
Phenytoin Decreases Pain-like Behaviors and Improves Opioid Analgesia in a Rat Model of Neuropathic Pain

 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Sciatic Nerve Surgery
2.3. Drug Administration
2.4. Behavioral Tests
2.4.1. Tactile Hypersensitivity (Von Frey Test)
2.4.2. Thermal Hypersensitivity (Cold Plate Test)
2.5. Western Blotting
2.6. Statistical Analysis
3. Results
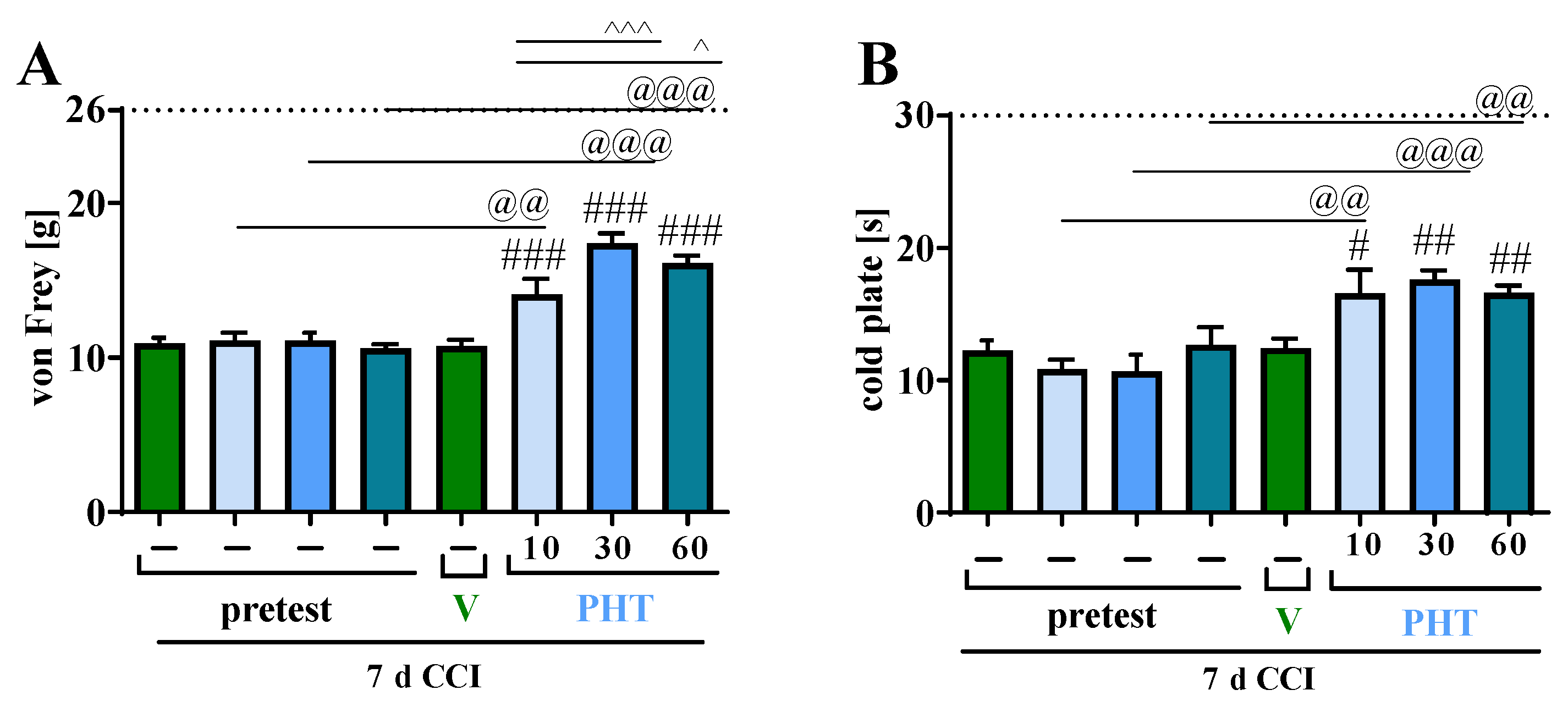
3.1. The Influence of a Single i.p. Phenytoin Administration on Pain-Related Behavior Measured on Day 7 after Chronic Constriction Injury of the Sciatic Nerve in Rats
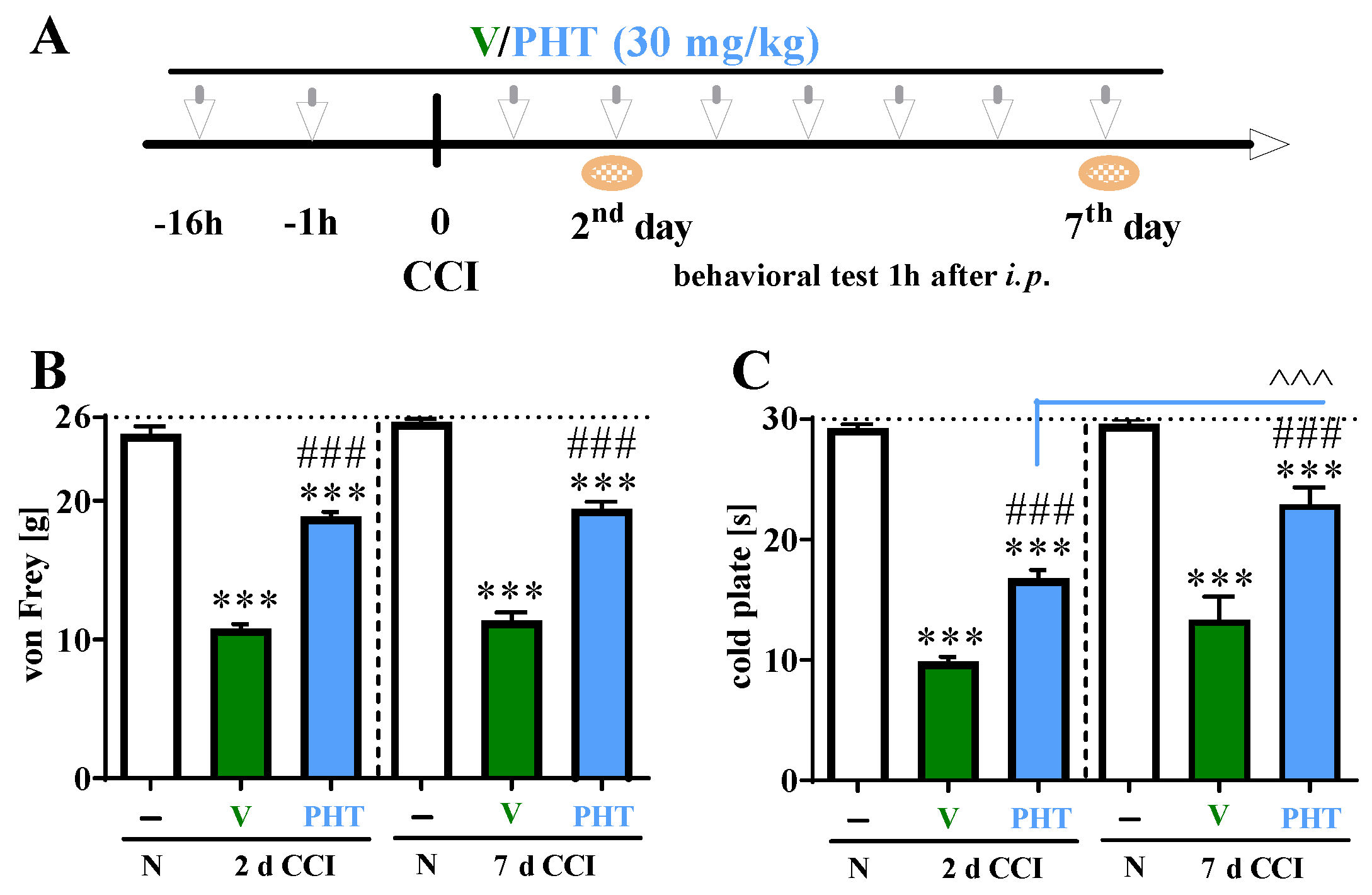
3.2. The Influence of Repeated i.p. Phenytoin Administration on Pain-Related Behavior Measured on Days 2 and 7 after Chronic Constriction Injury of the Sciatic Nerve in Rats
3.3. The Influence of Repeated i.p. Phenytoin Administration on Microglia/Macrophage Marker Measured on Day 7 after Chronic Constriction Injury of the Sciatic Nerve in Rats
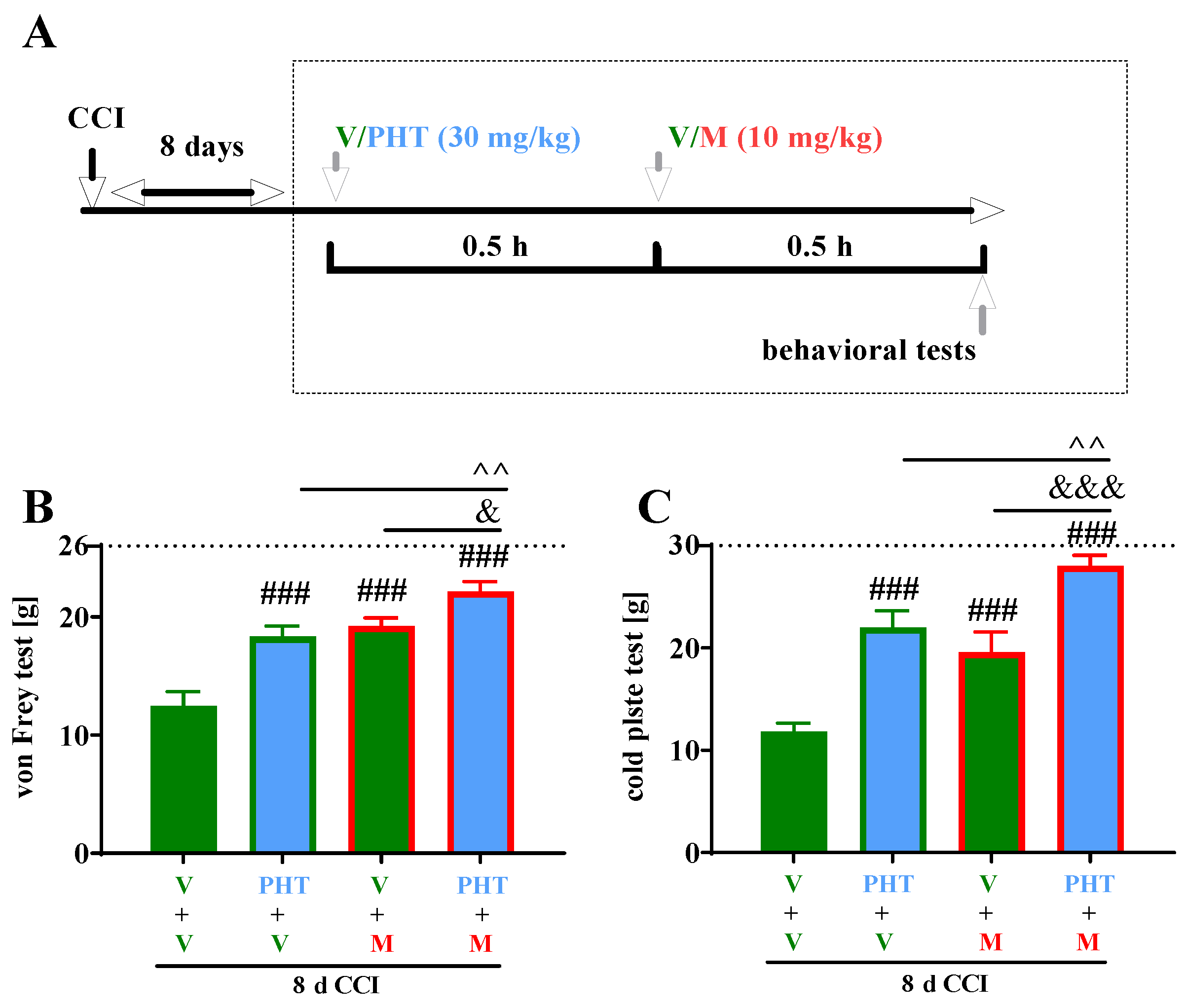
3.4. The Influence of a Single i.p. Administration of Phenytoin on the Efficacy of Morphine on Day 8 after Chronic Constriction Injury of the Sciatic Nerve in Rats
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jensen, T.S.; Baron, R.; Haanpää, M.; Kalso, E.; Loeser, J.D.; Rice, A.S.C.; Treede, R.D. A new definition of neuropathic pain. Pain 2011, 152, 2204–2205. [Google Scholar] [CrossRef] [PubMed]
- Malcangio, M. Role of the immune system in neuropathic pain. Scand. J. Pain 2019, 20, 33–37. [Google Scholar] [CrossRef] [PubMed]
- Finnerup, N.B.; Kuner, R.; Jensen, T.S. Neuropathic Pain: From Mechanisms to Treatment. Physiol. Rev. 2021, 101, 259–301. [Google Scholar] [CrossRef] [PubMed]
- Holbech, J.V.; Jung, A.; Jonsson, T.; Wanning, M.; Bredahl, C.; Bach, F.W. Combination treatment of neuropathic pain: Danish expert recommendations based on a Delphi process. J. Pain Res. 2017, 10, 1467–1475. [Google Scholar] [CrossRef]
- Moisset, X.; Bouhassira, D.; Avez Couturier, J.; Alchaar, H.; Conradi, S.; Delmotte, M.H.; Lanteri-Minet, M.; Lefaucheur, J.P.; Mick, G.; Piano, V.; et al. Pharmacological and non-pharmacological treatments for neuropathic pain: Systematic review and French recommendations. Rev. Neurol. 2020, 176, 325–352. [Google Scholar] [CrossRef]
- Braham, J.; Saja, A. Phenytoin in the treatment of trigeminal and other neuralgias. Lancet 1960, 276, 892–893. [Google Scholar] [CrossRef]
- Rushton, J.G.; Stevens, J.C.; Miller, R.H. Glossopharyngeal (vagoglossopharyngeal) neuralgia: A study of 217 cases. Arch. Neurol. 1981, 38, 201–205. [Google Scholar] [CrossRef]
- Yajnik, S.; Singh, G.P.; Singh, G.; Kumar, M. Phenytoin as a coanalgesic in cancer pain. J. Pain Symptom Manag. 1992, 7, 209–213. [Google Scholar] [CrossRef]
- Chang, V.T. Intravenous phenytoin in the management of crescendo pelvic cancer-related pain. J. Pain Symptom Manag. 1997, 13, 238–240. [Google Scholar] [CrossRef]
- Chadda, V.S.; Mathur, M.S. Double blind study of the effects of diphenylhydantoin sodium on diabetic neuropathy. J. Assoc. Physicians India 1978, 26, 403–406. [Google Scholar]
