
Exploring the Temporal Patterns of Dynamic Information Flow during Attention Network Test (ANT)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
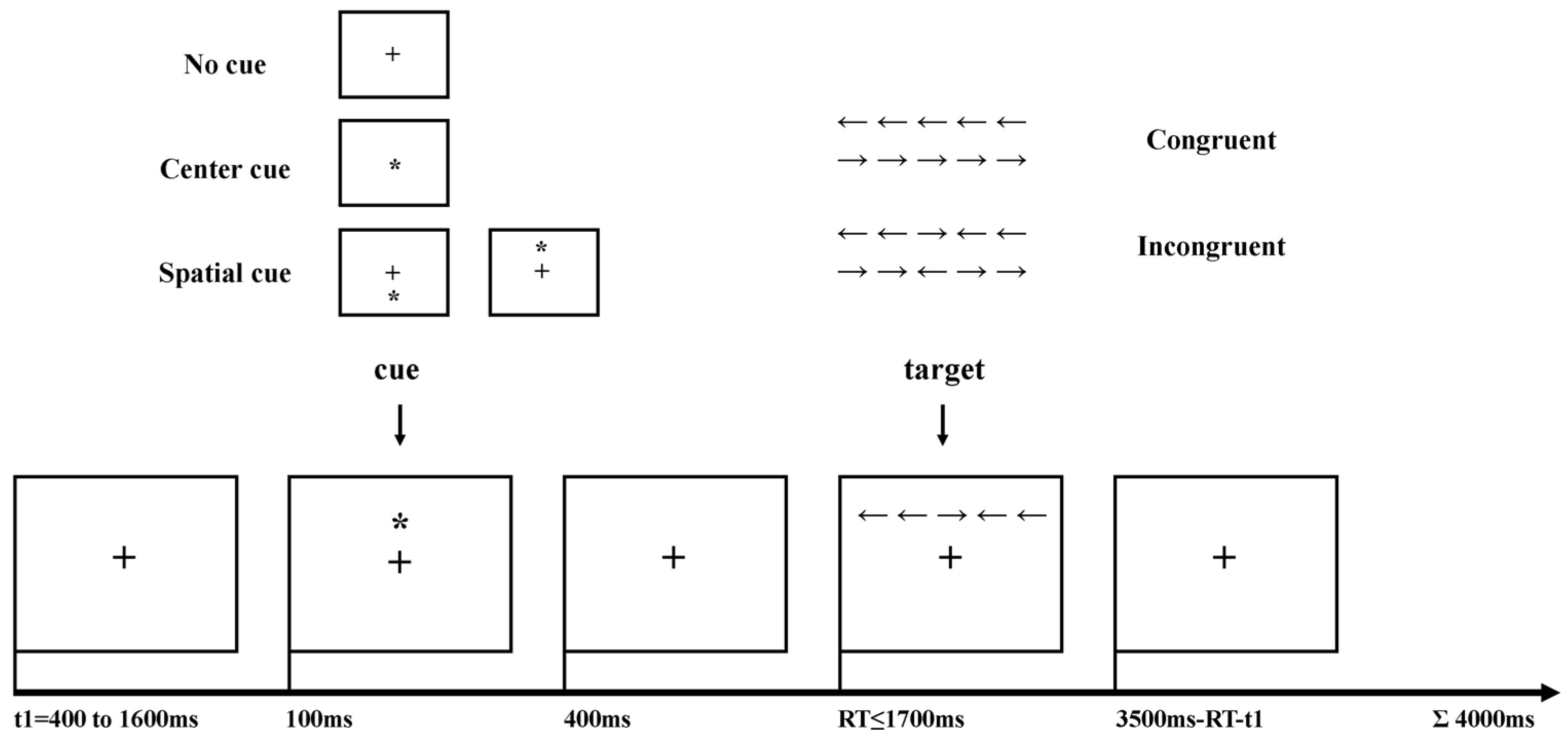
2.2. Experimental Procedures
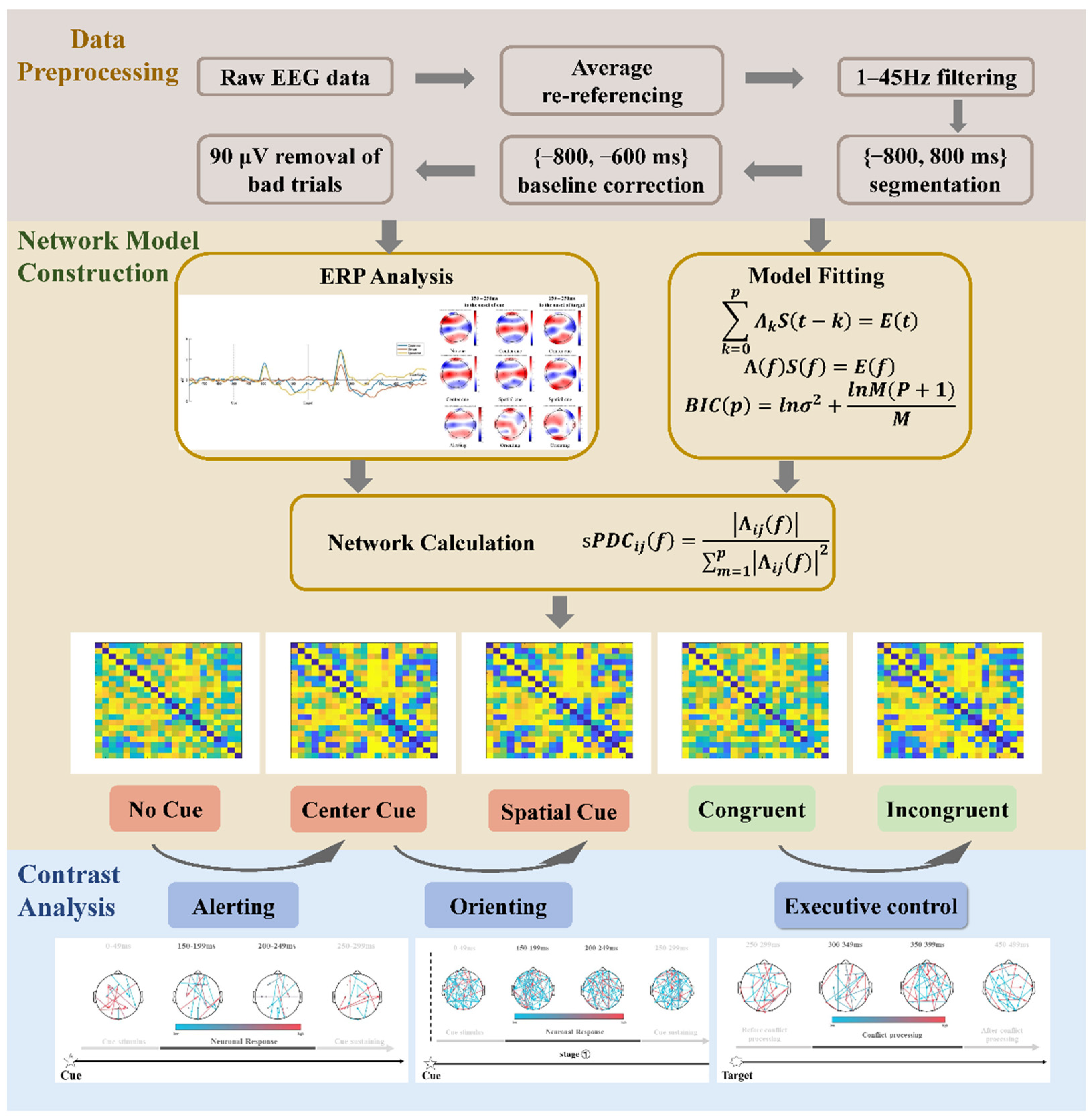
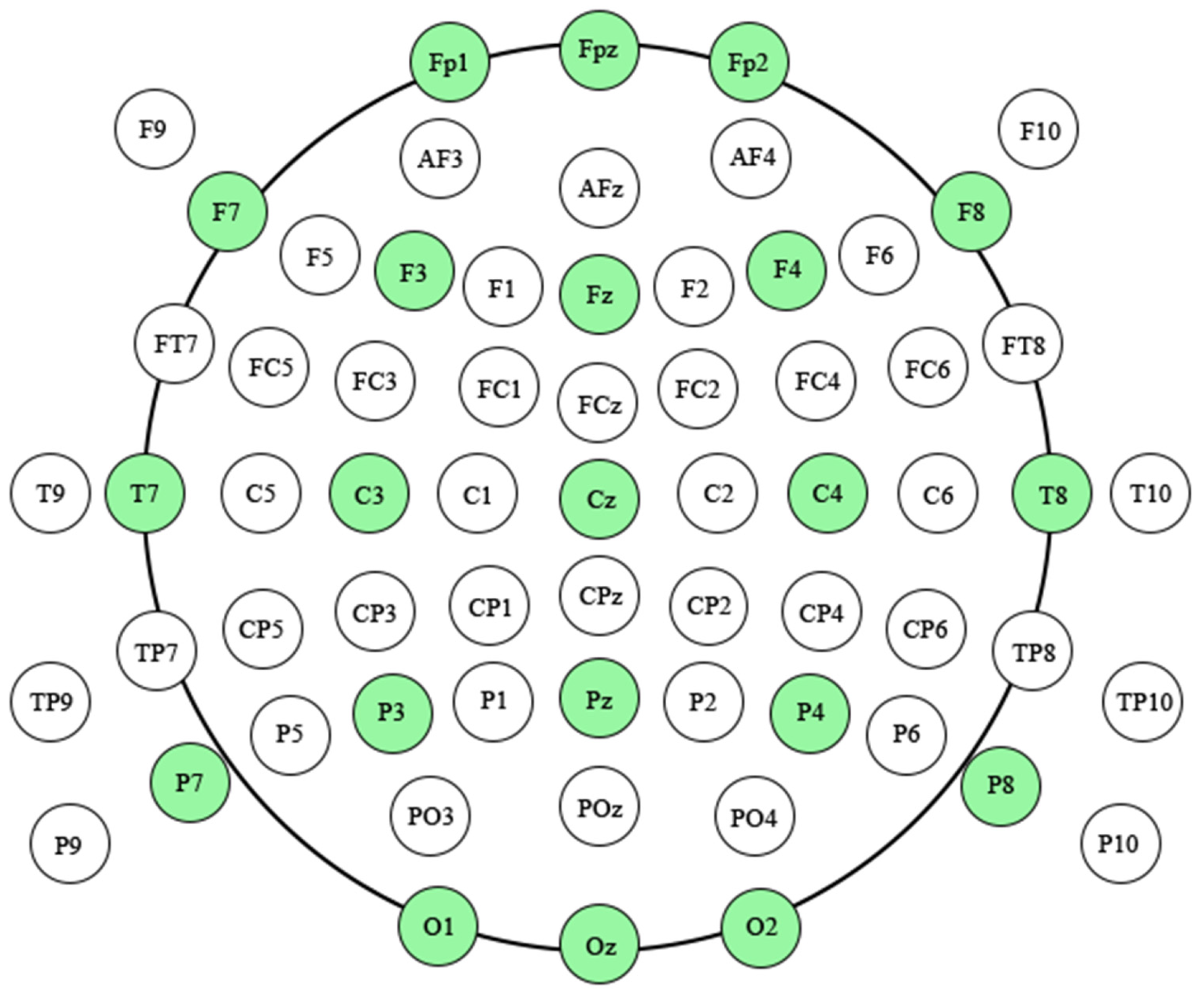
2.3. EEG Data Acquisition and ERP Construction
2.4. Fitting Model of EEG Signal
2.5. Dynamic Network Construction
2.6. The Sub-Stages of Attention Networks
2.7. Statistical Analysis
3. Results
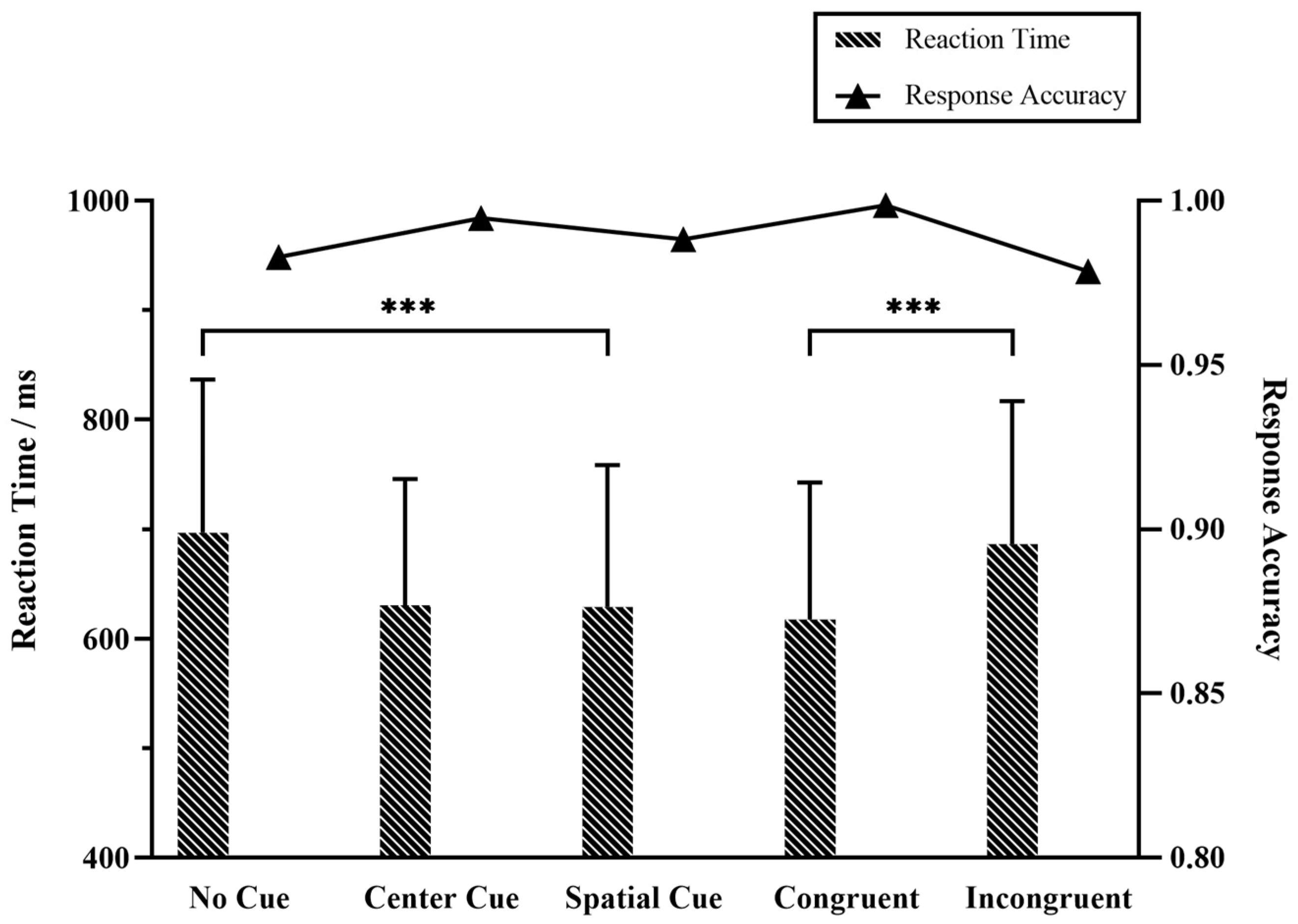
3.1. Behavior Performance
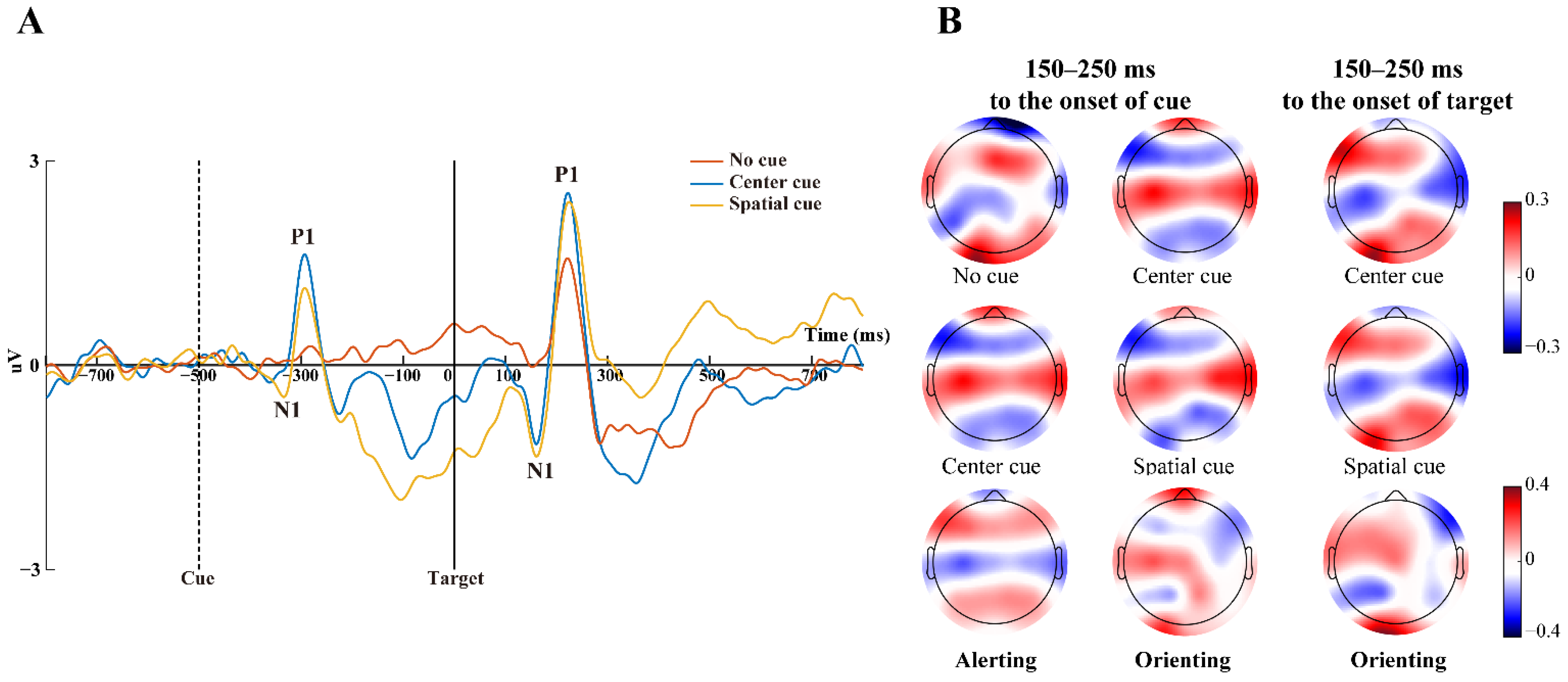
3.2. ERP Potentials Associated with Attention Networks
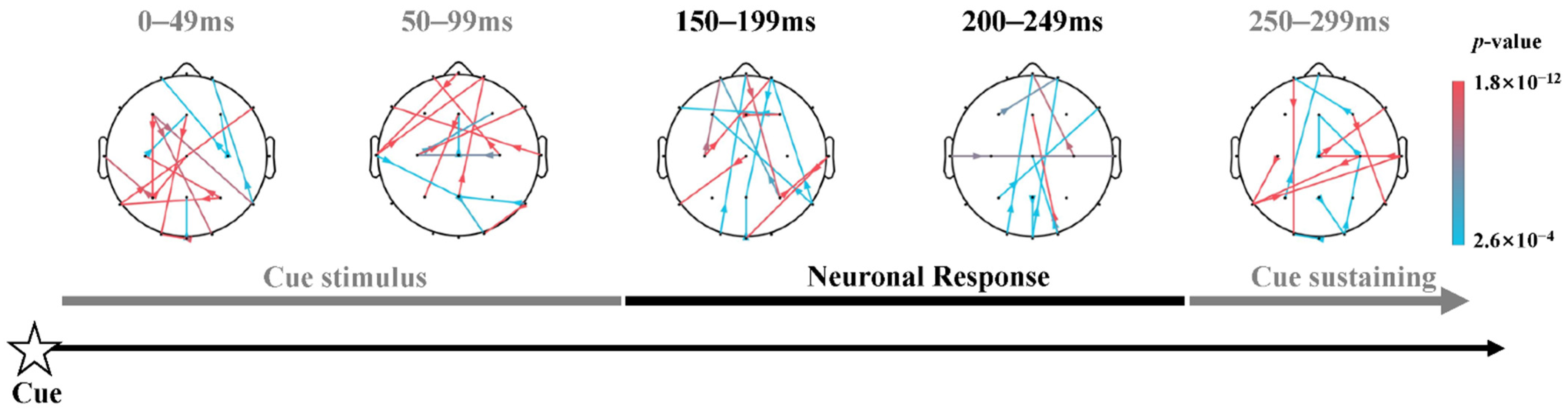
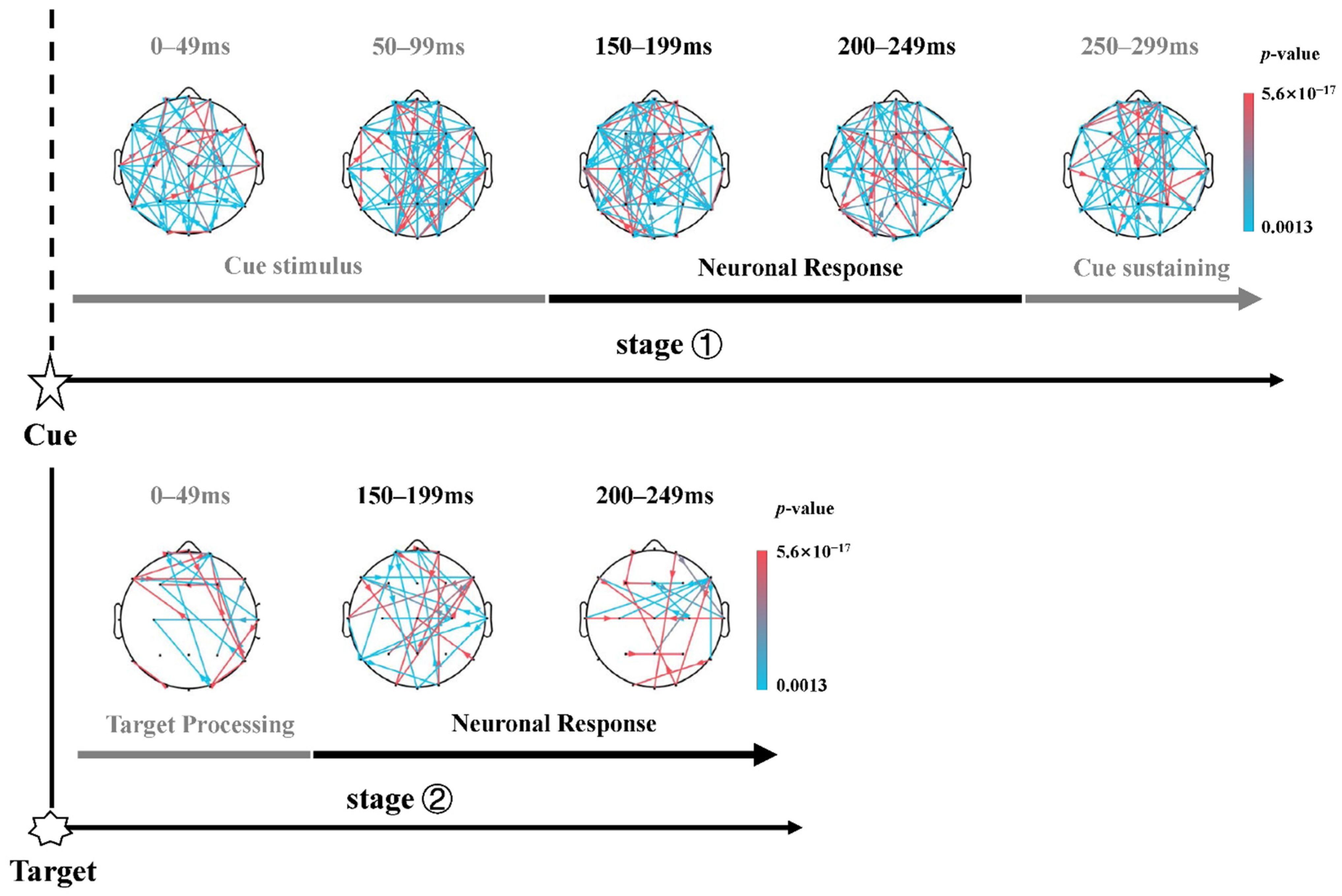
