
Source Localization of Somatosensory Neural Generators in Adults with Attention-Deficit/Hyperactivity Disorder
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
1.1. Neural Attributes of ADHD
1.2. Behavioural Characteristics of ADHD
1.3. Somatosensory Function and Sensorimotor Integration
1.4. Source Localization
1.5. Rationale and Purpose
2. Materials and Methods
2.1. Ethical Approval
2.2. Participants
2.3. Procedures
2.3.1. SEPs Stimulation Parameters
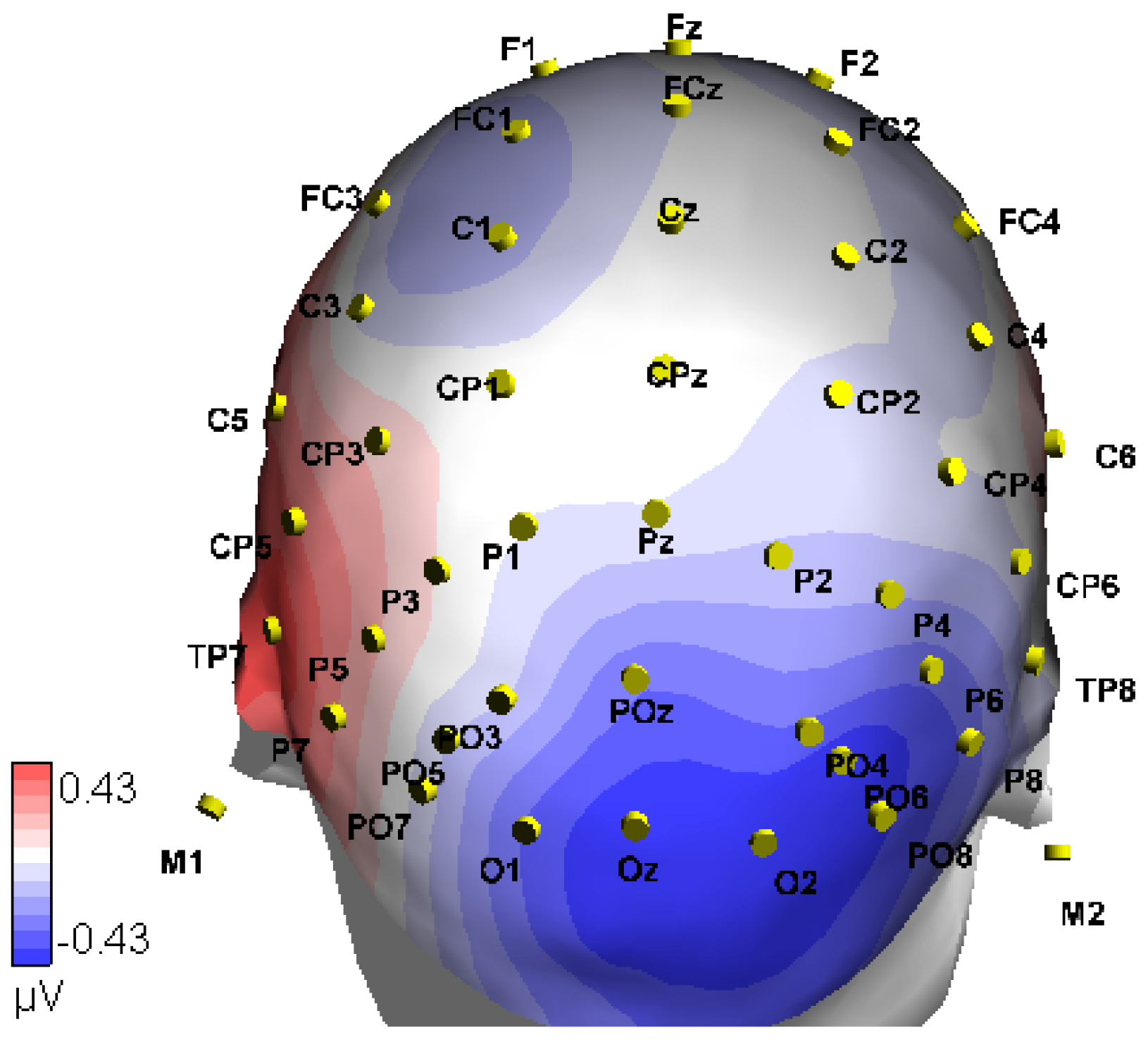
2.3.2. EEG Collection Parameters
2.3.3. Paradigm(s)
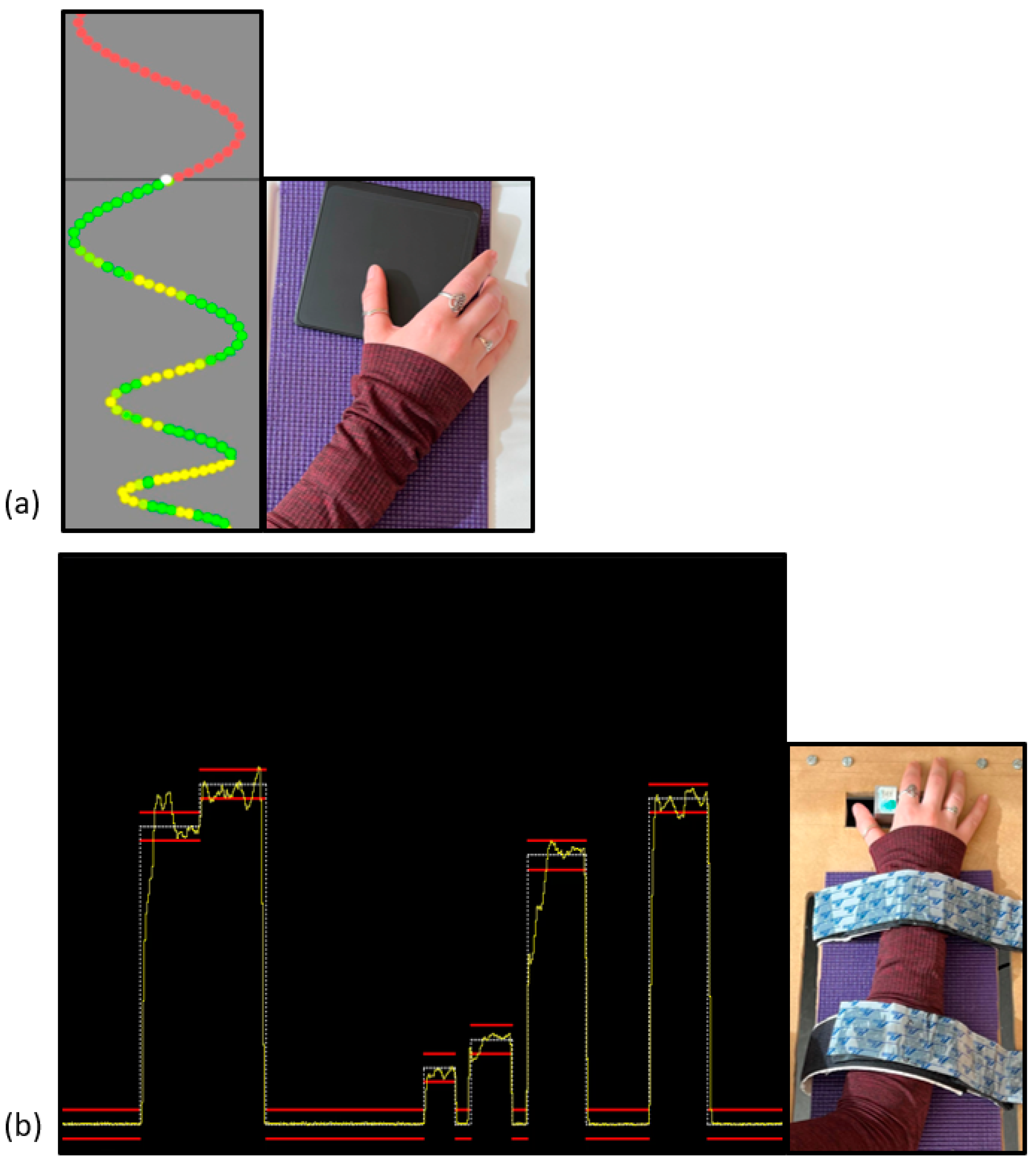
Novel Visuomotor Tracing Task (MTT) Parameters
Novel Force-Matching Task (FMT) Parameters
2.4. Data Processing and Statistical Analysis
2.4.1. EEG/SEPs
2.4.2. Source Localization–sLORETA Analysis
