamic melanin-concentrating hormone and estrogen-induced weight loss. J. Neurosci. 2000, 20, 8637–8642. [Google Scholar] [CrossRef] [PubMed]
- Klump, K.L.; Racine, S.E.; Hildebrandt, B.; Burt, S.A.; Neale, M.; Sisk, C.L.; Boker, S.; Keel, P.K. Ovarian hormone influences on dysregulated eating: A comparison of associations in women with versus without binge episodes. Clin. Psychol. Sci. J. Assoc. Psychol. Sci. 2014, 2, 545–559. [Google Scholar] [CrossRef]
- Asarian, L.; Geary, N. Modulation of appetite by gonadal steroid hormones. Philos. Trans. R Soc. B Biol. Sci. 2006, 361, 1251–1263. [Google Scholar] [CrossRef] [PubMed]
- Brown, L.M.; Clegg, D.J. Central effects of estradiol in the regulation of food intake, body weight, and adiposity. J. Steroid Biochem. Mol. Biol. 2010, 122, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Klump, K.L.; O’Connor, S.M.; Hildebrandt, B.A.; Keel, P.K.; Neale, M.; Sisk, C.L.; Boker, S.; Burt, S.A. Differential effects of estrogen and progesterone on genetic and environmental risk for emotional eating in women. Clin. Psychol. Sci. J. Assoc. Psychol. Sci. 2016, 4, 895–908. [Google Scholar] [CrossRef] [PubMed]
- Lerner, L.J. Hormone Antagonists: Inhibitors of Specific Activities of estrogen and Androgen. Recent Prog. Horm. Res. 1964, 20, 435–490. [Google Scholar]
- Baker, J.H.; Runfola, C.D. Eating disorders in midlife women: A perimenopausal eating disorder? Maturitas 2016, 85, 112–116. [Google Scholar] [CrossRef] [PubMed]
- Dandekar, M.P.; Nakhate, K.T.; Kokare, D.M.; Subhedar, N.K. Involvement of CART in estradiol-induced anorexia. Physiol. Behav. 2012, 105, 460–469. [Google Scholar] [CrossRef] [PubMed]
- Rivera, H.M.; Santollo, J.; Nikonova, L.V.; Eckel, L.A. Estradiol increases the anorexia associated with increased 5-HT2C receptor activation in ovariectomized rats. Physiol. Behav. 2012, 105, 188–194. [Google Scholar] [CrossRef]
- Di Bonaventura, M.V.M.; Lutz, T.A.; Romano, A.; Pucci, M.; Geary, N.; Asarian, L.; Cifani, C. Estrogenic suppression of binge-like eating elicited by cyclic food restriction and frustrative-nonreward stress in female rats. Int. J. Eat. Disord. 2017, 50, 624–635. [Google Scholar] [CrossRef]
- Baker, J.H.; Eisenlohr-Moul, T.; Wu, Y.K.; Schiller, C.E.; Bulik, C.M.; Girdler, S.S. Ovarian hormones influence eating disorder symptom variability during the menopause transition: A pilot study. Eat. Behav. 2019, 35, 101337. [Google Scholar] [CrossRef] [PubMed]
- Carmina, E.; Orio, F.; Palomba, S.; Longo, R.A.; Lombardi, G.; Lobo, R.A. Ovarian size and blood flow in women with polycystic ovary syndrome and their correlations with endocrine parameters. Fertil. Steril. 2005, 84, 413–419. [Google Scholar] [CrossRef]
- Eckel, L.A. The ovarian hormone estradiol plays a crucial role in the control of food intake in females. Physiol. Behav. 2011, 104, 517–524. [Google Scholar] [CrossRef]
- Racine, S.E.; Culbert, K.M.; Keel, P.K.; Sisk, C.L.; Alexandra Burt, S.; Klump, K.L. Differential associations between ovarian hormones and disordered eating symptoms across the menstrual cycle in women. Int. J. Eat. Disord. 2012, 45, 333–344. [Google Scholar] [CrossRef]
- Klump, K.L.; Fowler, N.; Mayhall, L.; Sisk, C.L.; Culbert, K.M.; Burt, S.A. Estrogen moderates genetic influences on binge eating during puberty: Disruption of normative processes? J. Abnorm. Psychol. 2018, 127, 458–470. [Google Scholar] [CrossRef]
- Culbert, K.M.; Sisk, C.L.; Klump, K.L. Sex steroid hormones and differential risk for eating pathology: A review of genetic and phenotypic effects across development. Curr. Opin. Behav. Sci. 2018, 23, 124–130. [Google Scholar] [CrossRef]
