

Effects of Group Size on Behavior, Reproduction, and mRNA Expression in Brains of Brandt’s Voles

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
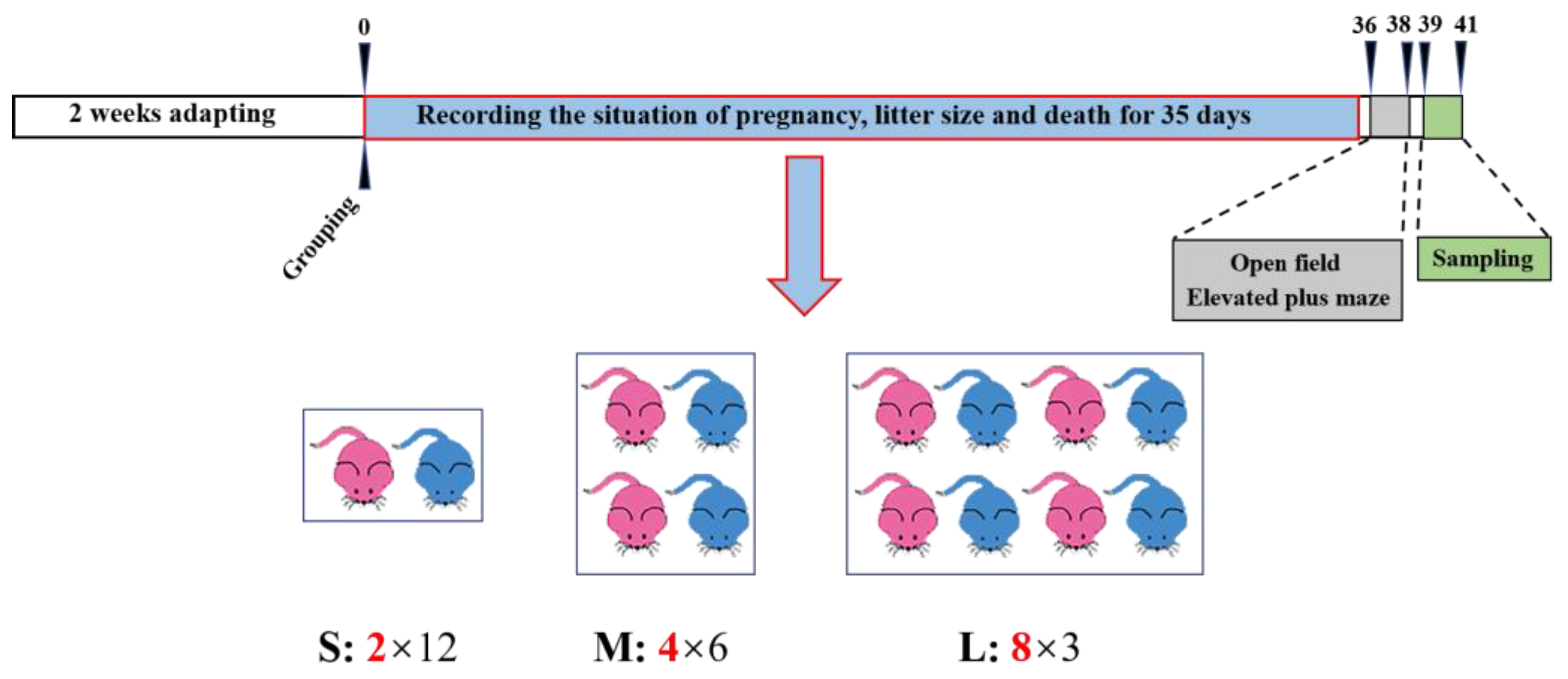
2.2. Experimental Design
2.3. Behavior Observations
2.4. Sampling
2.5. Reproductivity and Survival Rate
2.6. RNA Isolation and Reverse Transcription-PCR
2.7. Statistical Analysis
3. Results
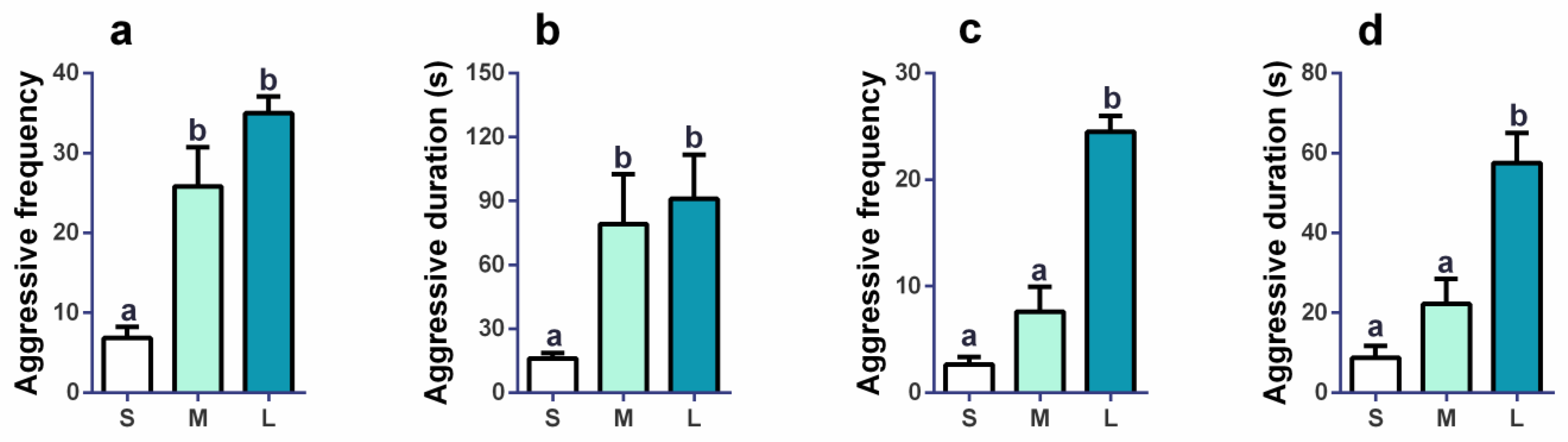
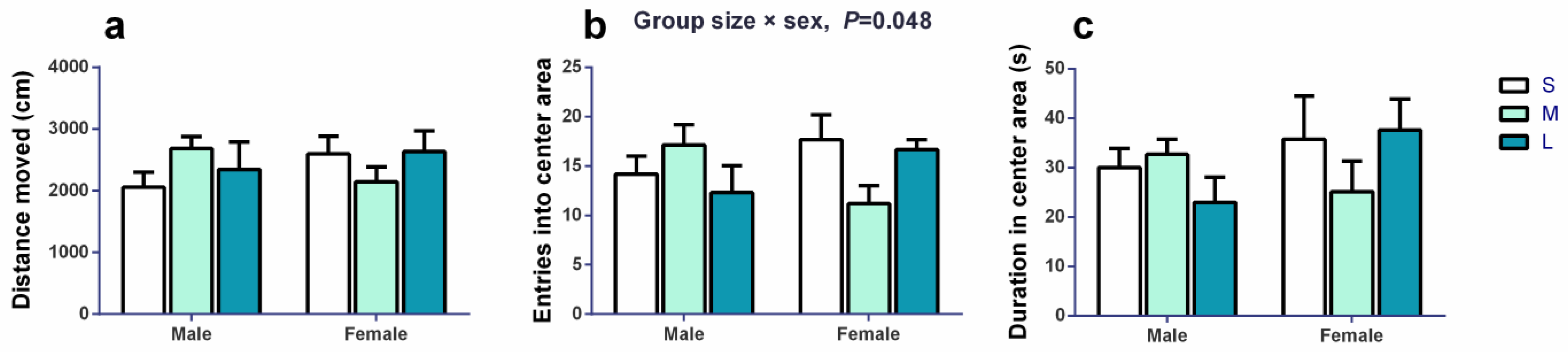
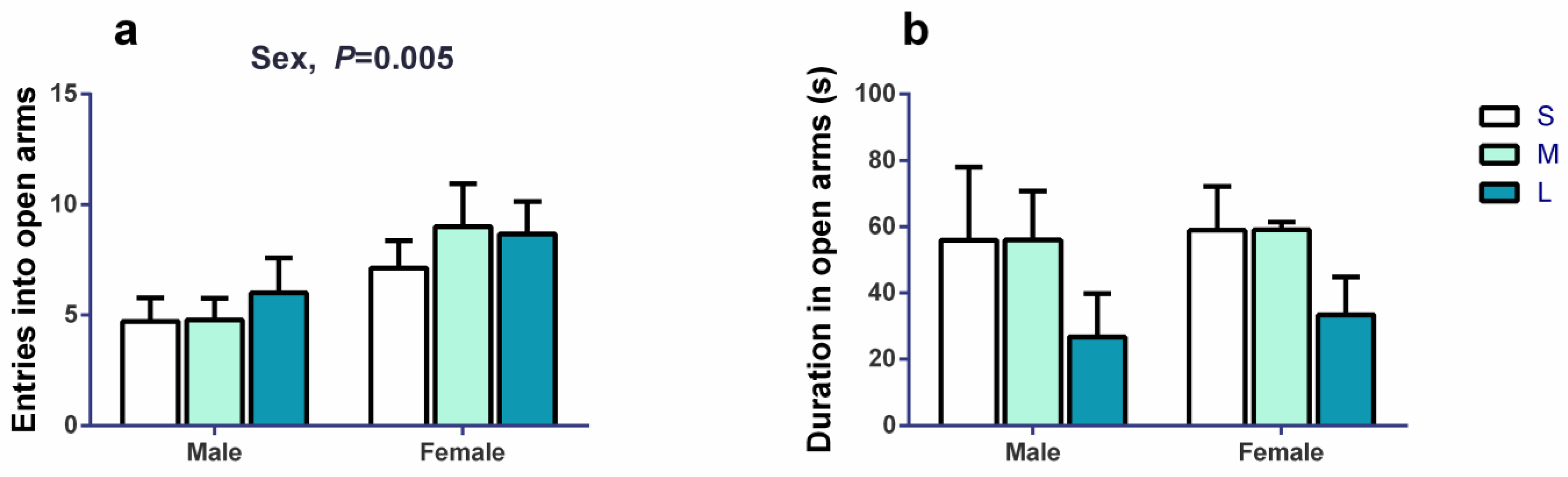
3.1. Impacts of Initial Groups Size on Aggression Behaviors and Anxiety Behaviors
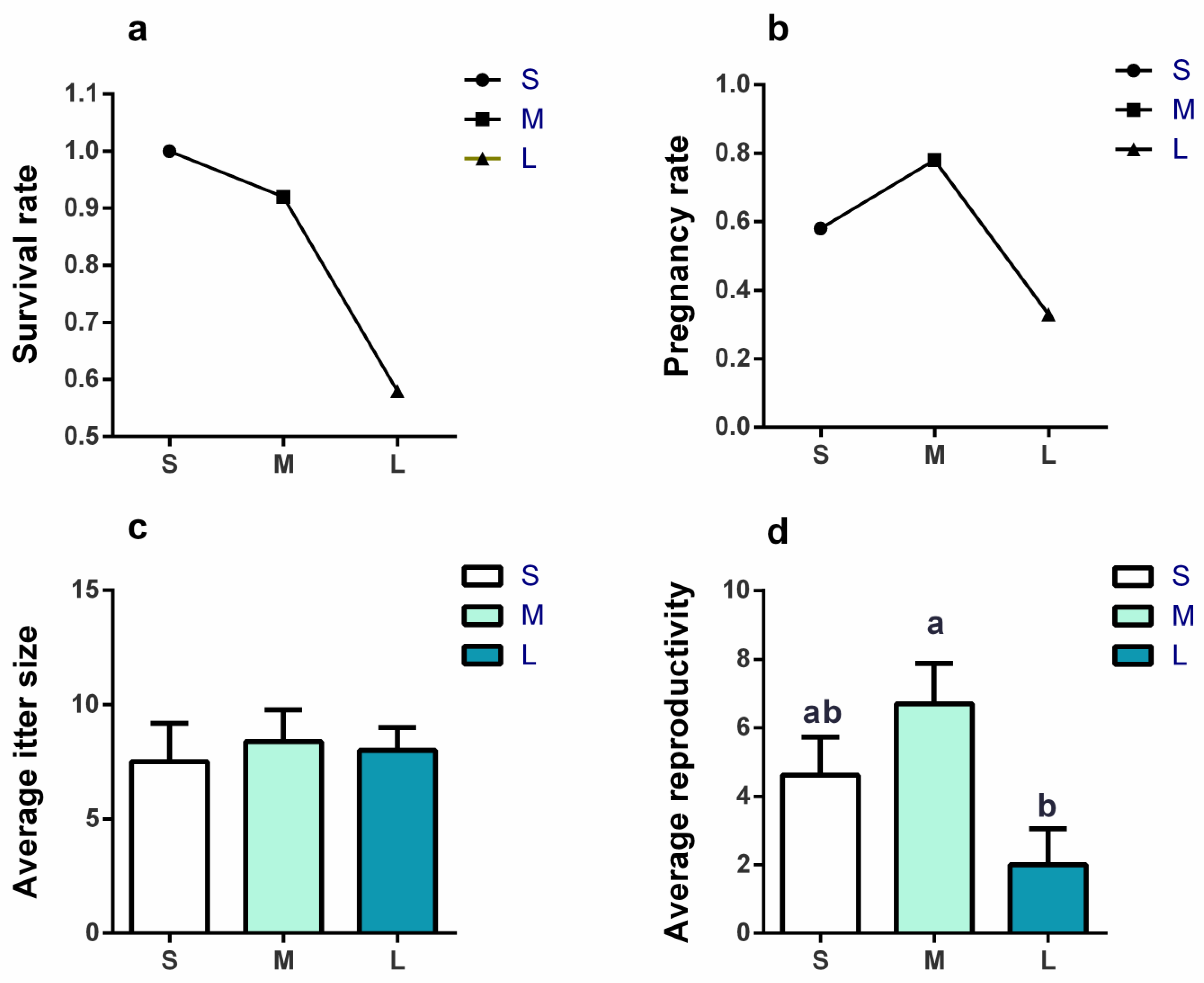
3.2. Effects of Initial Group Size on Survival Rate and Reproductivity
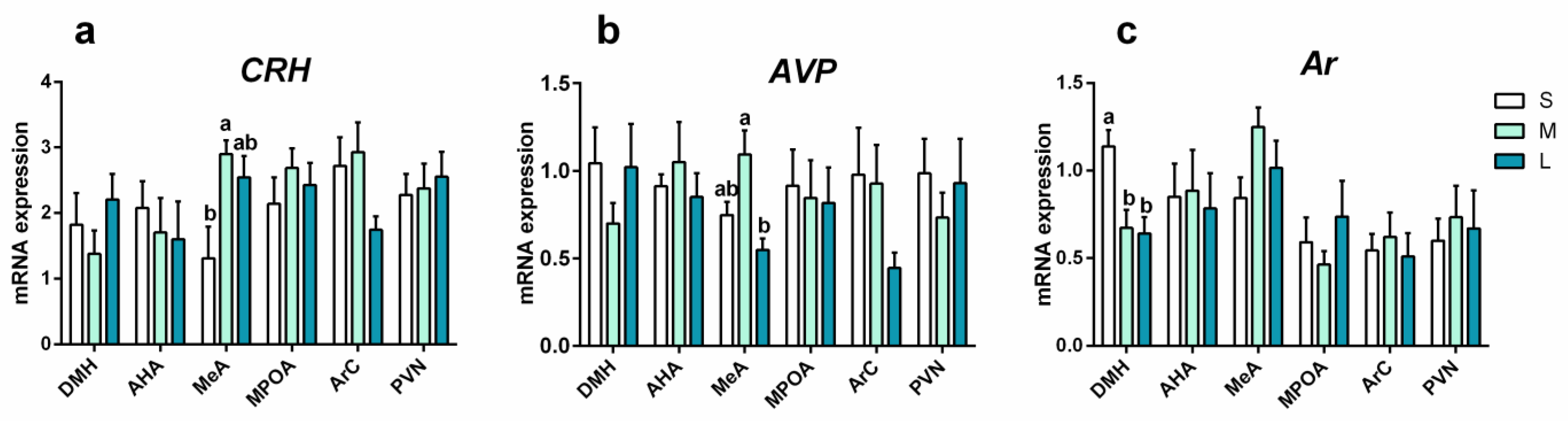
3.3. Effects of Initial Group Size on mRNA Expression in Male Brain
3.3.1. mRNA Expression of Stress-Response Genes
3.3.2. mRNA Expression of Reproduction-Response Genes
3.4. Effects of Final Density or Group Size on mRNA Expression in Male Brain
4. Discussion
4.1. Impact of Group Size on Behaviors and mRNA Expression
4.2. Impacts of Group Size on Reproduction and Gene Expression
4.3. Limitation of the Study
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alcock, J. Animal Behavior: An Evolutionary Approach, 10th ed.; Sinauer Associates: Sunderland, UK, 2013. [Google Scholar]
- Ebensperger, L.A. A review of the evolutionary causes of rodent group-living. Acta Theriol. 2001, 46, 115–144. [Google Scholar] [CrossRef]
- Ebensperger, L.A.; Correa, L.A.; Leon, C.; Ramirez-Estrada, J.; Abades, S.; Villegas, A.; Hayes, L.D. The modulating role of group stability on fitness effects of group size is different in females and males of a communally rearing rodent. J. Anim. Ecol. 2016, 85, 1502–1515. [Google Scholar] [CrossRef] [PubMed]
- McGuire, B.; Getz, L.L.; Oli, M.K. Fitness consequences of sociality in prairie voles, Microtus ochrogaster: Influence of group size and composition. Anim. Behav. 2002, 64, 645–654. [Google Scholar] [CrossRef]
- Beery, A.K.; Kaufer, D. Stress, social behavior, and resilience: Insights from rodents. Neurobiol. Stress 2015, 1, 116–127. [Google Scholar] [CrossRef] [PubMed]
- Jirkof, P.; Bratcher, N.; Medina, L.; Strasburg, D.; Ebert, P.; Gaskill, B.N. The effect of group size, age and handling frequency on inter-male aggression in CD 1 mice. Sci. Rep. 2020, 10, 2253. [Google Scholar] [CrossRef]
- Van Loo, P.L.; Mol, J.A.; Koolhaas, J.M.; Van Zutphen, B.F.; Baumans, V. Modulation of aggression in male mice: Influence of group size and cage size. Physiol. Behav. 2001, 72, 675–683. [Google Scholar] [CrossRef]
- Slattery, D.A.; Uschold, N.; Magoni, M.; Bär, J.; Popoli, M.; Neumann, I.D.; Reber, S.O. Behavioural consequences of two chronic psychosocial stress paradigms: Anxiety without depression. Psychoneuroendocrinology 2012, 37, 702–714. [Google Scholar] [CrossRef] [PubMed]