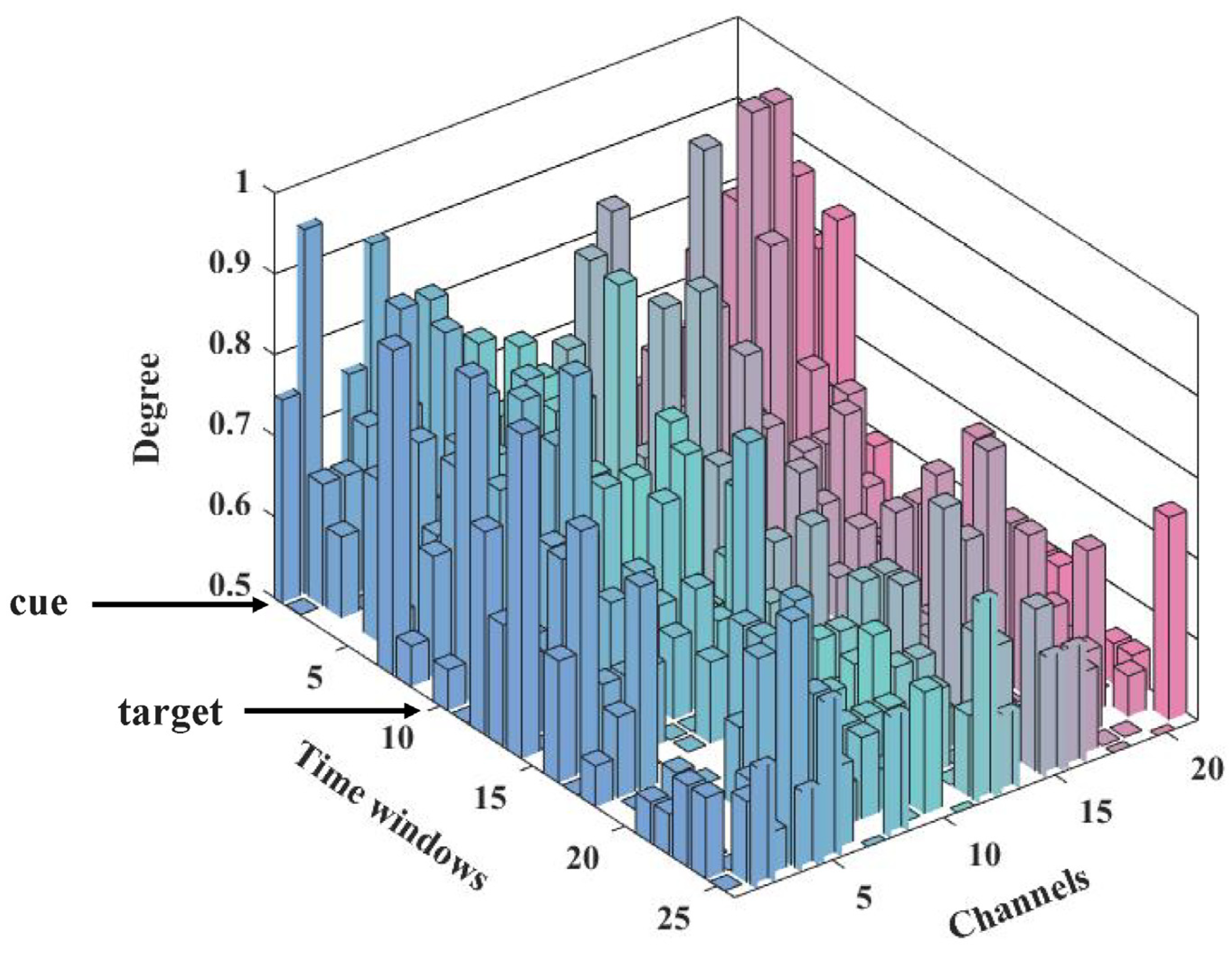
3.3. Dynamic Information Flow of Attention Networks
4. Discussion
4.1. The Role of ERP Potentials Associated with Each Attention Network
4.2. Temporal Patterns of Dynamic Information Flow in Alerting
4.3. Sparser Information Flow of Orienting Network after the Target Onset
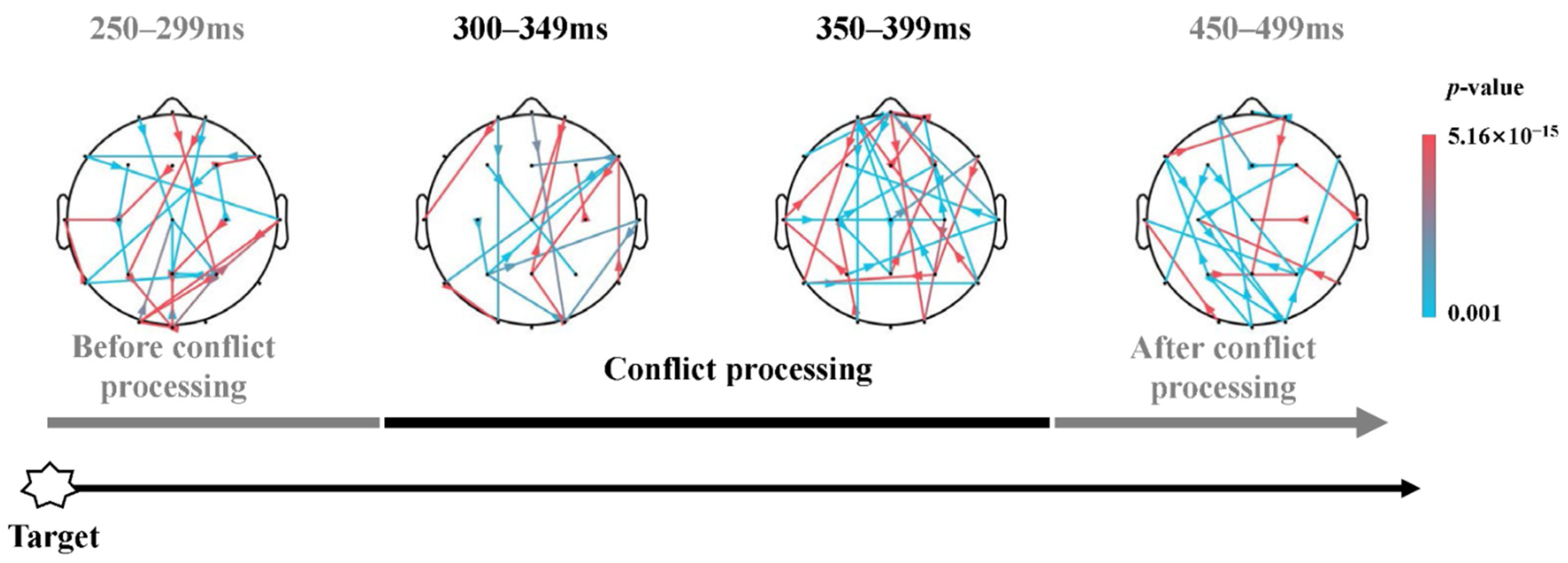
4.4. Top-Down Processes of Response Conflict
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cowan, N. An Embedded-Processes Model of Working Memory; Cambridge University Press: Cambridge, UK, 1999. [Google Scholar]
- Petersen, S.E.; Posner, M.I. The attention system of the human brain: 20 years after. Annu. Rev. Neurosci. 