- Buffenstein, R.; Poppitt, S.D.; McDevitt, R.M.; Prentice, A.M. Food intake and the menstrual cycle: A retrospective analysis, with implications for appetite research. Physiol. Behav. 1995, 58, 1067–1077. [Google Scholar] [CrossRef]
- Gong, E.J.; Garrel, D.; Calloway, D.H. Menstrual cycle and voluntary food intake. Am. J. Clin. Nutr. 1989, 49, 252–258. [Google Scholar] [CrossRef] [PubMed]
- Klump, K.L.; Keel, P.K.; Racine, S.E.; Burt, S.A.; Neale, M.; Sisk, C.L.; Boker, S.; Hu, J.Y. The interactive effects of estrogen and progesterone on changes in emotional eating across the menstrual cycle. J. Abnorm. Psychol. 2013, 122, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Young, J.K. Anorexia nervosa and estrogen: Current status of the hypothesis. Neurosci. Biobehav. Rev. 2010, 34, 1195–1200. [Google Scholar] [CrossRef]
- Golden, N.H.; Shenker, I.R. Amenorrhea in anorexia nervosa. Neuroendocrine control of hypothalamic dysfunction. Int. J. Eat. Disord. 1994, 6, 53–60. [Google Scholar] [CrossRef]
- Cassin, S.E.; von Ranson, K.M. Personality and eating disorders: A decade in review. Clin. Psychol. Rev. 2005, 25, 895–916. [Google Scholar] [CrossRef] [PubMed]
- Frost, R.O.; Marten, P.; Lahart, C.; Rosenblate, R. The dimensions of perfectionism. Cogn. Ther. Res. 1990, 14, 449–468. [Google Scholar] [CrossRef]
- Hewitt, P.L.; Flett, G.L. Perfectionism in the self and social contexts: Conceptualization, assessment, and association with psychopathology. J. Personal. Soc. Psychol. 1991, 60, 456–470. [Google Scholar] [CrossRef]
- Bastiani, A.M.; Rao, R.; Weltzin, T.; Kaye, W.H. Perfectionism in anorexia nervosa. Int. J. Eat. Disord. 1995, 17, 147–152. [Google Scholar] [CrossRef] [PubMed]
- Halmi, K.A.; Sunday, S.R.; Strober, M.; Kaplan, A.; Woodside, D.B.; Fichter, M.; Treasure, J.; Berrettini, W.H.; Kaye, W.H. Perfectionism in anorexia nervosa: Variation by clinical subtype, obsessionality, and pathological eating behavior. Am. J. Psychiatry. 2000, 157, 1799–1805. [Google Scholar] [CrossRef] [PubMed]
- Pratt, E.M.; Telch, C.F.; Labouvie, E.W.; Wilson, G.T.; Agras, W.S. Perfectionism in women with binge eating disorder. Int. J. Eat. Disord. 2001, 29, 177–186. [Google Scholar] [CrossRef]
- Lilenfeld, L.R.R.; Stein, D.; Bulik, C.M.; Strober, M.; Plotnicov, K.; Pollice, C.; Rao, R.; Merikangas, K.R.; Nagy, L.; Kaye, W.H. Personality traits among currently eating disordered, recovered and never ill first-degree female relatives of bulimic and control women. Psychol. Med. 2000, 30, 1399–1410. [Google Scholar] [CrossRef] [PubMed]
- De Zwaan, M.; Mitchell, J.E.; Seim, H.C.; Spercker, S.M.; Pyle, R.L.; Raymond, N.C.; Crosby, R.B. Eating related and general psychopathology in obese females with binge eating disorder. Int. J. Eat. Disord. 1994, 15, 43–52. [Google Scholar] [CrossRef]
- Kehayes, I.L.L.; Smith, M.M.; Sherry, S.B.; Vidovic, V.; Saklofske, D.H. Are perfectionism dimensions risk factors for bulimic symptoms? A meta-analysis of longitudinal studies. Personal. Individ. Differ. 2019, 138, 117–125. [Google Scholar] [CrossRef]
- Halmi, K.A.; Sunday, S.R.; Klump, K.L.; Strober, M.; Leckman, J.F.; Fichter, M.; Kaplan, A.; Woodside, B.; Treasure, J.; Berrettini, W.H.; et al. Obsessions and compulsions in anorexia nervosa subtypes. Int. J. Eat. Disord. 2003, 33, 308–319. [Google Scholar] [CrossRef] [PubMed]
- Rubenstein, C.S.; Pigott, T.A.; L’Heureux, F.; Hill, J.L.; Murphy, D.L. A preliminary investigation of the lifetime prevalence of anorexia and bulimia nervosa in patients with obsessive compulsive disorder. J. Clin. Psychiatry 1992, 53, 309–314. [Google Scholar] [PubMed]