- Lardy, S.; Allainé, D.; Bonenfant, C.; Cohas, A. Sex-specific determinants of fitness in a social mammal. Ecology 2015, 96, 2947–2959. [Google Scholar] [CrossRef]
- Wrangham, R.W. An ecological model of female-bonded primate groups. Behaviour 1980, 75, 262–300. [Google Scholar] [CrossRef]
- Zhang, Z.; Yan, C.; Krebs, C.J.; Stenseth, N.C. Ecological non-monotonicity and its effects on complexity and stability of populations, communities and ecosystems. Ecol. Model. 2015, 312, 374–384. [Google Scholar] [CrossRef]
- McEwen, B.S. The ever-changing brain: Cellular and molecular mechanisms for the effects of stressful experiences. Dev. Neurobiol. 2012, 72, 878–890. [Google Scholar] [CrossRef] [PubMed]
- Lupien, S.J.; McEwen, B.S.; Gunnar, M.R.; Heim, C. Effects of stress throughout the lifespan on the brain, behaviour and cognition. Nat. Rev. Neurosci. 2009, 10, 434–445. [Google Scholar] [CrossRef]
- Hauger, R.L.; Millan, M.A.; Catt, K.J.; Aguilera, G. Differential regulation of brain and pituitary corticotropin-releasing factor receptors by corticosterone. Endocrinology 1987, 120, 1527–1533. [Google Scholar] [CrossRef]
- Drouin, J.; Sun, Y.L.; Chamberland, M.; Gauthier, Y.; Delean, A.; Nemer, M.; Schmidt, T.J. Novel glucocorticoid receptor complex with DNA element of the hormone-repressed POMC gene. EMBO J. 1993, 12, 145–156. [Google Scholar] [CrossRef] [PubMed]
- Keeney, A.; Jessop, D.S.; Harbuz, M.S.; Marsden, C.A.; Hogg, S.; Blackburn-Munro, R.E. Differential effects of acute and chronic social defeat stress on hypothalamic-pituitary-adrenal axis function and hippocampal serotonin release in mice. J. Neuroendocrinol. 2006, 18, 330–338. [Google Scholar] [CrossRef]
- Winslow, J.T.; Hearn, E.F.; Ferguson, J.; Young, L.J.; Matzuk, M.M.; Insel, T.R. Infant vocalization, adult aggression, and fear behavior of an oxytocin null mutant mouse. Horm. Behav. 2000, 37, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Acevedo-Rodriguez, A.; Mani, S.K.; Handa, R.J. Oxytocin and estrogen receptor beta in the brain: An Overview. Front. Endocrinol. 2015, 6, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Ferris, C.F.; Potegal, M. Vasopressin receptor blockade in the anterior hypothalamus suppresses aggression in hamsters. Physiol. Behav. 1988, 44, 235–239. [Google Scholar] [CrossRef] [PubMed]
- Ruscio, M.G.; Sweeny, T.; Hazelton, J.; Suppatkul, P.; Sue Carter, C. Social environment regulates corticotropin releasing factor, corticosterone and vasopressin in juvenile prairie voles. Horm. Behav. 2007, 51, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Miczek, K.A.; Hussain, S.; Faccidomo, S. Alcohol-heightened aggression in mice: Attenuation by 5-HT1A receptor agonists. Psychopharmacology 1998, 139, 160–168. [Google Scholar] [CrossRef] [PubMed]