2012, 35, 73–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Posner, M.I. Imaging attention networks. NeuroImage 2012, 61, 450–456. [Google Scholar] [CrossRef] [Green Version]
- Posner, M.I.; Petersen, S.E. The attention system of the human brain. Annu. Rev. Neurosci. 1990, 13, 25–42. [Google Scholar] [CrossRef]
- Fan, J.; McCandliss, B.D.; Sommer, T.; Raz, A.; Posner, M.I. Testing the efficiency and independence of attentional networks. J. Cogn. Neurosci. 2002, 14, 340–347. [Google Scholar] [CrossRef]
- Fan, J.; McCandliss, B.D.; Fossella, J.; Flombaum, J.I.; Posner, M.I. The activation of attentional networks. NeuroImage 2005, 26, 471–479. [Google Scholar] [CrossRef]
- Markett, S.; Nothdurfter, D.; Focsa, A.; Reuter, M.; Jawinski, P. Attention networks and the intrinsic network structure of the human brain. Hum. Brain Mapp. 2022, 43, 1431–1448. [Google Scholar] [CrossRef]
- Sadaghiani, S.; D’Esposito, M. Functional Characterization of the Cingulo-Opercular Network in the Maintenance of Tonic Alertness. Cereb. Cortex 2015, 25, 2763–2773. [Google Scholar] [CrossRef] [Green Version]
- Power, J.D.; Cohen, A.L.; Nelson, S.M.; Wig, G.S.; Barnes, K.A.; Church, J.A.; Vogel, A.C.; Laumann, T.O.; Miezin, F.M.; Schlaggar, B.L.; et al. Functional network organization of the human brain. Neuron 2011, 72, 665–678. [Google Scholar] [CrossRef] [Green Version]
- Dosenbach, N.U.; Fair, D.A.; Miezin, F.M.; Cohen, A.L.; Wenger, K.K.; Dosenbach, R.A.; Fox, M.D.; Snyder, A.Z.; Vincent, J.L.; Raichle, M.E.; et