- (1)
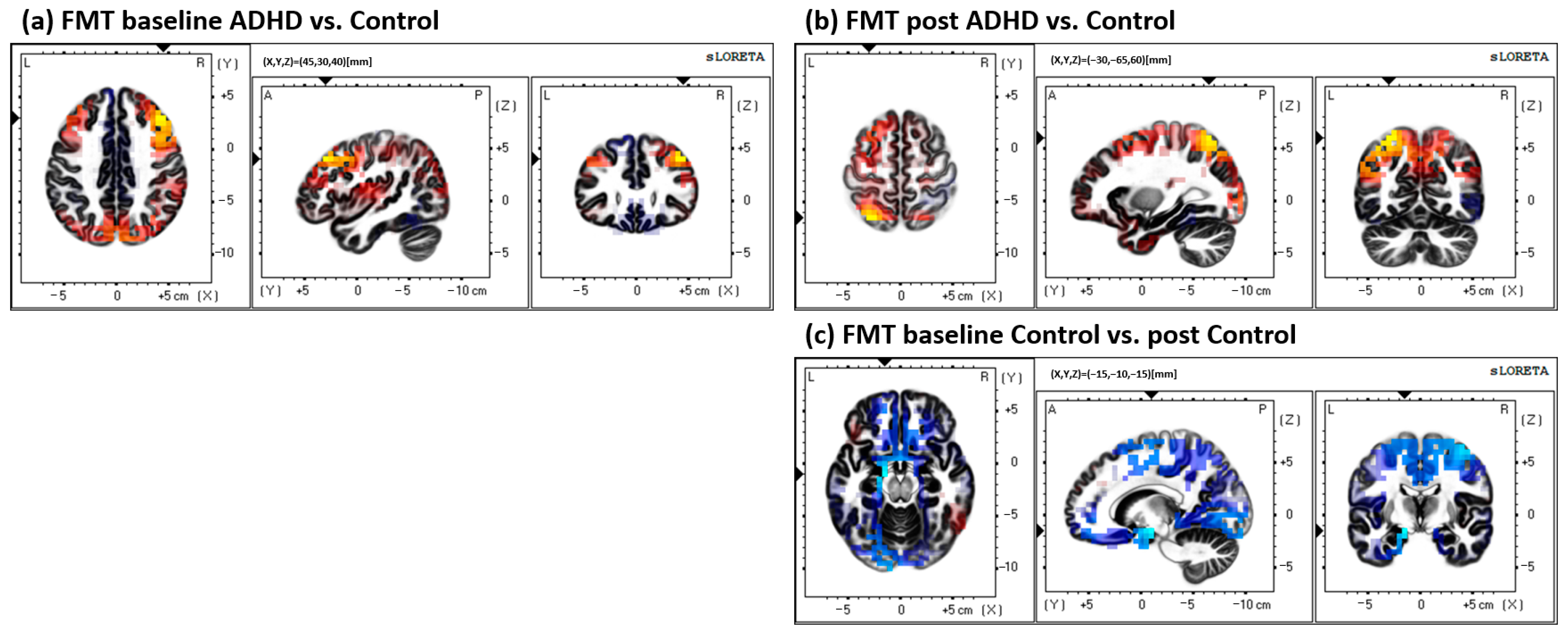
- Between groups (ADHD vs. control) at both baseline and post measures. Comparisons were performed for both the MTT and FMT.
- Baseline ADHD vs. baseline control, to assess potential group differences at baseline measures.
- Post ADHD vs. post control, assessing group differences in source activity after the acquisition of the motor paradigm.
- (2)
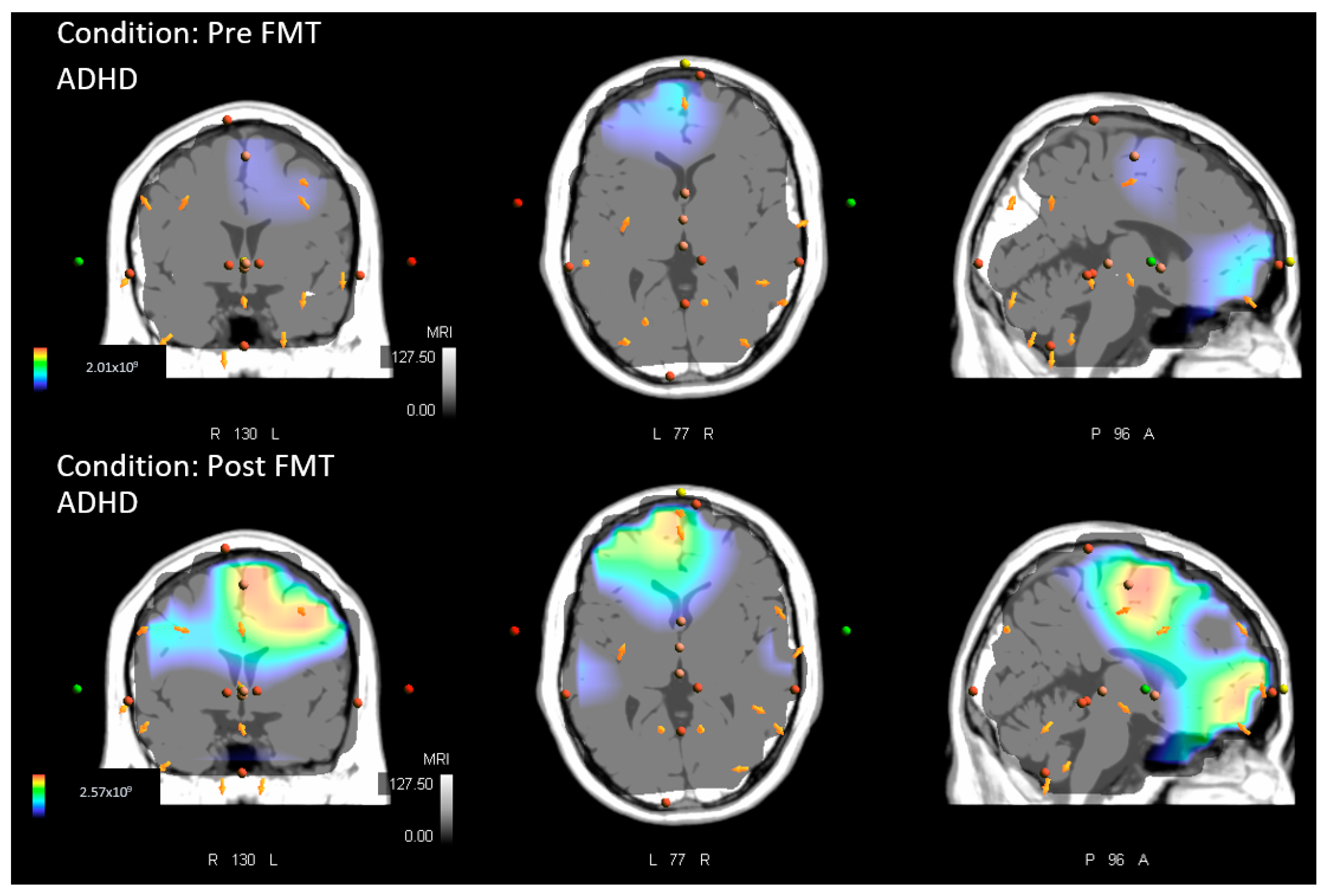
- Within groups (pre-measures vs. post-measures) for both the ADHD and control group. Similarly, comparisons were performed for both tasks, the MTT and FMT. This comparison was performed to assess whether locations of source activity differed within each group after acquisition of either of the motor paradigms.