- Jiménez-Murcia, S.; Fernández-Aranda, F.; Raich, R.M.; Alonso, P.; Krug, I.; Jaurrieta, N.; Álvarez-Moya, E.; Labad, J.; Menchón, J.M.; Vallejo, J. Obsessive-compulsive and eating disorders: Comparison of clinical and personality features. Psychiatry Clin. Neurosci. 2007, 61, 385–391. [Google Scholar] [CrossRef]
- Fahy, T.A.; Eisler, I. Impulsivity and eating disorders. Br. J. Psychiatry 1993, 162, 193–197. [Google Scholar] [CrossRef] [PubMed]
- McElroy, S.L.; Phillips, K.A.; Keck, P.E. Obsessive compulsive spectrum disorder. J. Clin. Psychiatry 1994, 55, 33–51, Discussion 52–53. [Google Scholar]
- Claes, L.; Vandereycken, W.; Vertommen, H. Impulsive and compulsive traits in eating disordered patients compared with controls. Personal. Individ. Differ. 2002, 32, 707–714. [Google Scholar] [CrossRef]
- Rossier, V.; Bolognini, M.; Plancherel, B.; Halfon, O. Sensation seeking: A personality trait characteristic of adolescent girls and young women with eating disorders? Eur. Eat. Disord. Rev. 2000, 8, 245–252. [Google Scholar] [CrossRef]
- Brewerton, T.D.; Cotton, B.D.; Kilpatrick, D.G. Sensation seeking, binge-type eating disorders, victimization, and PTSD in the National Women’s Study. Eat. Behav. 2018, 30, 120–124. [Google Scholar] [CrossRef] [PubMed]
- Steiger, H.; Bruce, K.R. Personality traits and disorders associated with anorexia nervosa, bulimia nervosa, and binge eating disorder. In Clinical Handbook of Eating Disorders; CRC Press: Boca Raton, FL, USA, 2004. [Google Scholar]
- Cohen-Bendahan, C.C.C.; Buitelaar, J.K.; van Goozen, S.H.M.; Orlebeke, J.F.; Cohen-Kettenis, P.T. Is there an effect of prenatal testosterone on aggression and other behavioral traits? A study comparing same-sex and opposite-sex twin girls. Horm. Behav. 2005, 47, 230–237. [Google Scholar] [CrossRef]
- Reinisch, J.M. Prenatal exposure to synthetic progestins increases potential for aggression in humans. Science 1981, 211, 1171–1173. [Google Scholar] [CrossRef] [PubMed]
- Gerra, G.; Avanzini, P.; Zaimovic, A.; Sartori, R.; Bocchi, C.; Timpano, M.; Zambelli, U.; Delsignore, R.; Gardini, F.; Talarico, E.; et al. Neurotransmitters, neuroendocrine correlates of sensation-seeking temperament in normal humans. Neuropsychobiology 1999, 39, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Garcia, D.; Lester, N.; Cloninger, K.M.; Robert Cloninger, C. Temperament and Character Inventory (TCI). In Encyclopedia of Personality and Individual Differences; Zeigler-Hill, V., Shackelford, T.K., Eds.; Springer International Publishing: Berlin/Heidelberg, Germany, 2020; pp. 5408–5410. [Google Scholar] [CrossRef]
- Flynn, D.; van Schaik, P.; van Wersch, A. A Comparison of Multi-Item Likert and Visual Analogue Scales for the Assessment of Transactionally Defined Coping Function. Eur. J. Psychol. Assess. 2004, 20, 49–58. [Google Scholar] [CrossRef]
- Mathews, G.A.; Fane, B.A.; Conway, G.S.; Brook, C.G.D.; Hines, M. Personality and congenital adrenal hyperplasia: Possible effects of prenatal androgen exposure. Horm. Behav. 2009, 55, 285–291. [Google Scholar] [CrossRef]
- Daitzman, R.; Zuckerman, M. Disinhibitory sensation seeking, personality and gonadal hormones. Personal. Individ. Differ. 1980, 1, 103–110. [Google Scholar] [CrossRef]
- Zuckerman, M.; Bone, R.N.; Neary, R.; Mangelsdorff, D.; Brustman, B. What is the sensation seeker? Personality trait and experience correlates of the Sensation-Seeking Scales. J. Consult. Clin. Psychol. 1972, 39, 308–321. [Google Scholar] [CrossRef]