al. Distinct brain networks for adaptive and stable task control in humans. Proc. Natl. Acad. Sci. USA 2007, 104, 11073–11078. [Google Scholar] [CrossRef] [Green Version]
- Fan, J.; Kolster, R.; Ghajar, J.; Suh, M.; Knight, R.T.; Sarkar, R.; McCandliss, B.D. Response anticipation and response conflict: An event-related potential and functional magnetic resonance imaging study. J. Neurosci. 2007, 27, 2272–2282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, J.; Byrne, J.; Worden, M.S.; Guise, K.G.; McCandliss, B.D.; Fossella, J.; Posner, M.I. The relation of brain oscillations to attentional networks. J. Neurosci. Off. J. Soc. Neurosci. 2007, 27, 6197–6206. [Google Scholar] [CrossRef]
- Neuhaus, A.H.; Urbanek, C.; Opgen-Rhein, C.; Hahn, E.; Ta, T.M.; Koehler, S.; Gross, M.; Dettling, M. Event-related potentials associated with Attention Network Test. Int. J. Psychophysiol. 2010, 76, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Neuhaus, A.H.; Popescu, F.C.; Grozea, C.; Hahn, E.; Hahn, C.; Opgen-Rhein, C.; Urbanek, C.; Dettling, M. Single-subject classification of schizophrenia by event-related potentials during selective attention. NeuroImage 2011, 55, 514–521. [Google Scholar] [CrossRef] [PubMed]
- Hopfinger, J.B.; West, V.M. Interactions between endogenous and exogenous attention on cortical visual processing. NeuroImage 2006, 31, 774–789. [Google Scholar] [CrossRef] [PubMed]