- ADHD baseline vs. ADHD post.
- Control baseline vs. control post.
- (3)
- Finally, comparing between tasks, to assess whether somatosensory neural processing differed significantly between the two task conditions (MTT vs. FMT), discerning neural sources were activated in response to visuomotor vs. force-matching demands of each task, respectively.
- ADHD
- Baseline MTT vs. baseline FMT.
- Post MTT vs. post FMT.
- Control
- Baseline MTT vs. baseline FMT.
- Post MTT vs. post FMT.
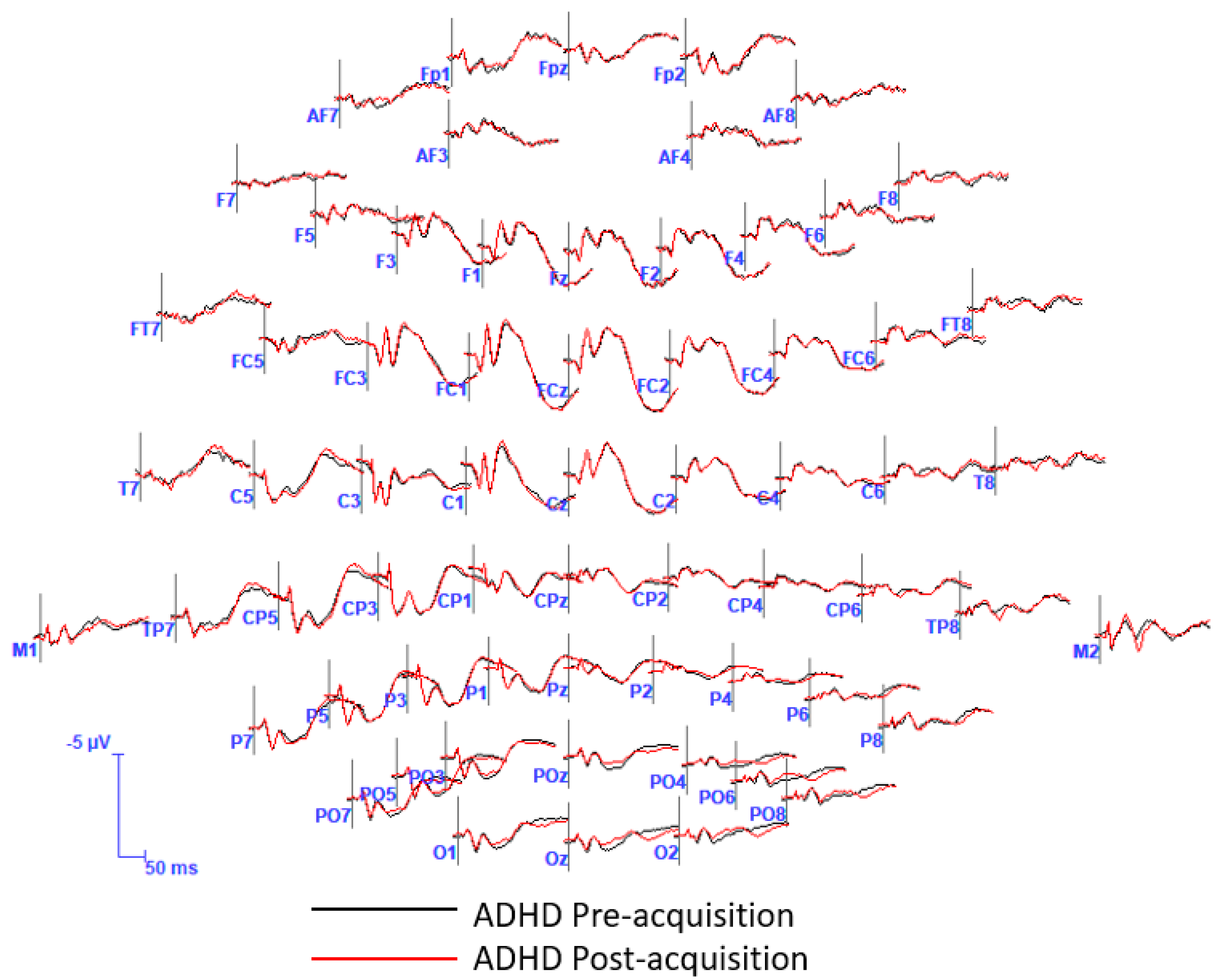
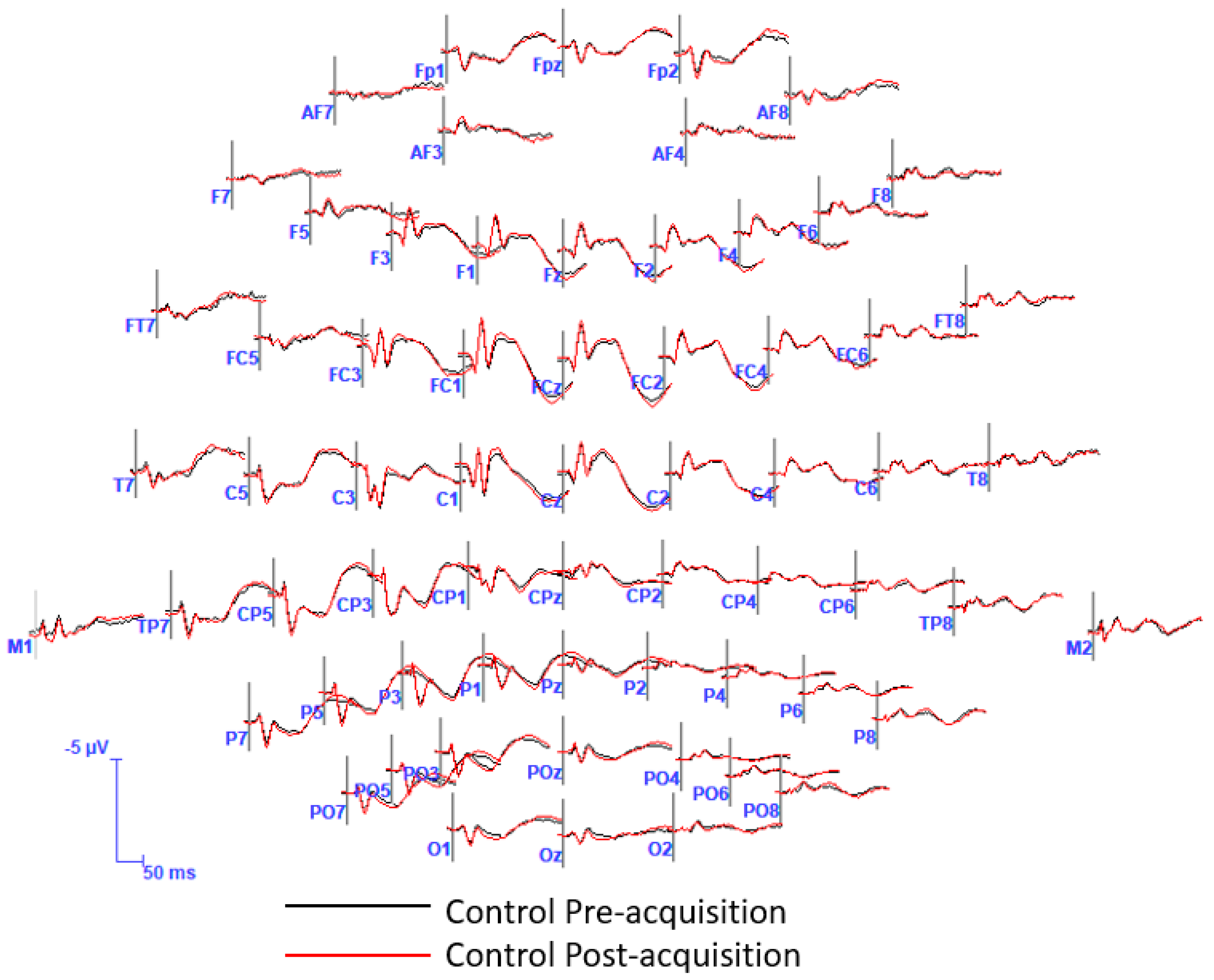
2.4.3. Statistical Analysis in the Time-Domain
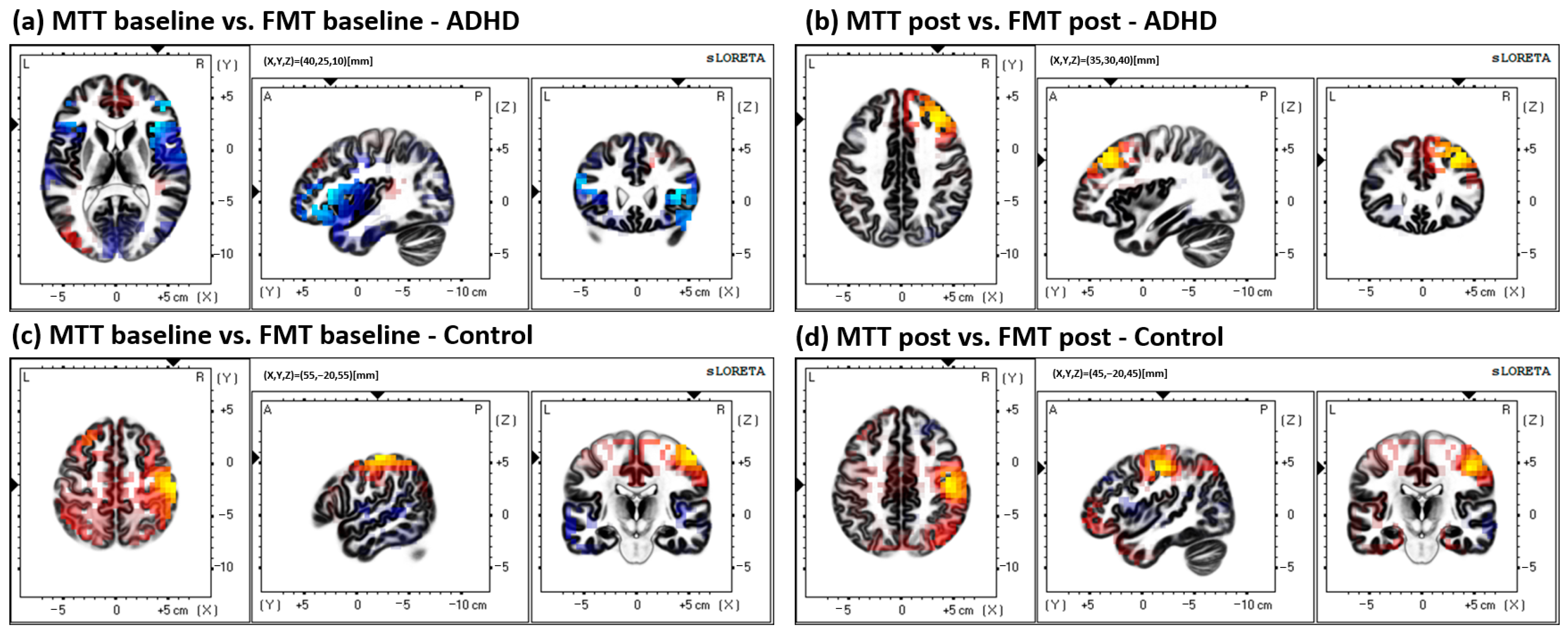
3. Results
4. Discussion
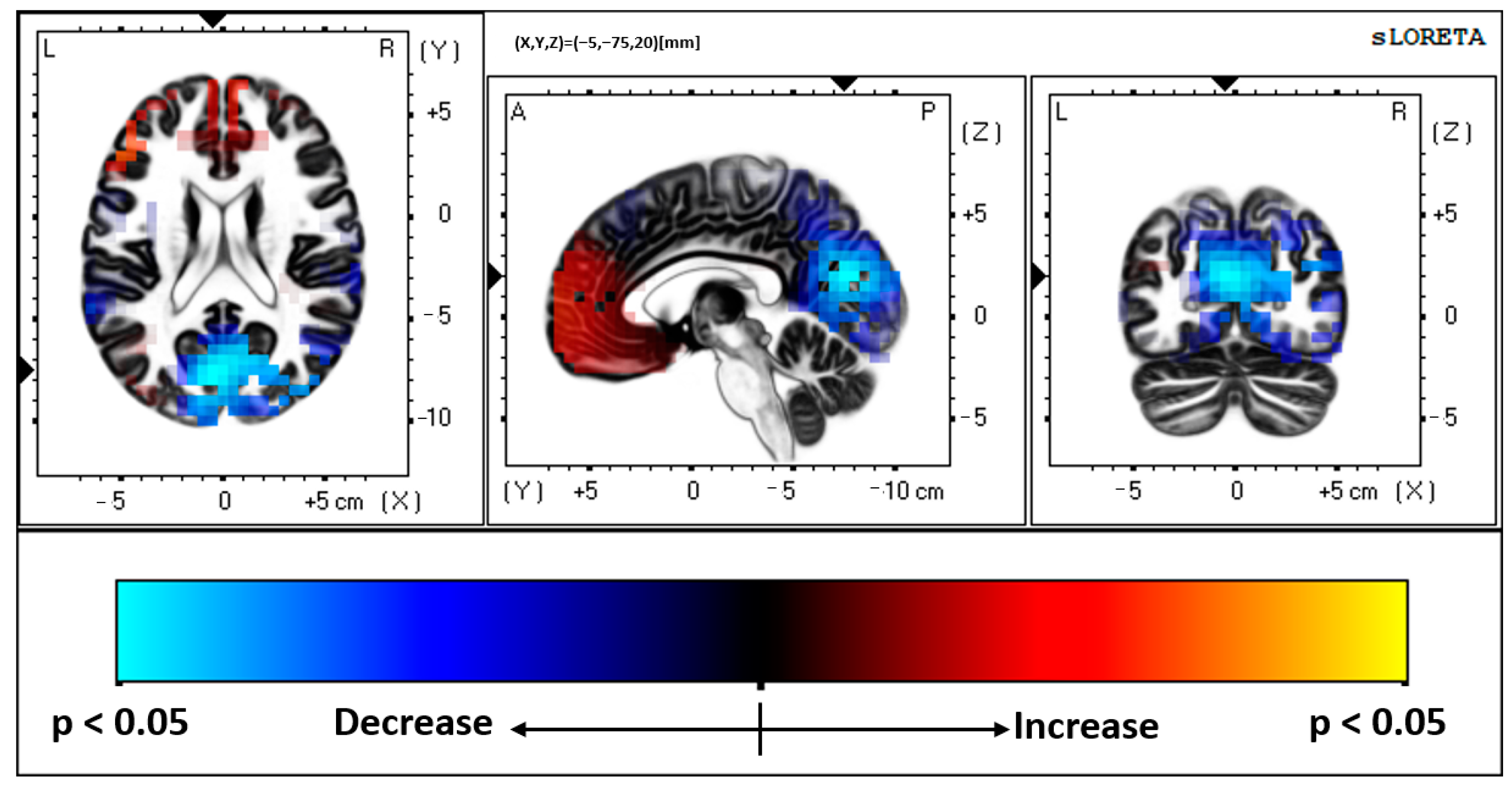
4.1. Brodmann Area (BA) 31
4.2. Latency
4.3. Limitations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A



References
- Visser, S.N.; Danielson, M.L.; Bitsko, R.H.; Holbrook, J.R.; Kogan, M.D.; Ghandour, R.M.; Perou, R.; Blumberg, S.J. Trends in the parent-report of health care provider-diagnosed and medicated attention-deficit/hyperactivity disorder: United States, 2003–2011. J. Am. Acad. Child Adolesc. Psychiatry 2014, 53, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders: DSM-5, 5th ed.; American Psychiatric Association: Washington, DC, USA, 2013; Volume 5. [Google Scholar]
- McAlonan, G.M.; Cheung, V.; Cheung, C.; Chua, S.E.; Murphy, D.G.; Suckling, J.; Tai, K.-S.; Yip, L.K.; Leung, P.; Ho, T.P. Mapping brain structure in attention deficit-hyperactivity disorder: A voxel-based MRI study of regional grey and white matter volume. Psychiatry Res. Neuroimaging 2007, 154, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Castellanos, F.X.; Lee, P.P.; Sharp, W.; Jeffries, N.O.; Greenstein, D.K.; Clasen, L.S.; Blumenthal, J.D.; James, R.S.; Ebens, C.L.; Walter, J.M.; et al. Developmental Trajectories of Brain Volume Abnormalities in Children and Adolescents with Attention-Deficit/Hyperactivity Disorder. JAMA 2002, 288, 1740–1748. [Google Scholar] [CrossRef] [Green Version]
- Proal, E.; Reiss, P.T.; Klein, R.G.; Mannuzza, S.; Gotimer, K.; Ramos-Olazagasti, M.A.; Lerch, J.P.; He, Y.; Zijdenbos, A.; Kelly, C. Brain gray matter deficits at 33-year follow-up in adults with attention-deficit/hyperactivity disorder established in childhood. Arch. Gen. Psychiatry 2011, 68, 1122–1134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castellanos, F.X.; Margulies, D.S.; Kelly, C.; Uddin, L.Q.; Ghaffari, M.; Kirsch, A.; Shaw, D.; Shehzad, Z.; Di Martino, A.; Biswal, B. Cingulate-precuneus interactions: A new locus of dysfunction in adult attention-deficit/hyperactivity disorder. Biol. Psychiatry 2008, 63, 332–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ehlis, A.-C.; Bähne, C.G.; Jacob, C.P.; Herrmann, M.J.; Fallgatter, A.J. Reduced lateral prefrontal activation in adult patients with attention-deficit/hyperactivity disorder (ADHD) during a working memory task: A functional near-infrared spectroscopy (fNIRS) study. J. Psychiatr. Res. 2008, 42, 1060–1067. [Google Scholar] [CrossRef] [PubMed]
- Liston, C.; Cohen, M.M.; Teslovich, T.; Levenson, D.; Casey, B. Atypical prefrontal connectivity in attention-deficit/hyperactivity disorder: Pathway to disease or pathological end point? Biol. Psychiatry 2011, 69, 1168–1177. [Google Scholar] [CrossRef] [PubMed]
- Makris, N.; Buka, S.L.; Biederman, J.; Papadimitriou, G.M.; Hodge, S.M.; Valera, E.M.; Brown, A.B.; Bush, G.; Monuteaux, M.C.; Caviness, V.S. Attention and executive systems abnormalities in adults with childhood ADHD: A DT-MRI study of connections. Cereb. Cortex 2008, 18, 1210–1220. [Google Scholar] [CrossRef] [Green Version]
- Sidlauskaite, J.; Caeyenberghs, K.; Sonuga-Barke, E.; Roeyers, H.; Wiersema, J.R. Whole-brain structural topology in adult attention-deficit/hyperactivity disorder: Preserved global–disturbed local network organization. NeuroImage Clin. 2015, 9, 506–512. [Google Scholar] [CrossRef]
- Sun, L.; Cao, Q.; Long, X.; Sui, M.; Cao, X.; Zhu, C.; Zuo, X.; An, L.; Song, Y.; Zang, Y. Abnormal functional connectivity between the anterior cingulate and the default mode network in drug-naïve boys with attention deficit hyperactivity disorder. Psychiatry Res. Neuroimaging 2012, 201, 120–127. [Google Scholar] [CrossRef]
- Krain, A.L.; Castellanos, F.X. Brain development and ADHD. Clin. Psychol. Rev. 2006, 26, 433–444. [Google Scholar] [CrossRef] [PubMed]
- Koziol, L.F.; Budding, D.E.; Chidekel, D. The Development of Motor Skills, Executive Functions, and a Relationship to ADHD: A Preliminary Review. In ADHD as a Model of Brain-Behavior Relationships; Springer: Berlin/Heidelberg, Germany, 2013; pp. 57–62. [Google Scholar]