- Avgoustinaki, P.D.; Mitsopoulou, E.; Chlouverakis, G.; Triantafillou, T.; Venihaki, M.; Koukouli, S.; Margioris, A.N. Sex steroids and personality traits in the middle luteal phase of healthy normally menstruating young professional women. Hormones 2012, 11, 333–343. [Google Scholar] [CrossRef]
- Hodgson, R.J.; Rachman, S. Obsessional-compulsive complaints. Behav. Res. Ther. 1977, 15, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Goodman, W.K.; Price, L.H.; Rasmussen, S.A.; Mazure, C.; Fleischmann, R.L.; Hill, C.L.; Heninger, G.R.; Charney, D.S. The Yale-Brown Obsessive Compulsive Scale. I. Development, use, and reliability. Arch. Gen. Psychiatry 1989, 46, 1006–1011. [Google Scholar] [CrossRef] [PubMed]
- Hollander, E. Treatment of obsessive-compulsive spectrum disorders with SSRIs. Br. J. Psychiatry Suppl. 1998, 173, 7–12. [Google Scholar] [CrossRef]
- Cyders, M.A.; Smith, G.T. Emotion-based Dispositions to Rash Action: Positive and Negative Urgency. Psychol. Bull. 2008, 134, 807–828. [Google Scholar] [CrossRef]
- Whiteside, S.P.; Lynam, D.R. The Five Factor Model and impulsivity: Using a structural model of personality to understand impulsivity. Personal. Individ. Differ. 2001, 30, 669–689. [Google Scholar] [CrossRef]
- Anestis, M.D.; Smith, A.R.; Fink, E.L.; Joiner, T.E. Dysregulated Eating and Distress: Examining the Specific Role of Negative Urgency in a Clinical Sample. Cogn. Ther. Res. 2009, 33, 390–397. [Google Scholar] [CrossRef]
- Claes, L.; Vandereycken, W.; Vertommen, H. Impulsivity-related traits in eating disorder patients. Personal. Individ. Differ. 2005, 39, 739–749. [Google Scholar] [CrossRef]
- Fischer, S.; Smith, G.T. Binge eating, problem drinking, and pathological gambling: Linking behavior to shared traits and social learning. Personal. Individ. Differ. 2008, 44, 789–800. [Google Scholar] [CrossRef]
- Fischer, S.; Peterson, C.M.; McCarthy, D. A prospective test of the influence of negative urgency and expectancies on binge eating and purging. Psychol. Addict. Behav. J. Soc. Psychol. Addict. Behav. 2013, 27, 294–300. [Google Scholar] [CrossRef]
- Pearson, C.M.; Combs, J.L.; Zapolski, T.C.B.; Smith, G.T. A longitudinal transactional risk model for early eating disorder onset. J. Abnorm. Psychol. 2012, 121, 707–718. [Google Scholar] [CrossRef]
- Pearson, C.M.; Zapolski, T.C.B.; Smith, G.T. A longitudinal test of impulsivity and depression pathways to early binge eating onset. Int. J. Eat. Disord. 2015, 48, 230–237. [Google Scholar] [CrossRef]
- Fossati, A.; Maffei, C.; Bagnato, M.; Battaglia, M.; Donati, D.; Donini, M.; Fiorilli, M.; Novella, L.; Prolo, F. Patterns of covariation of DSM-IV personality disorders in a mixed psychiatric sample. Compr. Psychiatry 2000, 41, 206–215. [Google Scholar] [CrossRef]
- García-Forero, C.; Gallardo-Pujol, D.; Maydeu-Olivares, A.; Andrés-Pueyo, A. Disentangling impulsiveness, aggressiveness and impulsive aggression: An empirical approach using self-report measures. Psychiatry Res. 2009, 168, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Eysenck, H.J.; Eysenck, S.B.G. Manual of the Eysenck Personality Inventory; University of London Press: London, UK, 1964. [Google Scholar]
- Zuckerman, M. Personality in the third dimension: A psychobiological approach. Personal. Individ. Differ. 1989, 10, 391–418. [Google Scholar] [CrossRef]
- Beatty, W.W. Gonadal hormones and sex differences in nonreproductive behaviors in rodents: Organizational and activational influences. Horm. Behav. 1979, 12, 112–163. [Google Scholar] [CrossRef]