- D’Anglemont de Tassigny, X.; Colledge, W.H. The role of kisspeptin signaling in reproduction. Physiology 2010, 25, 207–217. [Google Scholar] [PubMed]
- Williamson, C.M.; Romeo, R.D.; Curley, J.P. Dynamic changes in social dominance and mPOA GnRH expression in male mice following social opportunity. Horm. Behav. 2017, 87, 80–88. [Google Scholar] [CrossRef]
- Javed, Z.; Qamar, U.; Sathyapalan, T. The role of kisspeptin signalling in the hypothalamic-pituitary-gonadal axis--current perspective. Endokrynol Pol 2015, 66, 534–547. [Google Scholar] [PubMed]
- Celik, O.; Aydin, S.; Celik, N.; Yilmaz, M. Peptides: Basic determinants of reproductive functions. Peptides 2015, 72, 34–43. [Google Scholar] [CrossRef]
- Kauffman, A.S.; Park, J.H.; McPhie-Lalmansingh, A.A.; Gottsch, M.L.; Bodo, C.; Hohmann, J.G.; Pavlova, M.N.; Rohde, A.D.; Clifton, D.K.; Steiner, R.A.; et al. The kisspeptin receptor gpr54 is required for sexual differentiation of the brain and behavior. J. Neurosci. 2007, 27, 8826–8835. [Google Scholar] [CrossRef]
- Kriegsfeld, L.J.; Mei, D.F.; Bentley, G.E.; Ubuka, T.; Mason, A.O.; Inoue, K.; Ukena, K.; Tsutsui, K.; Silver, R. Identification and characterization of a gonadotropin-inhibitory system in the brains of mammals. Proc. Natl. Acad. Sci. USA 2006, 103, 2410–2415. [Google Scholar] [CrossRef] [PubMed]
- Peragine, D.E.; Pokarowski, M.; Mendoza-Viveros, L.; Swift-Gallant, A.; Cheng, H.M.; Bentley, G.E.; Holmes, M.M. RFamide-related peptide-3 (RFRP-3) suppresses sexual maturation in a eusocial mammal. Proc. Natl. Acad. Sci. USA 2017, 114, 1207–1212. [Google Scholar] [CrossRef] [PubMed]
- Raskin, K.; de Gendt, K.; Duittoz, A.; Liere, P.; Verhoeven, G.; Tronche, F.; Mhaouty-Kodja, S. Conditional inactivation of androgen receptor gene in the nervous system: Effects on male behavioral and neuroendocrine responses. J. Neurosci. 2009, 29, 4461–4470. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.T.; Dungan, H.M.; Stoll, E.A.; Gottsch, M.L.; Braun, R.E.; Eacker, S.M.; Clifton, D.K.; Steiner, R.A. Differential regulation of KiSS-1 mRNA expression by sex steroids in the brain of the male mouse. Endocrinology 2005, 146, 2976–2984. [Google Scholar] [CrossRef] [PubMed]
- Simerly, R.B.; Chang, C.; Muramatsu, M.; Swanson, L.W. Distribution of androgen and estrogen receptor mRNA-containing cells in the rat brain: An in situ hybridization study. J. Comp. Neurol. 1990, 294, 76–95. [Google Scholar] [CrossRef] [PubMed]