- Cantor, F.K. Phenytoin treatment of thalamic pain. Br. Med. J. 1972, 4, 590. [Google Scholar] [CrossRef]
- Agnew, D.C.; Goldberg, V.D. A brief trial of phenytoin therapy for thalamic pain. Bull. Los Angel. Neurol. Soc. 1976, 41, 9–12. [Google Scholar]
- Chaturvedi, S.K. Phenytoin in reflex sympathetic dystrophy. Pain 1989, 36, 379–380. [Google Scholar] [CrossRef]
- Schuller, Y.; Linthorst, G.E.; Hollak, C.E.M.; Van Schaik, I.N.; Biegstraaten, M. Pain management strategies for neuropathic pain in Fabry disease—A systematic review. BMC Neurol. 2016, 16, 25. [Google Scholar] [CrossRef]
- McCleane, G.J. Intravenous infusion of phenytoin relieves neuropathic pain: A randomized, double-blinded, placebo-controlled, crossover study. Anesth. Analg. 1999, 89, 985–988. [Google Scholar] [CrossRef]
- Bendtsen, L.; Zakrzewska, J.M.; Abbott, J.; Braschinsky, M.; Di Stefano, G.; Donnet, A.; Eide, P.K.; Leal, P.R.L.; Maarbjerg, S.; May, A.; et al. European Academy of Neurology guideline on trigeminal neuralgia. Eur. J. Neurol. 2019, 26, 831–849. [Google Scholar] [CrossRef]
- Moore, D.; Chong, M.S.; Shetty, A.; Zakrzewska, J.M. A systematic review of rescue analgesic strategies in acute exacerbations of primary trigeminal neuralgia. Br. J. Anaesth. 2019, 123, e385–e396. [Google Scholar] [CrossRef]
- Noro, S.; Seo, Y.; Honjo, K.; Okuma, M.; Asayama, B.; Amano, Y.; Nakamura, H. Intravenous Fosphenytoin Therapy as an Acute Rescue Treatment for Glossopharyngeal Neuralgia Crisis in Patients Awaiting Neurosurgical Procedures: A Case Series. Clin. Neuropharmacol. 2022, 45, 142–144. [Google Scholar] [CrossRef]
- Iorga, A.; Horowitz, B. Phenytoin Toxicity; StatPearls: Treasure Island, FL, USA, 2022. [Google Scholar]
- Thorn, C.F.; Whirl-Carrillo, M.; Leeder, J.S.; Klein, T.E.; Altman, R.B. PharmGKB summary: Phenytoin pathway. Pharmacogenet. Genom. 2012, 22, 466–470. [Google Scholar] [CrossRef]
- Qiao, X.; Sun, G.; Clare, J.J.; Werkman, T.R.; Wadman, W.J. Properties of human brain sodium channel α-subunits expressed in HEK293 cells and their modulation by carbamazepine, phenytoin and lamotrigine. Br. J. Pharmacol. 2014, 171, 1054–1067. [Google Scholar] [CrossRef]
- Yang, M.; Kozminski, D.J.; Wold, L.A.; Modak, R.; Calhoun, J.D.; Isom, L.L.; Brackenbury, W.J. Therapeutic potential for phenytoin: Targeting Na(v)1.5 sodium channels to reduce migration and invasion in metastatic breast cancer. Breast Cancer Res. Treat. 2012, 134, 603–615. [Google Scholar] [CrossRef] [PubMed]
- Nelson, M.; Yang, M.; Dowle, A.A.; Thomas, J.R.; Brackenbury, W.J. The sodium channel-blocking antiepileptic drug phenytoin inhibits breast tumour growth and metastasis. Mol. Cancer 2015, 14, 13. [Google Scholar] [CrossRef] [PubMed]
- Black, J.A.; Liu, S.; Waxman, S.G. Sodium channel activity modulates multiple functions in microglia. Glia 2009, 57, 1072–1081. [Google Scholar] [CrossRef] [PubMed]
- Pappalardo, L.W.; Black, J.A.; Waxman, S.G. Sodium channels in astroglia and microglia. Glia 2016, 64, 1628–1645. [Google Scholar] [CrossRef]
- Chiu, S.Y.; Schrager, P.; Ritchie, J.M. Neuronal-type Na+ and K+ channels in rabbit cultured Schwann cells. Nature 1984, 311, 156–157. [Google Scholar] [CrossRef]