- Durston, S.; van Belle, J.; de Zeeuw, P. Differentiating frontostriatal and fronto-cerebellar circuits in attention-deficit/hyperactivity disorder. Biol. Psychiatry 2011, 69, 1178–1184. [Google Scholar] [CrossRef] [PubMed]
- Mackie, S.; Shaw, P.; Lenroot, R.; Pierson, R.; Greenstein, D.K.; Nugent, T.F., III; Sharp, W.S.; Giedd, J.N.; Rapoport, J.L. Cerebellar development and clinical outcome in attention deficit hyperactivity disorder. Am. J. Psychiatry 2007, 164, 647–655. [Google Scholar] [CrossRef] [PubMed]
- Makris, N.; Liang, L.; Biederman, J.; Valera, E.M.; Brown, A.B.; Petty, C.; Spencer, T.J.; Faraone, S.V.; Seidman, L.J. Toward defining the neural substrates of ADHD: A controlled structural MRI study in medication-naive adults. J. Atten. Disord. 2015, 19, 944–953. [Google Scholar] [CrossRef]
- Duerden, E.G.; Tannock, R.; Dockstader, C. Altered cortical morphology in sensorimotor processing regions in adolescents and adults with attention-deficit/hyperactivity disorder. Brain Res. 2012, 1445, 82–91. [Google Scholar] [CrossRef]
- Faraone, S.; Biederman, J.; Mick, E. The age-dependent decline of attention deficit hyperactivity disorder: A meta-analysis of follow-up studies. Psychol. Med. 2006, 36, 159–165. [Google Scholar] [CrossRef]
- Adi-Japha, E.; Fox, O.; Karni, A. Atypical acquisition and atypical expression of memory consolidation gains in a motor skill in young female adults with ADHD. Res. Dev. Disabil. 2011, 32, 1011–1020. [Google Scholar] [CrossRef]
- McCracken, H.; Murphy, B.; Ambalavanar, U.; Zabihhosseinian, M.; Yielder, P. Sensorimotor Integration and Motor Learning During a Novel Visuomotor Tracing Task in Young Adults with Attention-Deficit/Hyperactivity Disorder. J. Neurophysiol. 2022, 129, 247–261. [Google Scholar] [CrossRef]
- McCracken, H.S.; Murphy, B.A.; Ambalavanar, U.; Glazebrook, C.M.; Yielder, P.C. Sensorimotor Integration and Motor Learning During a Novel Force-Matching Task in Young Adults with Attention-Deficit/Hyperactivity Disorder. Front. Hum. Neurosci. 2022, 16, 877. [Google Scholar] [CrossRef]
- Depue, B.E.; Burgess, G.C.; Bidwell, L.C.; Willcutt, E.G.; Banich, M.T. Behavioral performance predicts grey matter reductions in the right inferior frontal gyrus in young adults with combined type ADHD. Psychiatry Res. Neuroimaging 2010, 182, 231–237. [Google Scholar] [CrossRef] [Green Version]
- Makris, N.; Biederman, J.; Valera, E.M.; Bush, G.; Kaiser, J.; Kennedy, D.N.; Caviness, V.S.; Faraone, S.V.; Seidman, L.J. Cortical Thinning of the Attention and Executive Function Networks in Adults with Attention-Deficit/Hyperactivity Disorder. Cereb. Cortex 2007, 17, 1364–1375. [Google Scholar] [CrossRef] [Green Version]
- Neely, K.A.; Chennavasin, A.P.; Yoder, A.; Williams, G.K.; Loken, E.; Huang-Pollock, C.L. Memory-guided force output is associated with self-reported ADHD symptoms in young adults. Exp. Brain Res. 2016, 234, 3203–3212. [Google Scholar] [CrossRef] [Green Version]
- Valera, E.M.; Spencer, R.M.; Zeffiro, T.A.; Makris, N.; Spencer, T.J.; Faraone, S.V.; Biederman, J.; Seidman, L.J. Neural substrates of impaired sensorimotor timing in adult attention-deficit/hyperactivity disorder. Biol. Psychiatry 2010, 68, 359–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brook, J.S.; Brook, D.W.; Zhang, C.; Seltzer, N.; Finch, S.J. Adolescent ADHD and adult physical and mental health, work performance, and financial stress. Pediatrics 2013, 131, 5–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, D.; Cherbuin, N.; Butterworth, P.; Anstey, K.J.; Easteal, S. A population-based study of attention deficit/hyperactivity disorder symptoms and associated impairment in middle-aged adults. PLoS ONE 2012, 7, e31500. [Google Scholar] [CrossRef] [Green Version]
- Biederman, J.; Faraone, S.V. The effects of attention-deficit/hyperactivity disorder on employment and household income. Medscape Gen. Med. 2006, 8, 12. [Google Scholar]
- Kessler, R.C.; Adler, L.; Ames, M.; Barkley, R.A.; Birnbaum, H.; Greenberg, P.; Johnston, J.A.; Spencer, T.; Üstün, T.B. The prevalence and effects of adult attention deficit/hyperactivity disorder on work performance in a nationally representative sample of workers. J. Occup. Environ. Med. 2005, 47, 565–572. [Google Scholar] [CrossRef] [Green Version]
- Rösler, M.; Casas, M.; Konofal, E.; Buitelaar, J. Attention deficit hyperactivity disorder in adults. World J. Biol. Psychiatry 2010, 11, 684–698. [Google Scholar] [CrossRef]