- Ruscio, M.G.; Sweeny, T.D.; Gomez, A.; Parker, K.; Carter, C.S. Social environment alters central distribution of estrogen receptor alpha in juvenile prairie voles. Physiol. Behav. 2009, 98, 296–301. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, S.; Kruijver, F.P.; Straub, R.H.; Sachser, N.; Swaab, D.F. Early social stress in male Guinea-pigs changes social behaviour, and autonomic and neuroendocrine functions. J. Neuroendocrinol. 2003, 15, 761–769. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhong, W. Investigations of reproduction in populations of Brandt’s voles. Acta Zool. Sin. 1979, 25, 250–259. [Google Scholar]
- Wang, D.; Li, N.; Tian, L.; Ren, F.; Li, Z.; Chen, Y.; Liu, L.; Hu, X.; Zhang, X.; Song, Y.; et al. Dynamic expressions of hypothalamic genes regulate seasonal breeding in a natural rodent population. Mol. Ecol. 2019, 28, 3508–3522. [Google Scholar] [CrossRef]
- Yin, F.; Fang, J. Research advances in population ecology and behavioral ecology of Brandt’s voles (Microtus brandti). Chin. J. Ecol. 1998, 17, 65–69. [Google Scholar]
- Zhong, W.; Wang, G.; Zhou, Q.; Wang, G. Communal food caches and social groups of Brandt’s voles in the typical steppes of Inner Mongolia, China. J. Arid Environ. 2007, 68, 398–407. [Google Scholar] [CrossRef]
- Huo, Y.J.; Wan, X.R.; Wolff, J.O.; Wang, G.; Thomas, S.; Iglay, R.B.; Leopold, B.D.; Liu, W. Multiple paternities increase genetic diversity of offspring in Brandt’s voles. Behav. Process 2010, 84, 745–749. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.H.; Yue, L.F.; Wang, D.W.; Li, N.; Cong, L. Inbreeding avoidance drives consistent variation of fine-scale genetic structure caused by dispersal in the seasonal mating system of Brandt’s voles. PLoS ONE 2013, 8, e58101. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Shi, D. Study on reproductive behavior of different social hierarchy of Brandt’ s vole. Acta Theriol. Sin. 2003, 23, 220–224. [Google Scholar]
- Huang, S.; Li, G.; Pan, Y.; Song, M.; Zhao, J.; Wan, X.; Krebs, C.J.; Wang, Z.; Han, W.; Zhang, Z. Density-induced social stress alters oxytocin and vasopressin activities in the brain of a small rodent species. Integr. Zool. 2020, 16, 149–159. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Zhou, L.Y.; Cao, J.X.; Zhang, Y.Q.; Yang, F.P.; Wang, A.Q.; Wei, W.H.; Yang, S.M. Effect of group density on the physiology and aggressive behavior of male Brandt’s voles (Lasiopodomys brandtii). Zool. Stud. 2018, 57, e35. [Google Scholar] [PubMed]
- Wan, X.; Liu, W.; Wang, G.; Wang, M.; Zhong, W. Seasonal changes of the activity patterns of Brandt’s vole (Lasiopodomys brandtii) in the typical steppe in Inner Mongolia. Acta Theriol. Sin. 2006, 26, 226–234. [Google Scholar]
- Sun, H.; Li, C.; Zhang, Y.; Jiang, M.; Dong, Q.; Wang, Z. Light-resetting impact on behavior and the central circadian clock in two vole species (genus: Lasiopodomys). Comp. Biochem. Phys. B. 2020, 248, 110478. [Google Scholar] [CrossRef] [PubMed]
