
Neck Muscle Vibration Alters Upper Limb Proprioception as Demonstrated by Changes in Accuracy and Precision during an Elbow Repositioning Task

Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
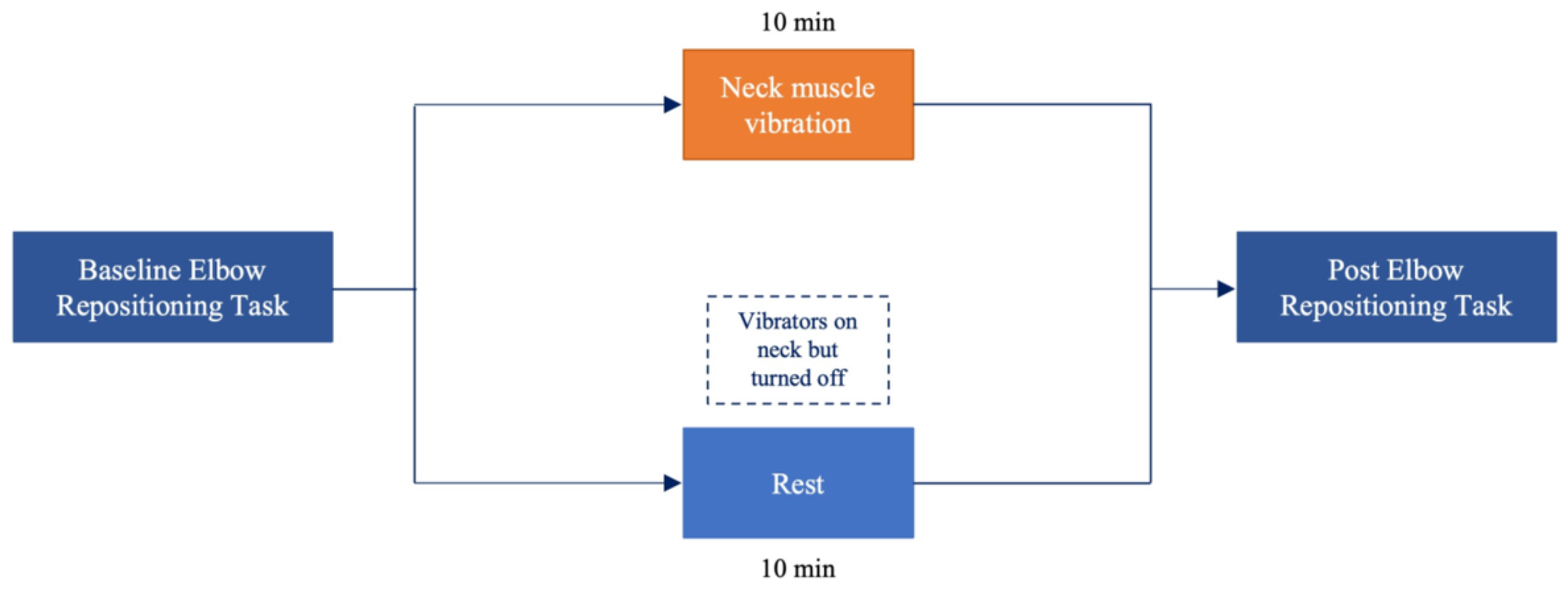
2.2. Elbow Repositioning Task (ERT)
2.3. Neck Muscle Vibration
2.4. Experimental Procedure
2.5. Data Processing
2.6. Statistical Analysis
3. Results
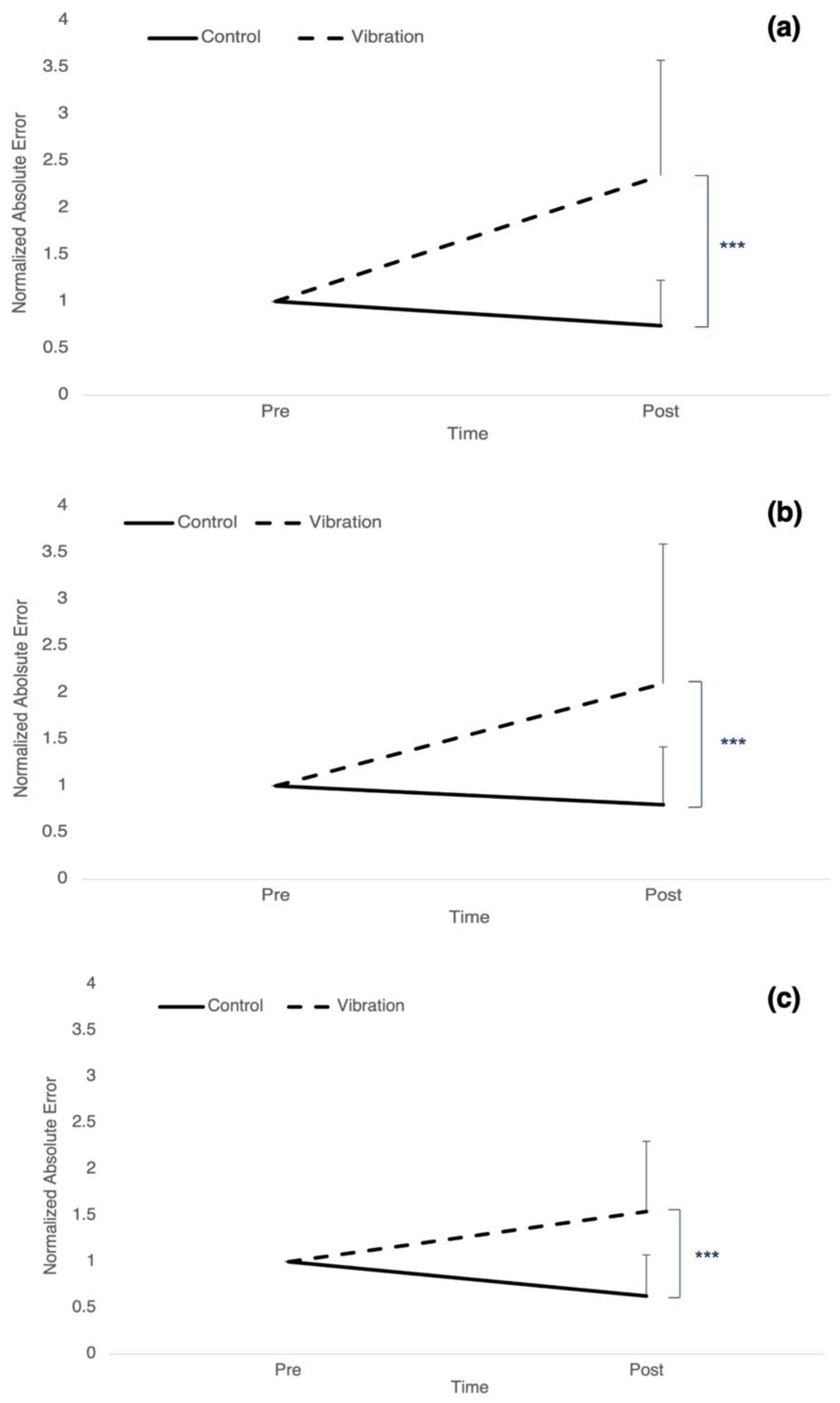
3.1. Accuracy: Absolute Error
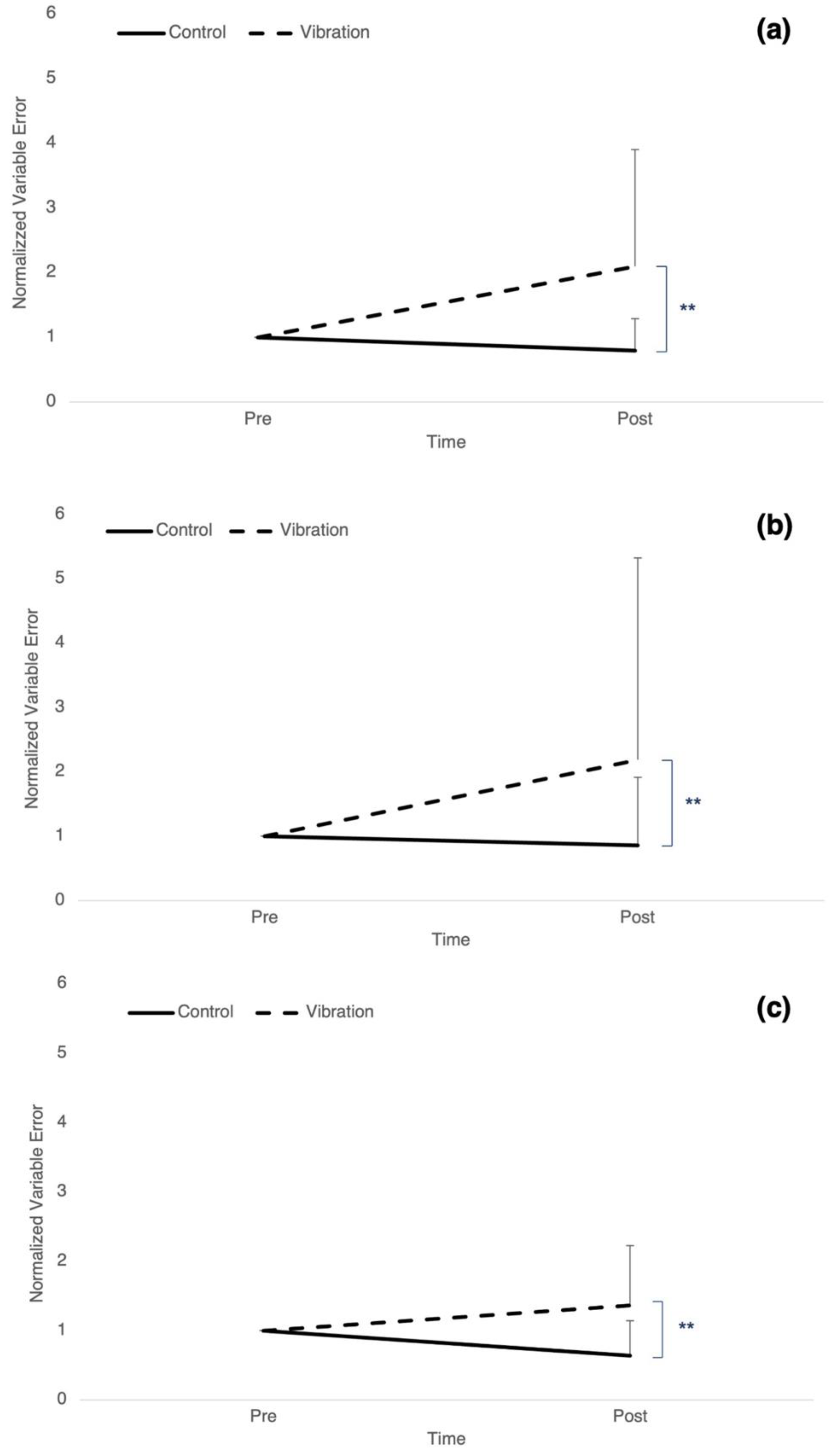
3.2. Precision: Variable Error
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Paulus, I.; Brumagne, S. Altered interpretation of neck proprioceptive signals in persons with subclinical recurrent neck pain. J. Rehabil. Med. 2008, 40, 426–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strimpakos, N.; Sakellari, V.; Gioftsos, G.; Kapreli, E.; Oldham, J. Cervical joint position sense: An intra- and inter-examiner reliability study. Gait Posture 2006, 23, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Proske, U.; Gandevia, S.C. The Proprioceptive Senses: Their Roles in Signaling Body Shape, Body Position and Movement, and Muscle Force. Physiol. Rev. 2012, 92, 1651–1697. [Google Scholar] [CrossRef] [PubMed]
- Richmond, F.; Abrahams, V. What Are the Proprioceptors of the Neck? Prog. Brain Res. 1979, 50, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Cooper, S.; Daniel, P.M. Muscle spindles in man; their morphology in the lumbricals and the deep muscles of the neck. Brain 1963, 86, 563–586. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, V.; Chandy, M.J.; Babu, K.S. Quantitative study of muscle spindles in suboccipital muscles of human foetuses. Neurol. India 2001, 49, 355. [Google Scholar]
- Holmes, N.P.; Spence, C. The body schema and multisensory representation(s) of peripersonal space. Cogn. Process. 2004, 5, 94–105. [Google Scholar] [CrossRef] [Green Version]
- Medina, J.; Coslett, H.B. From maps to form to space: Touch and the body schema. Neuropsychologia 2010, 48, 645–654. [Google Scholar] [CrossRef] [Green Version]
- Maravita, A.; Spence, C.; Driver, J. Multisensory integration and the body schema: Close to hand and within reach. Curr. Biol. 2003, 13, R531–R539. [Google Scholar] [CrossRef] [Green Version]