- Resnick, S.M.; Gottesman, I.I.; McGue, M. Sensation seeking in opposite-sex twins: An effect of prenatal hormones? Behav. Genet. 1993, 23, 323–329. [Google Scholar] [CrossRef]
- Al-Ayadhi, L.Y. Sex hormones, personality characters and professional status among Saudi females. Saudi Med. J. 2004, 25, 711–716. [Google Scholar] [PubMed]
- Kolin, E.A.; Price, L.; Zoob, I. Development of a sensation-seeking scale. J. Consult. Psychol. 1964, 28, 477–482. [Google Scholar] [CrossRef] [PubMed]
- Racine, S.E.; Keel, P.K.; Burt, S.A.; Sisk, C.L.; Neale, M.; Boker, S.; Klump, K.L. Individual differences in the relationship between ovarian hormones and emotional eating across the menstrual cycle: A role for personality? Eat. Behav. 2013, 14, 161–166. [Google Scholar] [CrossRef]
- Barke, A.; Bode, S.; Dechent, P.; Schmidt-Samoa, C.; Van Heer, C.; Stahl, J. To err is (perfectly) human: Behavioural and neural correlates of error processing and perfectionism. Soc. Cogn. Affect. Neurosci. 2017, 12, 1647–1657. [Google Scholar] [CrossRef] [PubMed]
- Lei, H.; Huang, L.; Li, J.; Liu, W.; Fan, J.; Zhang, X.; Xia, J.; Zhao, K.; Zhu, X.; Rao, H. Altered spontaneous brain activity in obsessive-compulsive personality disorder. Compr. Psychiatry 2020, 96, 152144. [Google Scholar] [CrossRef]
- Abler, B.; Walter, H.; Erk, S.; Kammerer, H.; Spitzer, M. Prediction error as a linear function of reward probability is coded in human nucleus accumbens. NeuroImage 2006, 31, 790–795. [Google Scholar] [CrossRef]
- Buckholtz, J.W.; Treadway, M.T.; Cowan, R.L.; Woodward, N.D.; Li, R.; Ansari, M.S.; Baldwin, R.M.; Schwartzman, A.N.; Shelby, E.S.; Smith, C.E.; et al. Dopaminergic network differences in human impulsivity. Science 2010, 329, 532. [Google Scholar] [CrossRef] [PubMed]
- DeVito, E.E.; Meda, S.A.; Jiantonio, R.; Potenza, M.N.; Krystal, J.H.; Pearlson, G.D. Neural correlates of impulsivity in healthy males and females with family histories of alcoholism. Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol. 2013, 38, 1854–1863. [Google Scholar] [CrossRef] [PubMed]
- Jahanshahi, M.; Obeso, I.; Baunez, C.; Alegre, M.; Krack, P. Parkinson’s disease, the subthalamic nucleus, inhibition, and impulsivity. Mov. Disord. Off. J. Mov. Disord. Soc. 2015, 30, 128–140. [Google Scholar] [CrossRef]
- Brevet-Aeby, C.; Brunelin, J.; Iceta, S.; Padovan, C.; Poulet, E. Prefrontal cortex and impulsivity: Interest of noninvasive brain stimulation. Neurosci. Biobehav. Rev. 2016, 71, 112–134. [Google Scholar] [CrossRef]
- Zhao, J.; Tomasi, D.; Wiers, C.E.; Shokri-Kojori, E.; Demiral, B.; Zhang, Y.; Volkow, N.D.; Wang, G.-J. Correlation between traits of emotion-based impulsivity and intrinsic default-mode network activity. Neural. Plast. 2017, 2017, 9297621. [Google Scholar] [CrossRef] [PubMed]
- Weafer, J.; Crane, N.A.; Gorka, S.M.; Phan, K.L.; de Wit, H. Neural correlates of inhibition and reward are negatively associated. NeuroImage 2019, 196, 188–194. [Google Scholar] [CrossRef] [PubMed]
- Gentili, C.; Vanello, N.; Podina, I.; Popita, R.; Voinescu, B.; Pietrini, P.; David, D. You do not have to act to be impulsive: Brain resting-state activity predicts performance and impulsivity on the Balloon Analogue Risk Task. Behav. Brain Res. 2020, 379, 112395. [Google Scholar] [CrossRef]
- Joseph, J.E.; Liu, X.; Jiang, Y.; Lynam, D.; Kelly, T.H. Neural correlates of emotional reactivity in sensation seeking. Psychol. Sci. 2009, 20, 215–223. [Google Scholar] [CrossRef]
- Wu, D.; Wang, K.; Wei, D.; Chen, Q.; Du, X.; Yang, J.; Qiu, J. Perfectionism mediated the relationship between brain structure variation and negative emotion in a nonclinical sample. Cogn. Affect. Behav. Neurosci. 2017, 17, 211–223. [Google Scholar] [CrossRef] [PubMed]