- Hathaway, G. Oxytocin, Aggression, and Neural Activity in the Naked Mole-Rat. Master’s thesis, University of Toronto, Toronto, OT, Canada, 2014. [Google Scholar]
- Bigi, S.; Maestripieri, D.; Aloe, L.; Alleva, E. NGF decreases isolation-induced aggressive behavior, while increasing adrenal volume, in adult male mice. Physiol. Behav. 1992, 51, 337–343. [Google Scholar] [CrossRef] [PubMed]
- Alleva, E. Assessment of Aggressive Behavior in Rodents. In Methods in Neurosciences; Conn, P.M., Ed.; Academic Press: Cambridge, MA, USA, 1993; pp. 111–137. [Google Scholar]
- Yan, L.; Sun, X.; Wang, Z.; Song, M.; Zhang, Z. Regulation of social behaviors by p-Stat3 via oxytocin and its receptor in the nucleus accumbens of male Brandt’s voles (Lasiopodomys brandtii). Horm. Behav. 2020, 119, 1–8. [Google Scholar] [CrossRef]
- Kim, J.; Semaan, S.J.; Clifton, D.K.; Steiner, R.A.; Dhamija, S.; Kauffman, A.S. Regulation of kiss1 expression by sex steroids in the amygdala of the rat and mouse. Endocrinology 2011, 152, 2020–2030. [Google Scholar] [CrossRef]
- Nelson, R.J.; Trainor, B.C. Neural mechanisms of aggression. Nat. Rev. Neurosci. 2007, 8, 536–546. [Google Scholar] [CrossRef]
- Palanza, P. Animal models of anxiety and depression: How are females different? Neurosci. Biobehav. Rev. 2001, 25, 219–233. [Google Scholar] [CrossRef]
- Johnston, A.L.; File, S.E. Sex differences in animal tests of anxiety. Physiol. Behav. 1991, 49, 245–250. [Google Scholar] [CrossRef]
- Berry, A.; Bellisario, V.; Capoccia, S.; Tirassa, P.; Calza, A.; Alleva, E.; Cirulli, F. Social deprivation stress is a triggering factor for the emergence of anxiety- and depression-like behaviours and leads to reduced brain BDNF levels in C57BL/6J mice. Psychoneuroendocrinology 2012, 37, 762–772. [Google Scholar] [CrossRef]
- Figueiredo, H.F.; Bruestle, A.; Bodie, B.; Dolgas, C.M.; Herman, J.P. The medial prefrontal cortex differentially regulates stress-induced c-fos expression in the forebrain depending on type of stressor. Eur. J. Neurosci. 2003, 18, 2357–2364. [Google Scholar] [CrossRef]
- Figueiredo, H.F.; Bodie, B.L.; Tauchi, M.; Dolgas, C.M.; Herman, J.P. Stress integration after acute and chronic predator stress: Differential activation of central stress circuitry and sensitization of the hypothalamo-pituitary-adrenocortical axis. Endocrinology 2003, 144, 5249–5258. [Google Scholar] [CrossRef] [PubMed]
- Dallman, M.F.; Pecoraro, N.; Akana, S.F.; La Fleur, S.E.; Gomez, F.; Houshyar, H.; Bell, M.E.; Bhatnagar, S.; Laugero, K.D.; Manalo, S. Chronic stress and obesity: A new view of “comfort food”. Proc. Natl. Acad. Sci. USA 2003, 100, 11696–11701. [Google Scholar] [CrossRef] [PubMed]