- Black, J.A.; Waxman, S.G. Noncanonical roles of voltage-gated sodium channels. Neuron 2013, 80, 280–291. [Google Scholar] [CrossRef]
- Lo, W.L.; Donermeyer, D.L.; Allen, P.M. A voltage-gated sodium channel is essential for the positive selection of CD4(+) T cells. Nat. Immunol. 2012, 13, 880–887. [Google Scholar] [CrossRef]
- Kis-Toth, K.; Hajdu, P.; Bacskai, I.; Szilagyi, O.; Papp, F.; Szanto, A.; Posta, E.; Gogolak, P.; Panyi, G.; Rajnavolgyi, E. Voltage-gated sodium channel Nav1.7 maintains the membrane potential and regulates the activation and chemokine-induced migration of a monocyte-derived dendritic cell subset. J. Immunol. 2011, 187, 1273–1280. [Google Scholar] [CrossRef]
- Chatelier, A.; Mercier, A.; Tremblier, B.; Thériault, O.; Moubarak, M.; Benamer, N.; Corbi, P.; Bois, P.; Chahine, M.; Faivre, J.F. A distinct de novo expression of Nav1.5 sodium channels in human atrial fibroblasts differentiated into myofibroblasts. J. Physiol. 2012, 590, 4307–4319. [Google Scholar] [CrossRef]
- Black, J.A.; Westenbroek, R.E.; Catterall, W.A.; Waxman, S.G. Type II brain sodium channel expression in non-neuronal cells: Embryonic rat osteoblasts. Mol. Brain Res. 1995, 34, 89–98. [Google Scholar] [CrossRef]
- Zhao, P.; Barr, T.P.; Hou, Q.; Dib-Hajj, S.D.; Black, J.A.; Albrecht, P.J.; Petersen, K.; Eisenberg, E.; Wymer, J.P.; Rice, F.L.; et al. Voltage-gated sodium channel expression in rat and human epidermal keratinocytes: Evidence for a role in pain. Pain 2008, 139, 90–105. [Google Scholar] [CrossRef]
- Wu, W.K.K.; Li, G.R.; Wong, T.M.; Wang, J.Y.; Yu, L.; Cho, C.H. Involvement of voltage-gated K+ and Na+ channels in gastric epithelial cell migration. Mol. Cell. Biochem. 2008, 308, 219–226. [Google Scholar] [CrossRef]
- Patejdl, R.; Leroux, A.C.; Noack, T. Phenytoin inhibits contractions of rat gastrointestinal and portal vein smooth muscle by inhibiting calcium entry. Neurogastroenterol. Motil. 2015, 27, 1453–1465. [Google Scholar] [CrossRef]
- Todorovic, S.M.; Rastogi, A.J.; Jevtovic-Todorovic, V. Potent analgesic effects of anticonvulsants on peripheral thermal nociception in rats. Br. J. Pharmacol. 2003, 140, 255–260. [Google Scholar] [CrossRef]
- Granger, P.; Biton, B.; Faure, C.; Vige, X.; Depoortere, H.; Graham, D.; Langer, S.Z.; Scatton, B.; Avenet, P. Modulation of the γ-aminobutyric acid type A receptor by the antiepileptic drugs carbamazepine and phenytoin. Mol. Pharmacol. 1995, 47, 1189–1196. [Google Scholar]
- Na, J.; Zheng, L.; Wang, L.; Shi, Q.; Yang, Z.; Liu, N.; Guo, Y.; Fan, Y. Phenytoin Regulates Migration and Osteogenic Differentiation by MAPK Pathway in Human Periodontal Ligament Cells. Cell. Mol. Bioeng. 2021, 15, 151–160. [Google Scholar] [CrossRef]
- Okamoto, Y.; Shimizu, K.; Tamura, K.; Miyao, Y.; Yamada, M.; Tsuda, N.; Matsui, Y.; Mogami, H. Effects of phenytoin on cell-mediated immunity. Cancer Immunol. Immunother. 1988, 26, 176–179. [Google Scholar] [CrossRef]
- Andrade-Mena, C.E.; Sardo-Olmedo, J.A.J.; Ramirez-Lizardo, E.J. Effects of phenytoin administration on murine immune function. J. Neuroimmunol. 1994, 50, 3–7. [Google Scholar] [CrossRef]