- Nakao, T.; Okada, K.; Kanba, S. Neurobiological model of obsessive-compulsive disorder: Evidence from recent neuropsychological and neuroimaging findings. Psychiatry Clin. Neurosci. 2014, 68, 587–605. [Google Scholar] [CrossRef]
- Cservenka, A.; Herting, M.M.; Seghete, K.L.M.; Hudson, K.A.; Nagel, B.J. High and low sensation seeking adolescents show distinct patterns of brain activity during reward processing. NeuroImage 2013, 66, 184–193. [Google Scholar] [CrossRef]
- Zuckerman, M. Sensation Seeking: Beyond the Optimal Level of Arousal. L. Erlbaum Associates; Halsted Press Division of Wiley: Hoboken, NJ, USA, 1979. [Google Scholar]
- Tobiansky, D.J.; Korol, A.M.; Ma, C.; Hamden, J.; Jalabert, C.; Tomm, R.J.; Soma, K.K. Testosterone and corticosterone in the mesocorticolimbic system of male rats: Effects of gonadectomy and caloric restriction. Endocrinology 2018, 159, 450–464. [Google Scholar] [CrossRef]
- Tobiansky, D.J.; Wallin-Miller, K.G.; Floresco, S.B.; Wood, R.I.; Soma, K.K. Androgen regulation of the mesocorticolimbic system and executive function. Front. Endocrinol. 2018, 9. Available online: https://www.frontiersin.org/articles/10.3389/fendo.2018.00279 (accessed on 27 March 2023). [CrossRef]
- Simerly, R.B.; Chang, C.; Muramatsu, M.; Swanson, L.W. Distribution of androgen and estrogen receptor mRNA-containing cells in the rat brain: An in situ hybridization study. J. Comp. Neurol. 1990, 294, 76–95. [Google Scholar] [CrossRef] [PubMed]
- Abdelgadir, S.E.; Roselli, C.E.; Choate, J.V.A.; Resko, J.A. Distribution of aromatase cytochrome p450 messenger ribonucleic acid in adult rhesus monkey brains1. Biol. Reprod. 1997, 57, 772–777. [Google Scholar] [CrossRef]
- Finley, S.K.; Kritzer, M.F. Immunoreactivity for intracellular androgen receptors in identified subpopulations of neurons, astrocytes and oligodendrocytes in primate prefrontal cortex. J. Neurobiol. 1999, 40, 446–457. [Google Scholar] [CrossRef]
- Kritzer, M. The distribution of immunoreactivity for intracellular androgen receptors in the cerebral cortex of hormonally intact adult male and female rats: Localization in pyramidal neurons making corticocortical connections. Cereb. Cortex N Y N 1991. 2004, 14, 268–280. [Google Scholar] [CrossRef] [PubMed]
- Fuxjager, M.J.; Forbes-Lorman, R.M.; Coss, D.J.; Auger, C.J.; Auger, A.P.; Marler, C.A. Winning territorial disputes selectively enhances androgen sensitivity in neural pathways related to motivation and social aggression. Proc. Natl. Acad. Sci. USA 2010, 107, 12393–12398. [Google Scholar] [CrossRef] [PubMed]
- Hawrylycz, M.J.; Lein, E.S.; Guillozet-Bongaarts, A.L.; Shen, E.H.; Ng, L.; Miller, J.A.; Van De Lagemaat, L.N.; Smith, K.A.; Ebbert, A.; Riley, Z.L.; et al. An anatomically comprehensive atlas of the adult human brain transcriptome. Nature 2012, 489, 391–399. [Google Scholar] [CrossRef] [PubMed]
- Low, K.L.; Ma, C.; Soma, K.K. Tyramide signal amplification permits immunohistochemical analyses of androgen receptors in the rat prefrontal cortex. J. Histochem. Cytochem. 2017, 65, 295–308. [Google Scholar] [CrossRef] [PubMed]
- Camacho-Arroyo, I.; Mendoza-Rodríguez, C.A.; Mendoza-Garcés, L.; Vanoye-Carlo, A.; Cerbón, M. Progesterone receptor mRNA expression and distribution in the female rabbit brain. J. Steroid. Biochem. Mol. Biol. 2007, 104, 100–104. [Google Scholar] [CrossRef]
- O’Connell, L.A.; Matthews, B.J.; Patel, S.B.; O’Connell, J.D.; Crews, D. Molecular characterization and brain distribution of the progesterone receptor in whiptail lizards. Gen. Comp. Endocrinol. 2011, 171, 64–74. [Google Scholar] [CrossRef]