- Dockstader, C.; Gaetz, W.; Cheyne, D.; Tannock, R. Abnormal neural reactivity to unpredictable sensory events in attention-deficit/hyperactivity disorder. Biol. Psychiatry 2009, 66, 376–383. [Google Scholar] [CrossRef]
- Dockstader, C.; Gaetz, W.; Cheyne, D.; Wang, F.; Castellanos, F.X.; Tannock, R. MEG event-related desynchronization and synchronization deficits during basic somatosensory processing in individuals with ADHD. Behav. Brain Funct. 2008, 4, 8. [Google Scholar] [CrossRef] [Green Version]
- Toplak, M.; Tannock, R. Tapping and anticipation performance in attention deficit hyperactivity disorder. Percept. Mot. Ski. 2005, 100, 659–675. [Google Scholar] [CrossRef] [PubMed]
- Rubia, K.; Noorloos, J.; Smith, A.; Gunning, B.; Sergeant, J. Motor timing deficits in community and clinical boys with hyperactive behavior: The effect of methylphenidate on motor timing. J. Abnorm. Child Psychol. 2003, 31, 301–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubia, K.; Taylor, A.; Taylor, E.; Sergeant, J.A. Synchronization, anticipation, and consistency in motor timing of children with dimensionally defined attention deficit hyperactivity behaviour. Percept. Mot. Ski. 1999, 89, 1237–1258. [Google Scholar] [CrossRef] [PubMed]
- Werry, J.S.; Minde, K.; Guzman, A.; Weiss, G.; Dogan, K.; Hoy, E. Studies on the hyperactive child-VII: Neurological status compared with neurotic and normal children. Am. J. Orthopsychiatry 1972, 42, 441. [Google Scholar] [CrossRef] [PubMed]
- Parush, S.; Sohmer, H.; Steinberg, A.; Kaitz, M. Somatosensory functioning in children with attention deficit hyperactivity disorder. Dev. Med. Child Neurol. 1997, 39, 464–468. [Google Scholar] [CrossRef] [PubMed]
- Parush, S.; Sohmer, H.; Steinberg, A.; Kaitz, M. Somatosensory function in boys with ADHD and tactile defensiveness. Physiol. Behav. 2007, 90, 553–558. [Google Scholar] [CrossRef]
- Tucha, O.; Mecklinger, L.; Laufkoetter, R.; Klein, H.; Walitza, S.; Lange, K.W. Methylphenidate-induced improvements of various measures of attention in adults with attention deficit hyperactivity disorder. J. Neural Transm. 2006, 113, 1575–1592. [Google Scholar] [CrossRef]
- Kaiser, M.-L.; Schoemaker, M.; Albaret, J.-M.; Geuze, R. What is the evidence of impaired motor skills and motor control among children with attention deficit hyperactivity disorder (ADHD)? Systematic review of the literature. Res. Dev. Disabil. 2015, 36, 338–357. [Google Scholar] [CrossRef]
- Fliers, E.A.; Franke, B.; Buitelaar, J.K. Motor problems in children with ADHD receive too little attention in clinical practice. Ned. Tijdschr. Voor Geneeskd. 2011, 155, A3559. [Google Scholar]
- Karatekin, C.; Markiewicz, S.W.; Siegel, M.A. A preliminary study of motor problems in children with attention-deficit/hyperactivity disorder. Percept. Mot. Ski. 2003, 97, 1267–1280. [Google Scholar] [CrossRef]
- Lijffijt, M.; Kenemans, J.L.; Verbaten, M.N.; van Engeland, H. A meta-analytic review of stopping performance in attention-deficit/hyperactivity disorder: Deficient inhibitory motor control? J. Abnorm. Psychol. 2005, 114, 216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duda, T.A.; Casey, J.E.; O’Brien, A.M.; Frost, N.; Phillips, A.M. Reduced graphomotor procedural learning in children and adolescents with ADHD. Hum. Mov. Sci. 2019, 65, 60–70. [Google Scholar] [CrossRef] [PubMed]
- Shorer, Z.; Becker, B.; Jacobi-Polishook, T.; Oddsson, L.; Melzer, I. Postural control among children with and without attention deficit hyperactivity disorder in single and dual conditions. Eur. J. Pediatr. 2012, 171, 1087–1094. [Google Scholar] [CrossRef] [PubMed]
- Klotz, J.M.; Johnson, M.D.; Wu, S.W.; Isaacs, K.M.; Gilbert, D.L. Relationship between reaction time variability and motor skill development in ADHD. Child Neuropsychol. 2012, 18, 576–585. [Google Scholar] [CrossRef] [Green Version]
- Klimkeit, E.I.; Mattingley, J.B.; Sheppard, D.M.; Lee, P.; Bradshaw, J.L. Motor preparation, motor execution, attention, and executive functions in attention deficit/hyperactivity disorder (ADHD). Child Neuropsychol. 2005, 11, 153–173. [Google Scholar] [CrossRef]
- Yan, J.H.; Thomas, J.R. Arm movement control: Differences between children with and without attention deficit hyperactivity disorder. Res. Q. Exerc. Sport 2002, 73, 10–18. [Google Scholar] [CrossRef]
- Andrew, D.; Yielder, P.; Haavik, H.; Murphy, B. The effects of subclinical neck pain on sensorimotor integration following a complex motor pursuit task. Exp. Brain Res. 2018, 236, 1–11. [Google Scholar] [CrossRef]