- Herman, J.P.; Figueiredo, H.; Mueller, N.K.; Ulrich-Lai, Y.; Ostrander, M.M.; Choi, D.C.; Cullinan, W.E. Central mechanisms of stress integration: Hierarchical circuitry controlling hypothalamo-pituitary-adrenocortical responsiveness. Front. Neuroendocrinol. 2003, 24, 151–180. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Huang, S.; Li, G.; Zhao, J.; Lu, W.; Zhang, Z. High housing density increases stress hormone- or disease-associated fecal microbiota in male Brandt’s voles (Lasiopodomys brandtii). Horm. Behav. 2020, 126, 104838. [Google Scholar] [CrossRef]
- Caldwell, H.K.; Lee, H.J.; Macbeth, A.H.; Young, W.S. Vasopressin: Behavioral roles of an “original” neuropeptide. Prog. Neurobiol. 2008, 84, 1–24. [Google Scholar] [CrossRef]
- Wang, Z.; De Vries, G.J. Androgen and estrogen effects on vasopressin messenger RNA expression in the medial amygdaloid nucleus in male and female rats. J. Neuroendocrinol. 1995, 7, 827–831. [Google Scholar] [CrossRef]
- Wang, Z.; Bullock, N.A.; De Vries, G.J. Sexual differentiation of vasopressin projections of the bed nucleus of the stria terminals and medial amygdaloid nucleus in rats. Endocrinology 1993, 132, 2299–2306. [Google Scholar] [CrossRef]
- Cooke, B.M. Steroid-dependent plasticity in the medial amygdala. Neuroscience 2006, 138, 997–1005. [Google Scholar] [CrossRef]
- Auger, C.J.; Coss, D.; Auger, A.P.; Forbes-Lorman, R.M. Epigenetic control of vasopressin expression is maintained by steroid hormones in the adult male rat brain. Proc. Natl. Acad. Sci. USA 2011, 108, 4242–4247. [Google Scholar] [CrossRef]
- Chalmers, D.T.; Watson, S.J. Comparative anatomical distribution of 5-HT1A receptor mRNA and 5-HT1A binding in rat brain —A combined in situ hybridisation/in vitro receptor autoradiographic study. Brain Res. 1991, 561, 51–60. [Google Scholar] [CrossRef]
- Gao, G. Htr1a expression in brain is positively correlated with the aggressive behavior of male Chinese striped hamster (Cricetulus barabensis). Master’s thesis, Qufu Normal University, Qufu, China, 2019. [Google Scholar]
- Zhuang, X.; Gross, C.; Santarelli, L.; Compan, V.; Trillat, A.C.; Hen, R. Altered emotional states in knockout mice lacking 5-HT1A or 5-HT1B receptors. Neuropsychopharmacology 1999, 21, 52S–60S. [Google Scholar] [CrossRef]
- Christian, J.J. Population density and reproductive efficiency. Biol. Reprod. 1971, 4, 248–294. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Zhong, W.; Wang, G. Density factor in the regulation of Brandt’s vole population. Acta Theriol. Sin. 1992, 12, 49–56. [Google Scholar]
- Zhang, X.; Zhang, M.; Han, Q.H.; Guo, C.; Zhou, X.; Li, B.; Wáng, Y.X. Effects of density on sex organ development and female sexual maturity in laboratory-bred Microtus fortis. Anim. Biol. 2018, 68, 39–54. [Google Scholar] [CrossRef]
- Trivers, R. Parental Investment and Sexual Selection; Campbell, B., Ed.; Routledge: Abingdon, UK, 1972; Volume 112. [Google Scholar]