- Yamada, M.; Ohkawa, M.; Tamura, K.; Mabuchi, E.; Kishima, H.; Tamura, M.; Shimizu, K. Anticonvulsant-induced suppression of IFN-gamma production by lymphocytes obtained from cervical lymph nodes in glioma-bearing mice. J. Neurooncol. 2000, 47, 125–132. [Google Scholar] [CrossRef]
- Craner, M.J.; Damarjian, T.G.; Liu, S.; Hains, B.C.; Lo, A.C.; Black, J.A.; Newcombe, J.; Cuzner, M.L.; Waxman, S.G. Sodium channels contribute to microglia/macrophage activation and function in EAE and MS. Glia 2005, 49, 220–229. [Google Scholar] [CrossRef]
- Black, J.A.; Liu, S.; Hains, B.C.; Saab, C.Y.; Waxman, S.G. Long-term protection of central axons with phenytoin in monophasic and chronic-relapsing EAE. Brain 2006, 129, 3196–3208. [Google Scholar] [CrossRef] [PubMed]
- Kantarci, A.; Augustin, P.; Firatli, E.; Sheff, M.C.; Hasturk, H.; Graves, D.T.; Trackman, P.C. Apoptosis in gingival overgrowth tissues. J. Dent. Res. 2007, 86, 888–892. [Google Scholar] [CrossRef] [PubMed]
- Black, J.A.; Waxman, S.G. Phenytoin protects central axons in experimental autoimmune encephalomyelitis. J. Neurol. Sci. 2008, 274, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, A.M.M.; Yoshimura, A.; Ozaki, Y.; Kaneko, T.; Hara, Y. Cyclosporin A and phenytoin modulate inflammatory responses. J. Dent. Res. 2009, 88, 1131–1136. [Google Scholar] [CrossRef]
- Şimşek, G.; Ciftci, O.; Karadag, N.; Karatas, E.; Kizilay, A. Effects of topical phenytoin on nasal wound healing after mechanical trauma: An experimental study. Laryngoscope 2014, 124, E449–E454. [Google Scholar] [CrossRef]
- Dambach, H.; Hinkerohe, D.; Prochnow, N.; Stienen, M.N.; Moinfar, Z.; Haase, C.G.; Hufnagel, A.; Faustmann, P.M. Glia and epilepsy: Experimental investigation of antiepileptic drugs in an astroglia/microglia co-culture model of inflammation. Epilepsia 2014, 55, 184–192. [Google Scholar] [CrossRef]
- Candotto, V.; Pezzetti, F.; Baj, A.; Beltramini, G.; Lauritano, D.; Di Girolamo, M.; Cura, F. Phenytoin and gingival mucosa: A molecular investigation. Int. J. Immunopathol. Pharmacol. 2019, 33, 1–5. [Google Scholar] [CrossRef]
- Pottoo, F.H.; Salahuddin, M.; Khan, F.A.; Alomar, F.; Dhamen, M.A.A.L.; Alhashim, A.F.; Alqattan, H.H.; Gomaa, M.S.; Alomary, M.N. Thymoquinone Potentiates the Effect of Phenytoin against Electroshock-Induced Convulsions in Rats by Reducing the Hyperactivation of m-TOR Pathway and Neuroinflammation: Evidence from In Vivo, In Vitro and Computational Studies. Pharmaceuticals 2021, 14, 1132. [Google Scholar] [CrossRef]
- Austin, P.J.; Moalem-Taylor, G. The neuro-immune balance in neuropathic pain: Involvement of inflammatory immune cells, immune-like glial cells and cytokines. J. Neuroimmunol. 2010, 229, 26–50. [Google Scholar] [CrossRef]
- Mika, J.; Zychowska, M.; Popiolek-Barczyk, K.; Rojewska, E.; Przewlocka, B. Importance of glial activation in neuropathic pain. Eur. J. Pharmacol. 2013, 716, 106–119. [Google Scholar] [CrossRef]
- Bennett, G.J.; Xie, Y.K. A peripheral mononeuropathy in rat that produces disorders of pain sensation like those seen in man. Pain 1988, 33, 87–107. [Google Scholar] [CrossRef]