- Baarbé, J. Neck pain participants show impaired ability to perform a mental rotation task in a four week longitudinal study as compared to healthy controls. J. Chiropr. Med. 2015, 14, 99–100. [Google Scholar]
- Stanton, T.R.; Leake, H.B.; Chalmers, J.; Moseley, L. Evidence of Impaired Proprioception in Chronic, Idiopathic Neck Pain: Systematic Review and Meta-Analysis. Phys. Ther. 2016, 96, 876–887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wallwork, S.; Leake, H.B.; Peek, A.; Moseley, G.L.; Stanton, T.R. Implicit motor imagery performance is impaired in people with chronic, but not acute, neck pain. PeerJ 2020, 8, e8553. [Google Scholar] [CrossRef]
- Elsig, S.; Luomajoki, H.; Sattelmayer, M.; Taeymans, J.; Tal-Akabi, A.; Hilfiker, R. Sensorimotor tests, such as movement control and laterality judgment accuracy, in persons with recurrent neck pain and controls. A case-control study. Man. Ther. 2014, 19, 555–561. [Google Scholar] [CrossRef] [PubMed]
- Andrew, D.; Yielder, P.; Haavik, H.; Murphy, B. The effects of subclinical neck pain on sensorimotor integration following a complex motor pursuit task. Exp. Brain Res. 2017, 236, 1–11. [Google Scholar] [CrossRef]
- Farid, B.; Yielder, P.; Holmes, M.; Haavik, H.; Murphy, B.A. Association of Subclinical Neck Pain With Altered Multisensory Integration at Baseline and 4-Week Follow-up Relative to Asymptomatic Controls. J. Manip. Physiol. Ther. 2018, 41, 81–91. [Google Scholar] [CrossRef]
- Karellas, A.M.; Yielder, P.; Burkitt, J.J.; McCracken, H.S.; Murphy, B.A. The Influence of Subclinical Neck Pain on Neurophysiological and Behavioral Measures of Multisensory Integration. Brain Sci. 2019, 9, 362. [Google Scholar] [CrossRef] [Green Version]
- Zabihhosseinian, M.; Holmes, M.W.R.; Murphy, B. Neck muscle fatigue alters upper limb proprioception. Exp. Brain Res. 2015, 233, 1663–1675. [Google Scholar] [CrossRef] [PubMed]
- Zabihhosseinian, M.; Yielder, P.; Holmes, M.W.; Murphy, B. Neck muscle fatigue affects performance of an eye-hand tracking task. J. Electromyogr. Kinesiol. 2019, 47, 1–9. [Google Scholar] [CrossRef]