- Schumacher, M.; Coirini, H.; Robert, F.; Guennoun, R.; El-Etr, M. Genomic and membrane actions of progesterone: Implications for reproductive physiology and behavior. Behav. Brain Res. 1999, 105, 37–52. [Google Scholar] [CrossRef] [PubMed]
- Frye, C.A.; Bayon, L.E.; Pursnani, N.K.; Purdy, R.H. The neurosteroids, progesterone and 3alpha,5alpha-THP, enhance sexual motivation, receptivity, and proceptivity in female rats. Brain Res. 1998, 808, 72–83. [Google Scholar] [CrossRef]
- Frye, C.A.; Petralia, S.M.; Rhodes, M.E. Estrous cycle and sex differences in performance on anxiety tasks coincide with increases in hippocampal progesterone and 3alpha,5alpha-THP. Pharmacol. Biochem. Behav. 2000, 67, 587–596. [Google Scholar] [CrossRef] [PubMed]
- Klump, K.L.; Keel, P.K.; Culbert, K.M.; Edler, C. Ovarian hormones and binge eating: Exploring associations in community samples. Psychol. Med. 2008, 38, 1749–1757. [Google Scholar] [CrossRef] [PubMed]
- Slap, G.B.; Khalid, N.; Paikoff, R.L.; Brooks-Gunn, J.; Warren, M.P. Evolving self-image, pubertal manifestations, and pubertal hormones: Preliminary findings in young adolescent girls. J. Adolesc. Health Off. Publ. Soc. Adolesc. Med. 1994, 15, 327–335. [Google Scholar] [CrossRef]
- Warren, M.P.; Brooks-Gunn, J. Mood and behavior at adolescence: Evidence for hormonal factors. J. Clin. Endocrinol. Metab. 1989, 69, 77–83. [Google Scholar] [CrossRef]
- Watson, H.J.; Yilmaz, Z.; Thornton, L.M.; Hübel, C.; Coleman, J.R.I.; Gaspar, H.A.; Bryois, J.; Hinney, A.; Leppä, V.M.; Mattheisen, M.; et al. Genome-wide association study identifies eight risk loci and implicates metabo-psychiatric origins for anorexia nervosa. Nat. Genet. 2019, 51, 1207–1214. [Google Scholar] [CrossRef] [PubMed]


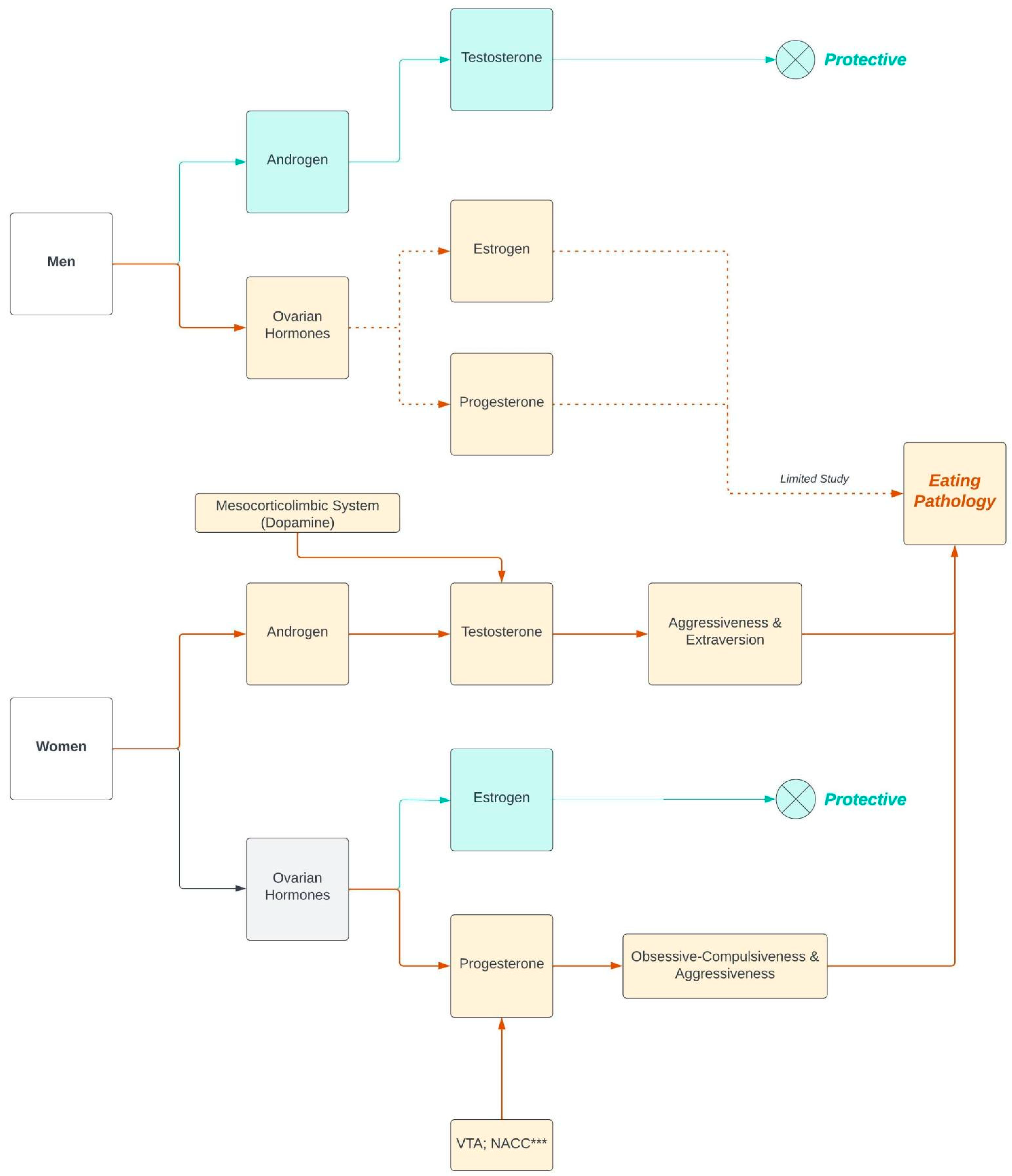{kind=link}
| Anorexic Behaviors (Restrictive) | Association Indicator | Anorexic Behaviors (Binge/Purge) | Association Indicator | Bulimic Behaviors | Association Indicator | Binge Eating | Association Indicator | |
|---|---|---|---|---|---|---|---|---|
| Androgen | [24] | male ↓ | [24] | male ↓ | [25] | female ↑ | [17] | female ↓ |
| [20] | female ↑ | [20] | female ↑ | [22] | male and female; nonsignificant | [22] | male and female; nonsignificant | |
| [22] | male and female; nonsignificant | [22] | male and female; nonsignificant | [20] | female ↓ | [21] | male and female; nonsignificant | |
| [21] | male and female; nonsignificant | [21] | male and female; nonsignificant | [19] | female ↓ | [26] | male ↓ | |
| [21] | male and female; nonsignificant | [27] | male ↓ | |||||
| [28] | female ↑ | [13] | male ↓ | |||||
| [29] | male ↓ | |||||||
| Ovarian Hormones | [30] | female ↑ | [30] | female ↑ | [30] | female ↑ | [30] | female ↑ |
| [31] | male and female rodents; ↑ | [32] | female rodent ↓ | [33] | female; E (↓); PR (↑) | [34] | female rodent ↓ | |
| [35] | male and female rodents; ↑ | [36] | female; E (↓); PR (↑) | [32] | female ↓ | [37] | female ↓ | |
| [38] | ↑ | [39] | female; PR (↑) | [36] | female; E (↓); PR (↑) | [40] | female; E (↓); PR (↑) | |
| [41] | female ↓ | [36] | female; E (↓); PR (↑) | |||||
| [42] | male and female rodents; ↑ | [41] | female ↓ | [39] | female; PR (↑) | [32] | female rodent ↓ | |
| [43] | female rodent; ↑ | [44] | female rodent ↓ | [44] | female rodent ↓ | [41] | female ↓ | |
| [41] | female ↓ | [39] | female; PR (↑) | |||||
| [39] | female; PR (↑) | [44] | female rodent ↓ | |||||
| [44] | female rodent ↓ | [45] | female; E (↑); PR (↑) |