- Haavik, H.; Murphy, B. Selective changes in cerebellar-cortical processing following motor training. Exp. Brain Res. 2013, 231, 397–403. [Google Scholar] [CrossRef]
- Noël, P.; Ozaki, I.; Desmedt, J.E. Origin of N18 and P14 far-fields of median nerve somatosensory evoked potentials studied in patients with a brain-stem lesion. Electroencephalogr. Clin. Neurophysiol. 1996, 98, 167–170. [Google Scholar] [CrossRef]
- Rossi, S.; della Volpe, R.; Ginanneschi, F.; Ulivelli, M.; Bartalini, S.; Spidalieri, R.; Rossi, A. Early somatosensory processing during tonic muscle pain in humans: Relation to loss of proprioception and motor ‘defensive’strategies. Clin. Neurophysiol. 2003, 114, 1351–1358. [Google Scholar] [CrossRef]
- Sonoo, M. Anatomic origin and clinical application of the widespread N18 potential in median nerve somatosensory evoked potentials. J. Clin. Neurophysiol. 2000, 17, 258–268. [Google Scholar] [CrossRef] [PubMed]
- McCracken, H.S.; Murphy, B.A.; Ambalavanar, U.; Glazebrook, C.M.; Yielder, P.C. Source Localization of Audiovisual Multisensory Neural Generators in Young Adults with Attention-Deficit/Hyperactivity Disorder. Brain Sci. 2022, 12, 809. [Google Scholar] [CrossRef] [PubMed]
- Vance, A.; Silk, T.; Casey, M.; Rinehart, N.J.; Bradshaw, J.L.; Bellgrove, M.A.; Cunnington, R. Right parietal dysfunction in children with attention deficit hyperactivity disorder, combined type: A functional MRI study. Mol. Psychiatry 2007, 12, 826–832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corbetta, M.; Miezin, F.M.; Shulman, G.L.; Petersen, S.E. A PET study of visuospatial attention. J. Neurosci. 1993, 13, 1202–1226. [Google Scholar] [CrossRef]
- Nobre, A.C.; Sebestyen, G.N.; Gitelman, D.R.; Mesulam, M.M.; Frackowiak, R.; Frith, C.D. Functional localization of the system for visuospatial attention using positron emission tomography. Brain J. Neurol. 1997, 120, 515–533. [Google Scholar] [CrossRef] [Green Version]
- Gitelman, D.R.; Nobre, A.C.; Parrish, T.B.; LaBar, K.S.; Kim, Y.-H.; Meyer, J.R.; Mesulam, M.-M. A large-scale distributed network for covert spatial attention: Further anatomical delineation based on stringent behavioural and cognitive controls. Brain 1999, 122, 1093–1106. [Google Scholar] [CrossRef] [Green Version]
- Chan, E.; Mattingley, J.B.; Huang-Pollock, C.; English, T.; Hester, R.; Vance, A.; Bellgrove, M.A. Abnormal spatial asymmetry of selective attention in ADHD. J. Child Psychol. Psychiatry 2009, 50, 1064–1072. [Google Scholar] [CrossRef]
- Sur, M. Receptive fields of neurons in areas 3b and 1 of somatosensory cortex in monkeys. Brain Res. 1980, 198, 465–471. [Google Scholar] [CrossRef]
- Pascual-Marqui, R.D. Standardized low-resolution brain electromagnetic tomography (sLORETA): Technical details. Methods Find Exp. Clin. Pharm. 2002, 24, 5–12. [Google Scholar]
- Dankner, Y.; Shalev, L.; Carrasco, M.; Yuval-Greenberg, S. Prestimulus Inhibition of Saccades in Adults with and without Attention-Deficit/Hyperactivity Disorder as an Index of Temporal Expectations. Psychol. Sci. 2017, 28, 835–850. [Google Scholar] [CrossRef] [Green Version]
- Van de Glind, G.; van den Brink, W.; Koeter, M.W.J.; Carpentier, P.-J.; van Emmerik-van Oortmerssen, K.; Kaye, S.; Skutle, A.; Bu, E.-T.H.; Franck, J.; Konstenius, M.; et al. Validity of the Adult ADHD Self-Report Scale (ASRS) as a screener for adult ADHD in treatment seeking substance use disorder patients. Drug Alcohol Depend. 2013, 132, 587–596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gandevia, S.; Burke, D.; McKeon, B. The projection of muscle afferents from the hand to cerebral cortex in man. Brain 1984, 107, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Nuwer, M.R.; Aminoff, M.; Desmedt, J.; Eisen, A.A.; Goodin, D.; Matsuoka, S.; Mauguière, F.; Shibasaki, H.; Sutherling, W.; Vibert, J.-F. IFCN recommended standards for short latency somatosensory evoked potentials. Report of an IFCN committee. Electroencephalogr. Clin. Neurophysiol. 1994, 91, 6–11. [Google Scholar] [CrossRef] [PubMed]
- Andrew, D.; Haavik, H.; Dancey, E.; Yielder, P.; Murphy, B. Somatosensory evoked potentials show plastic changes following a novel motor training task with the thumb. Clin. Neurophysiol. 2015, 126, 575–580. [Google Scholar] [CrossRef]
- Holland, L.; Murphy, B.; Passmore, S.; Yielder, P. Differences in corticomotor excitability between hemispheres following performance of a novel motor training task. Neurosci. Biomed. Eng. 2017, 5, 116–125. [Google Scholar] [CrossRef]