- Guerraz, M.; Blouin, J.; Vercher, J.-L. From head orientation to hand control: Evidence of both neck and vestibular involvement in hand drawing. Exp. Brain Res. 2003, 150, 40–49. [Google Scholar] [CrossRef]
- Knox, J.J.; Hodges, P.W. Changes in head and neck position affect elbow joint position sense. Exp. Brain Res. 2005, 165, 107–113. [Google Scholar] [CrossRef]
- Guerraz, M.; Caudron, S.; Thomassin, N.; Blouin, J. Influence of head orientation on visually and memory-guided arm movements. Acta Psychol. 2011, 136, 390–398. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.C.; Engberg, I.; Matthews, P.B.C. The relative sensitivity to vibration of muscle receptors of the cat. J. Physiol. 1967, 192, 773–800. [Google Scholar] [CrossRef] [PubMed]
- Goodwin, G.M.; McCloskey, D.I.; Matthews, P.B. Proprioceptive illusions induced by muscle vibration: Contribution by muscle spindles to perception? Science 1972, 175, 1382–1384. [Google Scholar] [CrossRef]
- Cordo, P.; Bevan, L.; Gurfinkel, V.; Carlton, L.; Carlton, M.; Kerr, G. Proprioceptive coordination of discrete movement sequences: Mechanism and generality. Can. J. Physiol. Pharmacol. 1995, 73, 305–315. [Google Scholar] [CrossRef] [PubMed]
- Knox, J.; Cordo, P.; Skoss, R.; Durrant, S.; Hodges, P. Illusory changes in head position induced by neck muscle vibration can alter the perception of elbow position. Behav. Neurosci. 2006, 120, 1211–1217. [Google Scholar] [CrossRef]
- Pettorossi, V.E.; Panichi, R.; Botti, F.M.; Biscarini, A.; Filippi, G.M.; Schieppati, M. Long-lasting effects of neck muscle vibration and contraction on self-motion perception of vestibular origin. Clin. Neurophysiol. 2015, 126, 1886–1900. [Google Scholar] [CrossRef]
- Tsay, A.; Allen, T.J.; Proske, U. Position sense at the human elbow joint measured by arm matching or pointing. Exp. Brain Res. 2016, 234, 2787–2798. [Google Scholar] [CrossRef]
- Chen, B.; Allen, T.; Proske, U. Position sense at the human forearm over a range of elbow angles. Exp. Brain Res. 2021, 239, 675–686. [Google Scholar] [CrossRef] [PubMed]
- Herter, T.M.; Scott, S.H.; Dukelow, S.P. Systematic changes in position sense accompany normal aging across adulthood. J. Neuroeng. Rehabil. 2014, 11, 43. [Google Scholar] [CrossRef] [Green Version]