- Zarrow, M.X.; Clark, J.H. Ovulation following vaginal stimulation in a spontaneous ovulator and its implications. J. Endocrinol. 1968, 40, 343–352. [Google Scholar] [CrossRef]
- Solomon, N.G.; Keane, B. Chapter 4. Reproductive Strategies in Female Rodents. In Rodent Societies: An Ecological and Evolutionary Perspective; Jerry, O.W., Paul, W.S., Eds.; University of Chicago Press: Chicago, IL, USA, 2008; pp. 42–56. [Google Scholar]
- Milligan, S.R. Induced ovulation in mammals. Oxf. Rev. Reprod. Biol. 1982, 4, 1–46. [Google Scholar]
- Fisher, D.O.; Double, M.C.; Blomberg, S.P.; Jennions, M.D.; Cockburn, A. Post-mating sexual selection increases lifetime fitness of polyandrous females in the wild. Nature 2006, 444, 89–92. [Google Scholar] [CrossRef]
- Madsen, T.; Shine, R.; Loman, J.; Håkansson, T. Why do female adders copulate so frequently? Nature 1992, 355, 440–441. [Google Scholar] [CrossRef]
- Rivier, C.; Rivier, J.; Vale, W. Stress-induced inhibition of reproductive functions: Role of endogenous corticotropin-releasing factor. Science 1986, 231, 607–609. [Google Scholar] [CrossRef] [PubMed]
- Rivier, C.; Rivest, S. Effect of stress on the activity of the hypothalamic-pituitary-gonadal axis: Peripheral and central mechanisms. Biol. Reprod. 1991, 45, 523–532. [Google Scholar] [CrossRef]
- Li, X.F.; Knox, A.M.; O’Byrne, K.T. Corticotrophin-releasing factor and stress-induced inhibition of the gonadotrophin-releasing hormone pulse generator in the female. Brain Res. 2010, 1364, 153–163. [Google Scholar] [CrossRef]
- Kirby, E.D.; Geraghty, A.C.; Ubuka, T.; Bentley, G.E.; Kaufer, D. Stress increases putative gonadotropin inhibitory hormone and decreases luteinizing hormone in male rats. Proc. Natl. Acad. Sci. USA 2009, 106, 11324–11329. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.; Lee, S.O.; Wang, R.S.; Yeh, S.; Chang, T.M. Androgen receptor (AR) physiological roles in male and female reproductive systems: Lessons learned from AR-knockout mice lacking AR in selective cells. Biol. Reprod. 2013, 89, 21. [Google Scholar] [CrossRef] [PubMed]
- Thompson, R.H.; Swanson, L.W. Structural characterization of a hypothalamic visceromotor pattern generator network. Brain Res. Brain Res. Rev. 2003, 41, 153–202. [Google Scholar] [CrossRef]
- Maywood, E.; Bittman, E.; Hastings, M. Lesions of the melatonin- and androgen-responsive tissue of the dorsomedial nucleus of the hypothalamus block the gonadal response of male syrian hamsters to programmed infusions of melatonin1. Biol. Reprod. 1996, 54, 470–477. [Google Scholar] [CrossRef] [Green Version]
- Lewis, D.; Freeman, D.A.; Dark, J.; Wynne-Edwards, K.E.; Zucker, I. Photoperiodic control of oestrous cycles in Syrian hamsters: Mediation by the mediobasal hypothalamus. J. Neuroendocrinol. 2002, 14, 294–299. [Google Scholar] [CrossRef]