- Zajaczkowska, R.; Kwiatkowski, K.; Pawlik, K.; Piotrowska, A.; Rojewska, E.; Makuch, W.; Wordliczek, J.; Mika, J. Metamizole relieves pain by influencing cytokine levels in dorsal root ganglia in a rat model of neuropathic pain. Pharmacol. Rep. 2020, 72, 1310–1322. [Google Scholar] [CrossRef]
- Pawlik, K.; Ciechanowska, A.; Ciapała, K.; Rojewska, E.; Makuch, W.; Mika, J. Blockade of CC Chemokine Receptor Type 3 Diminishes Pain and Enhances Opioid Analgesic Potency in a Model of Neuropathic Pain. Front. Immunol. 2021, 12, 4627. [Google Scholar] [CrossRef]
- Pawlik, K.; Piotrowska, A.; Kwiatkowski, K.; Ciapała, K.; Popiolek-Barczyk, K.; Makuch, W.; Mika, J. The blockade of CC chemokine receptor type 1 influences the level of nociceptive factors and enhances opioid analgesic potency in a rat model of neuropathic pain. Immunology 2020, 159, 413–428. [Google Scholar] [CrossRef]
- Smith, M.D.; Woodhead, J.H.; Handy, L.J.; Pruess, T.H.; Vanegas, F.; Grussendorf, E.; Grussendorf, J.; White, K.; Bulaj, K.K.; Krumin, R.K.; et al. Preclinical Comparison of Mechanistically Different Antiseizure, Antinociceptive, and/or Antidepressant Drugs in a Battery of Rodent Models of Nociceptive and Neuropathic Pain. Neurochem. Res. 2017, 42, 1995–2010. [Google Scholar] [CrossRef]
- Rojewska, E.; Piotrowska, A.; Jurga, A.; Makuch, W.; Mika, J. Zaprinast diminished pain and enhanced opioid analgesia in a rat neuropathic pain model. Eur. J. Pharmacol. 2018, 839, 21–32. [Google Scholar] [CrossRef]
- Jurga, A.M.; Rojewska, E.; Makuch, W.; Mika, J. Lipopolysaccharide from rhodobacter sphaeroides (Tlr4 antagonhypersensitivity and modulates nociceptive factors. Pharm. Biol. 2018, 56, 275–286. [Google Scholar] [CrossRef]
- Jurga, A.M.; Piotrowska, A.; Makuch, W.; Przewlocka, B.; Mika, J. Blockade of P2X4 receptors inhibits neuropathic pain-related behavior by preventing MMP-9 activation and, consequently, pronociceptive interleukin release in a rat model. Front. Pharmacol. 2017, 8, 48. [Google Scholar] [CrossRef]
- Mika, J.; Osikowicz, M.; Makuch, W.; Przewlocka, B. Minocycline and pentoxifylline attenuate allodynia and hyperalgesia and potentiate the effects of morphine in rat and mouse models of neuropathic pain. Eur. J. Pharmacol. 2007, 560, 142–149. [Google Scholar] [CrossRef]
- Makuch, W.; Mika, J.; Rojewska, E.; Zychowska, M.; Przewlocka, B. Effects of selective and non-selective inhibitors of nitric oxide synthase on morphine- and endomorphin-1-induced analgesia in acute and neuropathic pain in rats. Neuropharmacology 2013, 75, 445–457. [Google Scholar] [CrossRef]
- Rojewska, E.; Popiolek-Barczyk, K.; Kolosowska, N.; Piotrowska, A.; Zychowska, M.; Makuch, W.; Przewlocka, B.; Mika, J. PD98059 Influences Immune Factors and Enhances Opioid Analgesia in Model of Neuropathy. PLoS ONE 2015, 10, e0138583. [Google Scholar] [CrossRef]
- Guevara-López, U.; Gutiérrez-Sougarret, B.; López-Pavón, L.; Aldrete, J.A.; Tamayo-Valenzuela, A. Antihyperalgesic activity of chlorimipramine and sodium phenytoin in an induced model of neuropathic pain in rats. Cirugía Cir. 2004, 72, 301–306. [Google Scholar]
- Ko, S.H.; Jochnowitz, N.; Lenkowski, P.W.; Batts, T.W.; Davis, G.C.; Martin, W.J.; Brown, M.L.; Patel, M.K. Reversal of neuropathic pain by alpha-hydroxyphenylamide: A novel sodium channel antagonist. Neuropharmacology 2006, 50, 865–873. [Google Scholar] [CrossRef] [PubMed]