- Alcaraz-Clariana, S.; García-Luque, L.; Garrido-Castro, J.; Fernández-De-Las-Peñas, C.; Carmona-Pérez, C.; Rodrigues-De-Souza, D.; Alburquerque-Sendín, F. Paravertebral Muscle Mechanical Properties and Spinal Range of Motion in Patients with Acute Neck or Low Back Pain: A Case-Control Study. Diagnostics 2021, 11, 352. [Google Scholar] [CrossRef]
- Zabihhosseinian, M.; Holmes, M.W.; Ferguson, B.; Murphy, B. Neck muscle fatigue alters the cervical flexion relaxation ratio in sub-clinical neck pain patients. Clin. Biomech. 2015, 30, 397–404. [Google Scholar] [CrossRef] [PubMed]
- Zabihhosseinian, M.; Holmes, M.W.; Howarth, S.; Ferguson, B.; Murphy, B. Neck muscle fatigue differentially alters scapular and humeral kinematics during humeral elevation in subclinical neck pain participants versus healthy controls. J. Electromyogr. Kinesiol. 2017, 33, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Baarbé, J.K.; Yielder, P.; Haavik, H.; Holmes, M.; Murphy, B.A. Subclinical recurrent neck pain and its treatment impacts motor training-induced plasticity of the cerebellum and motor cortex. PLoS ONE 2018, 13, e0193413. [Google Scholar] [CrossRef]
- Zabihhosseinian, M.; Yielder, P.; Berkers, V.; Ambalavanar, U.; Holmes, M.; Murphy, B.A. Neck muscle fatigue impacts plasticity and sensorimotor integration in cerebellum and motor cortex in response to novel motor skill acquisition. J. Neurophysiol. 2020, 124, 844–855. [Google Scholar] [CrossRef] [PubMed]
- Vernon, H. The Neck Disability Index: State-of-the-art, 1991–2008. J. Manip. Physiol. Ther. 2008, 31, 491–502. [Google Scholar] [CrossRef] [PubMed]
- Cohen, J. The earth is round (p <.05): Rejoinder. Am. Psychol. 1995, 50, 1103. [Google Scholar]
- Huysmans, M.; Hoozemans, M.; van der Beek, A.; de Looze, M.; van Dieën, J. Position sense acuity of the upper extremity and tracking performance in subjects with non-specific neck and upper extremity pain and healthy controls. J. Rehabil. Med. 2010, 42, 876–883. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.-M.; Liau, J.-J.; Cheng, C.-K.; Tan, C.-M.; Shih, J.-T. Evaluation of shoulder proprioception following muscle fatigue. Clin. Biomech. 2003, 18, 843–847. [Google Scholar] [CrossRef]