- Fuchs, M.; Kastner, J.; Wagner, M.; Hawes, S.; Ebersole, J.S. A standardized boundary element method volume conductor model. Clin. Neurophysiol. 2002, 113, 702–712. [Google Scholar] [CrossRef] [Green Version]
- Jurcak, V.; Tsuzuki, D.; Dan, I. 10/20, 10/10, and 10/5 systems revisited: Their validity as relative head-surface-based positioning systems. NeuroImage 2007, 34, 1600–1611. [Google Scholar] [CrossRef] [PubMed]
- Pascual-Marqui, R.D.; Michel, C.M.; Lehmann, D. Low resolution electromagnetic tomography: A new method for localizing electrical activity in the brain. Int. J. Psychophysiol. 1994, 18, 49–65. [Google Scholar] [CrossRef]
- Sekihara, K.; Sahani, M.; Nagarajan, S.S. Localization bias and spatial resolution of adaptive and non-adaptive spatial filters for MEG source reconstruction. NeuroImage 2005, 25, 1056–1067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mulert, C.; Jäger, L.; Schmitt, R.; Bussfeld, P.; Pogarell, O.; Möller, H.-J.; Juckel, G.; Hegerl, U. Integration of fMRI and simultaneous EEG: Towards a comprehensive understanding of localization and time-course of brain activity in target detection. NeuroImage 2004, 22, 83–94. [Google Scholar] [CrossRef]
- Nichols, T.E.; Holmes, A.P. Nonparametric permutation tests for functional neuroimaging: A primer with examples. Hum. Brain Mapp. 2002, 15, 1–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navid, M.S.; Lelic, D.; Niazi, I.K.; Holt, K.; Mark, E.B.; Drewes, A.M.; Haavik, H. The effects of chiropractic spinal manipulation on central processing of tonic pain-a pilot study using standardized low-resolution brain electromagnetic tomography (sLORETA). Sci. Rep. 2019, 9, 6925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoesen, G.W.V.; Morecraft, R.J.; Vogt, B.A. Connections of the monkey cingulate cortex. In Neurobiology of Cingulate Cortex and Limbic Thalamus; Springer: Berlin/Heidelberg, Germany, 1993; pp. 249–284. [Google Scholar]
- Cavanna, A.E.; Trimble, M.R. The precuneus: A review of its functional anatomy and behavioural correlates. Brain 2006, 129, 564–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leech, R.; Braga, R.; Sharp, D.J. Echoes of the brain within the posterior cingulate cortex. J. Neurosci. 2012, 32, 215–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raichle, M.E.; MacLeod, A.M.; Snyder, A.Z.; Powers, W.J.; Gusnard, D.A.; Shulman, G.L. A default mode of brain function. Proc. Natl. Acad. Sci. USA 2001, 98, 676–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buckner, R.L.; Andrews-Hanna, J.R.; Schacter, D.L. The brain’s default network: Anatomy, function, and relevance to disease. Ann. N. Y. Acad. Sci. 2008, 1124, 1–38. [Google Scholar] [CrossRef] [Green Version]
- Sormaz, M.; Murphy, C.; Wang, H.-T.; Hymers, M.; Karapanagiotidis, T.; Poerio, G.; Margulies, D.S.; Jefferies, E.; Smallwood, J. Default mode network can support the level of detail in experience during active task states. Proc. Natl. Acad. Sci. USA 2018, 115, 9318–9323. [Google Scholar] [CrossRef] [Green Version]
- Kennedy, D.P.; Redcay, E.; Courchesne, E. Failing to deactivate: Resting functional abnormalities in autism. Proc. Natl. Acad. Sci. USA 2006, 103, 8275–8280. [Google Scholar] [CrossRef] [Green Version]
- Shulman, G.; Fiez, J.; Corbetta, M.; Buckner, R.; Meizin, F.; Raichle, M. Common blood flow changes across visual tasks: II. Decreases in cerebral cortex. J. Cogn. Neurosci. 1997, 9, 648–663. [Google Scholar] [CrossRef] [Green Version]
- Hahn, B.; Ross, T.J.; Stein, E.A. Cingulate activation increases dynamically with response speed under stimulus unpredictability. Cereb. Cortex 2007, 17, 1664–1671. [Google Scholar] [CrossRef] [Green Version]
- Tian, L.; Jiang, T.; Wang, Y.; Zang, Y.; He, Y.; Liang, M.; Sui, M.; Cao, Q.; Hu, S.; Peng, M. Altered resting-state functional connectivity patterns of anterior cingulate cortex in adolescents with attention deficit hyperactivity disorder. Neurosci. Lett. 2006, 400, 39–43. [Google Scholar] [CrossRef] [PubMed]
- Broyd, S.J.; Demanuele, C.; Debener, S.; Helps, S.K.; James, C.J.; Sonuga-Barke, E.J. Default-mode brain dysfunction in mental disorders: A systematic review. Neurosci. Biobehav. Rev. 2009, 33, 279–296. [Google Scholar] [CrossRef] [PubMed]
- Uddin, L.Q.; Clare Kelly, A.; Biswal, B.B.; Xavier Castellanos, F.; Milham, M.P. Functional connectivity of default mode network components: Correlation, anticorrelation, and causality. Hum. Brain Mapp. 2009, 30, 625–637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uddin, L.Q.; Kelly, A.C.; Biswal, B.B.; Margulies, D.S.; Shehzad, Z.; Shaw, D.; Ghaffari, M.; Rotrosen, J.; Adler, L.A.; Castellanos, F.X. Network homogeneity reveals decreased integrity of default-mode network in ADHD. J. Neurosci. Methods 2008, 169, 249–254. [Google Scholar] [CrossRef]