- Takasaki, I.; Nagashima, R.; Ueda, T.; Ogata, T.; Inoue, A.; Shiraki, K.; Kitada, Y.; Arai, S. Fosphenytoin Alleviates Herpes Simplex Virus Infection-Induced Provoked and Spontaneous Pain-Like Behaviors in Mice. Biol. Pharm. Bull. 2022, 45, 360–363. [Google Scholar] [CrossRef] [PubMed]
- Hunskaar, S.; Hole, K. The formalin test in mice: Dissociation between inflammatory and non-inflammatory pain. Pain 1987, 30, 103–114. [Google Scholar] [CrossRef]
- Bornhof, M.; Ihmsen, H.; Schwilden, H.; Yeomans, D.C.; Tzabazis, A. The orofacial formalin test in mice revisited--effects of formalin concentration, age, morphine and analysis method. J. Pain 2011, 12, 633–639. [Google Scholar] [CrossRef]
- De Queiroz, R.B.; De Carvalho, F.L.; Da Fonsêca, D.V.; Barbosa-Filho, J.M.; Salgado, P.R.R.; Paulo, L.L.; De Queiroz, A.B.M.; De Morais Pordeus, L.C.; De Souza, S.A.; Da Silva Souza, H.D.; et al. Antinociceptive effect of hydantoin 3-phenyl-5-(4-ethylphenyl)-imidazolidine-2,4-dione in mice. Molecules 2015, 20, 974–986. [Google Scholar] [CrossRef]
- Sakaue, A.; Honda, M.; Tanabe, M.; Ono, H. Antinociceptive effects of sodium channel-blocking agents on acute pain in mice. J. Pharmacol. Sci. 2004, 95, 181–188. [Google Scholar] [CrossRef]
- Rundfeldt, C.; Loscher, W. Anticonvulsant efficacy and adverse effects of phenytoin during chronic treatment in amygdala-kindled rats. J. Pharmacol. Exp. Ther. 1993, 266, 216–223. [Google Scholar]
- Campero, M.; Baumann, T.K.; Bostock, H.; Ochoa, J.L. Human cutaneous C fibres activated by cooling, heating and menthol. J. Physiol. 2009, 587, 5633–5652. [Google Scholar] [CrossRef]
- Deuis, J.R.; Zimmermann, K.; Romanovsky, A.A.; Possani, L.D.; Cabot, P.J.; Lewis, R.J.; Vetter, I. An animal model of oxaliplatin-induced cold allodynia reveals a crucial role for Nav1.6 in peripheral pain pathways. Pain 2013, 154, 1749–1757. [Google Scholar] [CrossRef]
- Bennett, D.L.; Alex Clark, X.J.; Huang, J.; Waxman, S.G.; Dib-Hajj, S.D. The role of voltage-gated sodium channels in pain signaling. Physiol. Rev. 2019, 99, 1079–1151. [Google Scholar] [CrossRef]
- Obata, K.; Katsura, H.; Mizushima, T.; Yamanaka, H.; Kobayashi, K.; Dai, Y.; Fukuoka, T.; Tokunaga, A.; Tominaga, M.; Noguchi, K. TRPA1 induced in sensory neurons contributes to cold hyperalgesia after inflammation and nerve injury. J. Clin. Investig. 2005, 115, 2393–2401. [Google Scholar] [CrossRef]
- Lippoldt, E.K.; Ongun, S.; Kusaka, G.K.; McKemy, D.D. Inflammatory and neuropathic cold allodynia are selectively mediated by the neurotrophic factor receptor GFRα3. Proc. Natl. Acad. Sci. USA 2016, 113, 4506–4511. [Google Scholar] [CrossRef]
- Caminski, E.S.; de Freitas, L.M.; Dallegrave, E.; Junior, C.A.d.S.; Gomez, M.V.; Pereira, E.M.R.; Antunes, F.T.T.; de Souza, A.H. Analgesic effects of the CTK 01512-2 toxin in different models of orofacial pain in rats. Pharmacol. Rep. 2020, 72, 600–611. [Google Scholar] [CrossRef]
- Baldwin, A.N.; Banks, M.L.; Marsh, S.A.; Townsend, E.A.; Venniro, M.; Shaham, Y.; Negus, S.S. Acute pain-related depression of operant responding maintained by social interaction or food in male and female rats. Psychopharmacology 2022, 239, 561–572. [Google Scholar] [CrossRef]