- Freund, J.E.; Williams, F.J. Dictionary/Outline of Basic Statistics; Courier Corporation: Chelmsford, MA, USA, 1991. [Google Scholar]
- Stallings, W.M.; Gillmore, G.M. A note on “accuracy” and “precision”. J. Educ. Meas. 1971, 8, 127–129. [Google Scholar] [CrossRef]
- Janwantanakul, P.; Magarey, M.E.; Jones, M.A.; Dansie, B.R. Variation in shoulder position sense at mid and extreme range of motion. Arch. Phys. Med. Rehabil. 2001, 82, 840–844. [Google Scholar] [CrossRef]
- Lönn, J.; Crenshaw, A.G.; Djupsjöbacka, M.; Pedersen, J.; Johansson, H. Position sense testing: Influence of starting position and type of displacement. Arch. Phys. Med. Rehabil. 2000, 81, 592–597. [Google Scholar] [CrossRef]
- Grigg, P. Response of joint afferent neurons in cat medial articular nerve to active and passive movements of the knee. Brain Res. 1976, 118, 482–485. [Google Scholar] [CrossRef]
- Salo, P.T.; Tatton, W.G. Age-related loss of knee joint afferents in mice. J. Neurosci. Res. 1993, 35, 664–677. [Google Scholar] [CrossRef]
- Felten, D.L.; O’Banion, M.K.; Maida, M.E. Netter’s Atlas of Neuroscience; Elsevier Health Sciences: Amsterdam, The Netherlands, 2015. [Google Scholar]
- Noël, P.; Ozaki, I.; Desmedt, J.E. Origin of N18 and P14 far-fields of median nerve somatosensory evoked potentials studied in patients with a brain-stem lesion. Electroencephalogr. Clin. Neurophysiol. 1996, 98, 167–170. [Google Scholar] [CrossRef]
- Tabbert, H.; Ambalavanar, U.; Murphy, B. Effect of Neck Muscle Vibration on Upper Limb Sensorimotor Integration and Motor Performance. In Health Sciences; Ontario Tech University: Oshawa, ON, Canada, 2022. [Google Scholar]
- Miall, R.C.; Reckess, G.Z.; Imamizu, H. The cerebellum coordinates eye and hand tracking movements. Nat. Neurosci. 2001, 4, 638–644. [Google Scholar] [CrossRef]
- Miall, R.C.; Imamizu, H.; Miyauchi, S. Activation of the cerebellum in co-ordinated eye and hand tracking movements: An fMRI study. Exp. Brain Res. 2000, 135, 22–33. [Google Scholar] [CrossRef]
- Lundy-Ekman, L. Neuroscience Fundamentals for Rehabilitation, 5th ed.; Elsevier: North York, ON, Canada, 2018; p. 562. [Google Scholar]