| Restrictive Eating | Association Indicator | Binging/Purging | Bulimic Symptoms | Association Indicator | Binge Eating | Association Indicator | |
|---|---|---|---|---|---|---|---|
| Perfectionism | [59] | male and female; ↑ | [62] | female; ↑ | [63] | female; ↑ | |
| [60] | female; ↑ | [64] | female; ↑ | [61] | female; ↑ | ||
| Obsessive-Compulsiveness | [65] | female; ↑ | [66] | female; ↑ | |||
| [67] | male and female; ↑ | [67] | male and female; ↑ | ||||
| Impulsivity | [68] | male and female; ↑ | [68] | male and female; ↑ | |||
| [69] | female; ↑ | ||||||
| [70] | male and female; ↑ | ||||||
| Sensation Seeking | [71] | male and female; ↓ | [71] | male and female; ↑ | [72] | male and female; ↑ | |
| [72] | male and female; ↑ |
| Hormonal | Scale/Inventory | Association Indicator | Physiological | Scale/Inventory | Association Indicator | |
|---|---|---|---|---|---|---|
| Androgen | [74] | Aggression proneness, Reinisch Aggression Inventory [75] | Females; ↑ | [76] | Novelty Seeking; Temperament and Character Inventory [77]; Visual Analogue Scale [78] | Males; ↑ |
| [79] | Physical aggression; Reinisch Aggression Inventory [75] | Females; ↑ | ||||
| Ovarian Hormones | [80] | 1. Sensation Seeking; 2. Impulsivity; Sensation Seeking Scale [81] | Males; (1–2) ↑ | |||
| [82] | Obsessive-Compulsive behavior; The Minnesota Multiphasic Personality Inventory [82] | Females; P (↑) |
| ED-Related Personalities | Neural Correlates | Association Indicator |
|---|---|---|
| Perfectionism | Anterior Cingulate Cortex [103] | ↑ |
| Obsessive-compulsiveness | Bilateral Caudate; left Insula; left Medial Superior Frontal Gyrus [104] | ↑ |
| Impulsivity | Nucleus Accumbens [105] | ↑ |
| Ventral Striatum [106] | ↑ | |
| left Insula/Inferior Frontal Gyrus [107] | ↑ | |
| Sub-Thalamic Nucleus [108] | ↑ | |
| Prefrontal Cortex [109] | ↑ | |
| Anterior Cingulate Cortex; Medial Prefrontal Cortex [110] | ↑ | |
| Ventral Striatum [111] | ↑ | |
| Anterior Cingulate Cortex; Medial Prefrontal Cortex [112] | ↑ | |
| Sensation seeking | Ventral Tegmental Area [105] | ↑ |
| right Insula; Orbital frontal Cortex [113] | ↑ | |
| NAcc; Insula; Amygdala [113] | ↓ | |
| dorsal Anterior Cingulate Cortex [113] | ↓ |
| Sex Hormone Receptor (Density) | Neural Circuit | Association Indicator |
|---|---|---|
| Androgen | Mesocorticolimbic System (VTA, NAcc, mPFC, OFC) [120] | ↑ |
| Mesocorticolimbic System (VTA, NAcc, mPFC, OFC) [121] | ↑ | |
| Mesocorticolimbic System (VTA, NAcc, mPFC, OFC) [122] | ↑ | |
| Mesocorticolimbic System (VTA, NAcc, mPFC, OFC) [123] | ↑ | |
| Mesocorticolimbic System (VTA, NAcc, mPFC, OFC) [124] | ↑ | |
| Mesocorticolimbic System (VTA, NAcc, mPFC, OFC) [125] | ↑ | |
| Mesocorticolimbic System (VTA, NAcc, mPFC, OFC) [126] | ↑ | |
| Mesocorticolimbic System (VTA, NAcc, mPFC, OFC) [118] | ↑ | |
| Progesterone | Supraoptic area; Anterior Hypothalamic Nuclei; Hippocampus; Cerebral Cortex [127] | ↑ |
| VTA, NAcc [128] | ↑ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Z.; Gobrogge, K. Neurodevelopmental Model Explaining Associations between Sex Hormones, Personality, and Eating Pathology. Brain Sci. 2023, 13, 859. https://doi.org/10.3390/brainsci13060859
Zhao Z, Gobrogge K. Neurodevelopmental Model Explaining Associations between Sex Hormones, Personality, and Eating Pathology. Brain Sciences. 2023; 13(6):859. https://doi.org/10.3390/brainsci13060859
Chicago/Turabian StyleZhao, Ziyu, and Kyle Gobrogge. 2023. "Neurodevelopmental Model Explaining Associations between Sex Hormones, Personality, and Eating Pathology" Brain Sciences 13, no. 6: 859. https://doi.org/10.3390/brainsci13060859