- Voigt, C.; Bennett, N.C. Gene expression pattern of Kisspeptin and RFamide-related peptide (Rfrp) in the male Damaraland mole-rat hypothalamus. J. Chem. Neuroanat. 2021, 118, 102039. [Google Scholar] [CrossRef]
- Bialy, M.; Nikolaev-Diak, A.; Kalata, U.; Nikolaev, E. Blockade of androgen receptor in the medial amygdala inhibits noncontact erections in male rats. Physiol. Behav. 2011, 10, 295–301. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Primers | Products (bps) | |
|---|---|---|---|
| (A) Stress-response genes | |||
| Corticotropin-releasing hormone (CRH) | F | 5′ GCAAATGCTGCGTGCTTTC 3′ | 231 |
| R | 5′ TCTCTTCTCCTCCCTTGGTAG 3′ | ||
| Pro-opiomelanocortin-alpha (POMC) | F | 5′ TTGGAAAGATAGCGGGAGAG 3′ | 187 |
| R | 5′ GCAGAGGCAAACAAGATTGG 3′ | ||
| Arginine vasopressin (AVP) | F | 5′ ACGCTCTCCGCTTGTTTC 3′ | 110 |
| R | 5′ CACTGTCTCAGCTCCATGTC 3′ | ||
| Oxytocin (OT) | F | 5′ TGCCAGGAGGAGAACTACC 3′ | 149 |
| R | 5′ TCCGAGAAGGCAGACTCAG 3′ | ||
| 5-hydroxytryptamine receptor1A (Htr1a) | F | 5′ GTTGGACAGCGACAAAGTG 3′ | 212 |
| R | 5′ TGGAGCGGGAAGTTCTTAC 3′ | ||
| (B) Reproduction-response genes | |||
| Gonadotropin-releasing hormone 1 (GnRH) | F | 5′ CGGCATTCTACTGCTGACTG 3′ | 239 |
| R | 5′ GCCTGGCTTCCTCTTCAATC 3′ | ||
| Kiss1 | F | 5′ AAGGAATCGCGGTATGCAG 3′ | 203 |
| R | 5′ CCGAAGGAGTTCCAGTTGTAG 3′ | ||
| RFamide-related peptides (Rfrp) | F | 5′ CAAGACACCCGCTGATTTGC 3′ | 113 |
| R | 5′ TTCGCTTTCCACCAGGACTC 3′ | ||
| Estrogen receptor 1 (alpha) (Esr1) | F | 5′ AATGACTATGCCTCTGGCTACC 3′ | 184 |
| R | 5′ TGCCCACTTCGTAACACTTG 3′ | ||
| Androgen receptor (Ar) | F | 5′ CAGAGGCAAAGTCTAAAGC 3′ | 125 |
| R | 5′ CAACTATGGTGGAGATTCG 3′ | ||
| (C) Reference gene | |||
| Glyceraldehyde-3-phosphate dehydrogenase (Gapdh) | F | 5′ ATCACTGCCACCCAGAAG 3′ | 191 |
| R | 5′ TCCACGACGGACACATTG 3′ | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, W.; Huang, S.; Liu, J.; Batsuren, E.; Li, G.; Wan, X.; Zhao, J.; Wang, Z.; Han, W.; Zhang, Z. Effects of Group Size on Behavior, Reproduction, and mRNA Expression in Brains of Brandt’s Voles. Brain Sci. 2023, 13, 311. https://doi.org/10.3390/brainsci13020311
Lu W, Huang S, Liu J, Batsuren E, Li G, Wan X, Zhao J, Wang Z, Han W, Zhang Z. Effects of Group Size on Behavior, Reproduction, and mRNA Expression in Brains of Brandt’s Voles. Brain Sciences. 2023; 13(2):311. https://doi.org/10.3390/brainsci13020311
Chicago/Turabian StyleLu, Wei, Shuli Huang, Jing Liu, Erdenetuya Batsuren, Guoliang Li, Xinru Wan, Jidong Zhao, Zuoxin Wang, Wenxuan Han, and Zhibin Zhang. 2023. "Effects of Group Size on Behavior, Reproduction, and mRNA Expression in Brains of Brandt’s Voles" Brain Sciences 13, no. 2: 311. https://doi.org/10.3390/brainsci13020311