- Kandel, E.R.; Schwartz, J.H.; Jessell, T.M.; Siegelbaum, S.; Hudspeth, A.J.; Mack, S. Principles of Neural Science; McGraw-Hill: New York, NY, USA, 2000; Volume 4. [Google Scholar]
- Ernst, M.O.; Banks, M.S. Humans integrate visual and haptic information in a statistically optimal fashion. Nature 2002, 415, 429–433. [Google Scholar] [CrossRef]
- Ernst, M.O.; Bülthoff, H. Merging the senses into a robust percept. Trends Cogn. Sci. 2004, 8, 162–169. [Google Scholar] [CrossRef]
- Treleaven, J.; Jull, G.; Sterling, M. Dizziness and unsteadiness following whiplash injury: Characteristic features and relationship with cervical joint position error. J. Rehabil. Med. 2003, 35, 36–43. [Google Scholar] [CrossRef]
- Cheng, C.-H.; Wang, J.-L.; Lin, J.-J.; Wang, S.-F.; Lin, K.-H. Position accuracy and electromyographic responses during head reposition in young adults with chronic neck pain. J. Electromyogr. Kinesiol. 2010, 20, 1014–1020. [Google Scholar] [CrossRef]
- Revel, M.; Andre-Deshays, C.; Minguet, M. Cervicocephalic kinesthetic sensibility in patients with cervical pain. Arch. Phys. Med. Rehabil. 1991, 72, 288–291. [Google Scholar] [PubMed]
- Krajnak, K. Health effects associated with occupational exposure to hand-arm or whole body vibration. J. Toxicol. Environ. Health Part B 2018, 21, 320–334. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Reported Illusion | Frequency | Percentage |
|---|---|---|
| Neck Extension | 5 | 0.38 |
| Neck Flexion | 1 | 0.08 |
| Right Rotation | 2 | 0.15 |
| Left Rotation | 3 | 0.23 |
| Left Lateral Flexion | 1 | 0.08 |
| No Illusion | 1 | 0.08 |
| Time | |||||||
|---|---|---|---|---|---|---|---|
| Pre | Post | ||||||
| Normalized Elbow Repositioning Accuracy | |||||||
| Target 1: 80–90° | |||||||
| Absolute error controls (%) | *** p ≤ 0.001 | 1 ± 0 | 0.74 ± 0.49 *** | ||||
| Absolute error vibration (%) | 1 ± 0 | 2.34 ± 1.23 *** | |||||
| Variable error controls (%) | ** p ≤ 0.01 | 1 ± 0 | 0.79 ± 0.49 ** | ||||
| Variable error vibration (%) | 1 ± 0 | 2.09 ± 1.80 ** | |||||
| Target 2: 90–100° | |||||||
| Absolute error controls (%) | *** p ≤ 0.001 | 1 ± 0 | 0.79 ± 0.62 *** | ||||
| Absolute error vibration (%) | 1 ± 0 | 2.09 ± 1.49 *** | |||||