- Espenhahn, S.; Godfrey, K.J.; Kaur, S.; Ross, M.; Nath, N.; Dmitrieva, O.; McMorris, C.; Cortese, F.; Wright, C.; Murias, K. Tactile cortical responses and association with tactile reactivity in young children on the autism spectrum. Mol. Autism 2021, 12, 26. [Google Scholar] [CrossRef]
- Desmedt, J.E.; Robertson, D. Differential enhancement of early and late components of the cerebral somatosensory evoked potentials during forced-paced cognitive tasks in man. J. Physiol. 1977, 271, 761–782. [Google Scholar] [CrossRef]
- Hada, Y. Latency differences of N20, P40/N60, P100/N140 SEP components after stimulation of proximal and distal sites of the median nerve. Clin. EEG Neurosci. 2006, 37, 25–29. [Google Scholar] [CrossRef]
- Desmedt, J.E.; Tomberg, C. Mapping early somatosensory evoked potentials in selective attention: Critical evaluation of control conditions used for titrating by difference the cognitive P30, P40, P100 and N140. Electroencephalogr. Clin. Neurophysiol. Evoked Potentials Sect. 1989, 74, 321–346. [Google Scholar] [CrossRef]
- Michie, P.T. Selective attention effects on somatosensory event-related potentials. Ann. N. Y. Acad. Sci. 1984, 425, 250–255. [Google Scholar] [CrossRef]
- Giabbiconi, C.M.; Dancer, C.; Zopf, R.; Gruber, T.; Müller, M.M. Selective spatial attention to left or right hand flutter sensation modulates the steady-state somatosensory evoked potential. Cogn. Brain Res. 2004, 20, 58–66. [Google Scholar] [CrossRef]
- Forschack, N.; Nierhaus, T.; Müller, M.M.; Villringer, A. Dissociable neural correlates of stimulation intensity and detection in somatosensation. NeuroImage 2020, 217, 116908. [Google Scholar] [CrossRef]
- Schubert, R.; Blankenburg, F.; Lemm, S.; Villringer, A.; Curio, G. Now you feel it—Now you don’t: ERP correlates of somatosensory awareness. Psychophysiology 2006, 43, 31–40. [Google Scholar] [CrossRef]
- Ueno, T.; Hada, Y.; Shimizu, Y.; Yamada, T. Relationship between somatosensory event-related potential N140 aberrations and hemispatial agnosia in patients with stroke: A preliminary study. Int. J. Neurosci. 2018, 128, 487–494. [Google Scholar] [CrossRef] [Green Version]
- Josiassen, R.C.; Shagass, C.; Roemer, R.A.; Ercegovac, D.V.; Straumanis, J.J. Somatosensory evoked potential changes with a selective attention task. Psychophysiology 1982, 19, 146–159. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, Y.; Imamura, N. Relationships between attention effects and intensity effects on the cognitive N140 and P300 components of somatosensory ERPs. Clin. Neurophysiol. 2000, 111, 1711–1718. [Google Scholar] [CrossRef]
- Kida, T.; Kakigi, R. High temporal resolution neuroimaging of attentional and somatosensory-motor processing in the human brain. Curr. Med. Imaging 2008, 4, 144–162. [Google Scholar] [CrossRef]
- Nakata, H.; Sakamoto, K.; Kakigi, R. The relationship between reaction time and response variability and somatosensory No-go potentials. Eur. J. Appl. Physiol. 2012, 112, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Nakata, H.; Sakamoto, K.; Honda, Y.; Kakigi, R. Temporal dynamics of neural activity in motor execution and inhibition processing. Eur. J. Neurosci. 2015, 41, 1448–1458. [Google Scholar] [CrossRef] [PubMed]
- Leroy, A.; Petit, G.; Zarka, D.; Cebolla, A.M.; Palmero-Soler, E.; Strul, J.; Dan, B.; Verbanck, P.; Chéron, G. EEG dynamics and neural generators in implicit navigational image processing in adults with ADHD. Neuroscience 2018, 373, 92–105. [Google Scholar]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
McCracken, H.S.; Murphy, B.A.; Ambalavanar, U.; Glazebrook, C.M.; Yielder, P.C. Source Localization of Somatosensory Neural Generators in Adults with Attention-Deficit/Hyperactivity Disorder. Brain Sci. 2023, 13, 370. https://doi.org/10.3390/brainsci13020370
McCracken HS, Murphy BA, Ambalavanar U, Glazebrook CM, Yielder PC. Source Localization of Somatosensory Neural Generators in Adults with Attention-Deficit/Hyperactivity Disorder. Brain Sciences. 2023; 13(2):370. https://doi.org/10.3390/brainsci13020370
Chicago/Turabian StyleMcCracken, Heather S., Bernadette A. Murphy, Ushani Ambalavanar, Cheryl M. Glazebrook, and Paul C. Yielder. 2023. "Source Localization of Somatosensory Neural Generators in Adults with Attention-Deficit/Hyperactivity Disorder" Brain Sciences 13, no. 2: 370. https://doi.org/10.3390/brainsci13020370