- Vilceanu, D.; Honore, P.; Hogan, Q.H.; Stucky, C.L. Spinal nerve ligation in mouse upregulates TRPV1 heat function in injured IB4-positive nociceptors. J. Pain 2010, 11, 588–599. [Google Scholar] [CrossRef]
- Tseng, K.; Li, H.; Clark, A.; Sundem, L.; Zuscik, M.; Noble, M.; Elfar, J. 4-Aminopyridine promotes functional recovery and remyelination in acute peripheral nerve injury. EMBO Mol. Med. 2016, 8, 1409–1420. [Google Scholar] [CrossRef]
- Kwiatkowski, K.; Piotrowska, A.; Rojewska, E.; Makuch, W.; Mika, J. The RS504393 Influences the Level of Nociceptive Factors and Enhances Opioid Analgesic Potency in Neuropathic Rats. J. Neuroimmune Pharmacol. 2017, 12, 402–419. [Google Scholar] [CrossRef]
- Mika, J.; Osikowicz, M.; Rojewska, E.; Korostynski, M.; Wawrzczak-Bargiela, A.; Przewlocki, R.; Przewlocka, B. Differential activation of spinal microglial and astroglial cells in a mouse model of peripheral neuropathic pain. Eur. J. Pharmacol. 2009, 623, 65–72. [Google Scholar] [CrossRef]
- Rojewska, E.; Popiolek-Barczyk, K.; Jurga, A.M.; Makuch, W.; Przewlocka, B.; Mika, J. Involvement of pro- and antinociceptive factors in minocycline analgesia in rat neuropathic pain model. J. Neuroimmunol. 2014, 277, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Zhang, Y.Q.; Qadri, Y.J.; Serhan, C.N.; Ji, R.R. Microglia in Pain: Detrimental and Protective Roles in Pathogenesis and Resolution of Pain. Neuron 2018, 100, 1292–1311. [Google Scholar] [CrossRef] [PubMed]
- Rojewska, E.; Makuch, W.; Przewlocka, B.; Mika, J. Minocycline prevents dynorphin-induced neurotoxicity during neuropathic pain in rats. Neuropharmacology 2014, 86, 301–310. [Google Scholar] [CrossRef] [PubMed]
- Pilat, D.; Rojewska, E.; Jurga, A.M.; Piotrowska, A.; Makuch, W.; Przewlocka, B.; Mika, J. IL-1 receptor antagonist improves morphine and buprenorphine efficacy in a rat neuropathic pain model. Eur. J. Pharmacol. 2015, 5, 240–248. [Google Scholar] [CrossRef]
- Green, J.M.; Sundman, M.H.; Chou, Y.H. Opioid-induced microglia reactivity modulates opioid reward, analgesia, and behavior. Neurosci. Biobehav. Rev. 2022, 135, 104544. [Google Scholar] [CrossRef]
- Mika, J. Modulation of microglia can attenuate neuropathic pain symptoms and enhance morphine effectiveness. Pharmacol. Rep. 2008, 60, 297–307. [Google Scholar] [CrossRef]
- Białecka, M.; Jurewicz, A.; Machoy-Mokrzyńska, A.; Kurzawski, M.; Leźnicka, K.; Dziedziejko, V.; Safranow, K.; Droździk, M.; Bohatyrewicz, A. Effect of interleukin 6 -174GC gene polymorphism on opioid requirements after total hip replacement. J. Anesth. 2016, 30, 562–567. [Google Scholar] [CrossRef]
- Bogacka, J.; Popiolek-Barczyk, K.; Pawlik, K.; Ciechanowska, A.; Makuch, W.; Rojewska, E.; Dobrogowski, J.; Przeklasa-Muszynska, A.; Mika, J. CCR4 antagonist (C021) influences the level of nociceptive factors and enhances the analgesic potency of morphine in a rat model of neuropathic pain. Eur. J. Pharmacol. 2020, 880, 173166. [Google Scholar] [CrossRef]