| Variable error controls (%) | ** p ≤ 0.01 | 1 ± 0 | 0.86 ± 1.06 ** | ||||
| Variable error vibration (%) | 1 ± 0 | 2.19 ± 3.14 ** | |||||
| Target 3: 100–110° | |||||||
| Absolute error controls (%) | *** p ≤ 0.001 | 1 ± 0 | 0.63 ± 0.44 * | ||||
| Absolute error vibration (%) | 1 ± 0 | 1.54 ± 0.75 * | |||||
| Variable error controls (%) | ** p ≤ 0.01 | 1 ± 0 | 0.64 ± 0.51 | ||||
| Variable error vibration (%) | 1 ± 0 | 1.36 ± 0.86 | |||||
| Absolute Elbow Repositioning Accuracy | |||||||
| Target 1: 80–90° | |||||||
| Absolute error controls (%) | 4.13 ± 1.71 | 3.05 ± 1.35 | |||||
| Absolute error vibration (%) | 2.89 ± 1.59 | 6.79 ± 3.04 | |||||
| Variable error controls (%) | 6.42 ± 3.21 | 5.11 ± 2.53 | |||||
| Variable error vibration (%) | 4.96 ± 3.18 | 10.40 ± 5.66 | |||||
| Target 2: 90–100° | |||||||
| Absolute error controls (%) | 3.37 ± 1.82 | 2.68 ± 1.32 | |||||
| Absolute error vibration (%) | 2.52 ± 1.53 | 5.27 ± 1.24 | |||||
| Variable error controls (%) | 5.18 ± 2.86 | 4.44 ± 2.59 | |||||
| Variable error vibration (%) | 3.72 ± 2.85 | 8.15 ± 3.55 | |||||
| Target 3: 100–110° | |||||||
| Absolute error controls (%) | 3.45 ± 1.38 | 2.17 ± 1.02 | |||||
| Absolute error vibration (%) | 2.78 ± 1.63 | 4.28 ± 1.43 | |||||
| Variable error controls (%) | 5.42 ± 2.37 | 3.45 ± 1.76 | |||||
| Variable error vibration (%) | 4.85 ± 3.31 | 6.61 ± 3.01 | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tabbert, H.; Ambalavanar, U.; Murphy, B. Neck Muscle Vibration Alters Upper Limb Proprioception as Demonstrated by Changes in Accuracy and Precision during an Elbow Repositioning Task. Brain Sci. 2022, 12, 1532. https://doi.org/10.3390/brainsci12111532
Tabbert H, Ambalavanar U, Murphy B. Neck Muscle Vibration Alters Upper Limb Proprioception as Demonstrated by Changes in Accuracy and Precision during an Elbow Repositioning Task. Brain Sciences. 2022; 12(11):1532. https://doi.org/10.3390/brainsci12111532
Chicago/Turabian StyleTabbert, Hailey, Ushani Ambalavanar, and Bernadette Murphy. 2022. "Neck Muscle Vibration Alters Upper Limb Proprioception as Demonstrated by Changes in Accuracy and Precision during an Elbow Repositioning Task" Brain Sciences 12, no. 11: 1532. https://doi.org/10.3390/brainsci12111532