- Zychowska, M.; Rojewska, E.; Kreiner, G.; Nalepa, I.; Przewlocka, B.; Mika, J. Minocycline influences the anti-inflammatory interleukins and enhances the effectiveness of morphine under mice diabetic neuropathy. J. Neuroimmunol. 2013, 262, 35–45. [Google Scholar] [CrossRef]
- Kwiatkowski, K.; Pawlik, K.; Ciapała, K.; Piotrowska, A.; Makuch, W.; Mika, J. Bidirectional Action of Cenicriviroc, a CCR2/CCR5 Antagonist, Results in Alleviation of Pain-Related Behaviors and Potentiation of Opioid Analgesia in Rats with Peripheral Neuropathy. Front. Immunol. 2020, 11, 615327. [Google Scholar] [CrossRef]
- Pergolizzi, J.V.; Gharibo, C.; Magnusson, P.; Breve, F.; LeQuang, J.A.; Varrassi, G. Pharmacotherapeutic management of trigeminal neuropathic pain: An update. Expert Opin. Pharmacother. 2022, 23, 1155–1164. [Google Scholar] [CrossRef]
- Naseri, K.; Sabetkasaei, M.; Moini Zanjani, T.; Saghaei, E. Carbamazepine potentiates morphine analgesia on postoperative pain in morphine-dependent rats. Eur. J. Pharmacol. 2012, 674, 332–336. [Google Scholar] [CrossRef]
- Due, M.R.; Yang, X.F.; Allette, Y.M.; Randolph, A.L.; Ripsch, M.S.; Wilson, S.M.; Dustrude, E.T.; Khanna, R.; White, F.A. Carbamazepine potentiates the effectiveness of morphine in a rodent model of neuropathic pain. PLoS ONE 2014, 9, e107399. [Google Scholar] [CrossRef]
- Jun, I.G.; Kim, S.H.; Yoon, Y.I.; Park, J.Y. Intrathecal lamotrigine attenuates antinociceptive morphine tolerance and suppresses spinal glial cell activation in morphine-tolerant rats. J. Korean Med. Sci. 2013, 28, 300–307. [Google Scholar] [CrossRef]
- Codd, E.E.; Martinez, R.P.; Molino, L.; Rogers, K.E.; Stone, D.J.; Tallarida, R.J. Tramadol and several anticonvulsants synergize in attenuating nerve injury-induced allodynia. Pain 2008, 134, 254–262. [Google Scholar] [CrossRef]
- Due, M.R.; Piekarz, A.D.; Wilson, N.; Feldman, P.; Ripsch, M.S.; Chavez, S.; Yin, H.; Khanna, R.; White, F.A. Neuroexcitatory effects of morphine-3-glucuronide are dependent on Toll-like receptor 4 signaling. J. Neuroinflamm. 2012, 9, 200. [Google Scholar] [CrossRef]
- Kwiatkowski, K.; Mika, J. The importance of chemokines in neuropathic pain development and opioid analgesic potency. Pharmacol. Rep. 2018, 70, 821–830. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kocot-Kępska, M.; Pawlik, K.; Ciapała, K.; Makuch, W.; Zajączkowska, R.; Dobrogowski, J.; Przeklasa-Muszyńska, A.; Mika, J. Phenytoin Decreases Pain-like Behaviors and Improves Opioid Analgesia in a Rat Model of Neuropathic Pain. Brain Sci. 2023, 13, 858. https://doi.org/10.3390/brainsci13060858
Kocot-Kępska M, Pawlik K, Ciapała K, Makuch W, Zajączkowska R, Dobrogowski J, Przeklasa-Muszyńska A, Mika J. Phenytoin Decreases Pain-like Behaviors and Improves Opioid Analgesia in a Rat Model of Neuropathic Pain. Brain Sciences. 2023; 13(6):858. https://doi.org/10.3390/brainsci13060858
Chicago/Turabian StyleKocot-Kępska, Magdalena, Katarzyna Pawlik, Katarzyna Ciapała, Wioletta Makuch, Renata Zajączkowska, Jan Dobrogowski, Anna Przeklasa-Muszyńska, and Joanna Mika. 2023. "Phenytoin Decreases Pain-like Behaviors and Improves Opioid Analgesia in a Rat Model of Neuropathic Pain" Brain Sciences 13, no. 6: 858. https://doi.org/10.3390/brainsci13060858