- Roberts, L.E.; Rau, H.; Lutzenberger, W.; Birbaumer, N. Mapping P300 waves onto inhibition: Go/No-Go discrimination. Electroencephalogr. Clin. Neurophysiol. 1994, 92, 44–55. [Google Scholar] [CrossRef]
- Bullmore, E.; Sporns, O. The economy of brain network organization. Nat. Rev. Neurosci. 2012, 13, 336–349. [Google Scholar] [CrossRef]
- Duan, X.; Xie, S.; Xie, X.; Obermayer, K.; Cui, Y.; Wang, Z. An Online Data Visualization Feedback Protocol for Motor Imagery-Based BCI Training. Front. Hum. Neurosci. 2021, 15, 625983. [Google Scholar] [CrossRef]
- Si, Y.; Wu, X.; Li, F.; Zhang, L.; Duan, K.; Li, P.; Song, L.; Jiang, Y.; Zhang, T.; Zhang, Y.; et al. Different Decision-Making Responses Occupy Different Brain Networks for Information Processing: A Study Based on EEG and TMS. Cereb. Cortex 2019, 29, 4119–4129. [Google Scholar] [CrossRef]
- Li, F.; Chen, B.; Li, H.; Zhang, T.; Wang, F.; Jiang, Y.; Li, P.; Ma, T.; Zhang, R.; Tian, Y.; et al. The Time-Varying Networks in P300: A Task-Evoked EEG Study. IEEE Trans. Neural. Syst. Rehabil. Eng. 2016, 24, 725–733. [Google Scholar] [CrossRef]
- Qi, P.; Hu, H.; Zhu, L.; Gao, L.; Yuan, J.; Thakor, N.; Bezerianos, A.; Sun, Y. EEG Functional Connectivity Predicts Individual Behavioural Impairment During Mental Fatigue. IEEE Trans. Neural. Syst. Rehabil. Eng. 2020, 28, 2080–2089. [Google Scholar] [CrossRef]
- Duan, K.; Wu, Q.; Liao, Y.; Si, Y.; Bore, J.C.; Li, F.; Tao, Q.; Lin, L.; Lei, W.; Hu, X.; et al. Discrimination of Tourette Syndrome Based on the Spatial Patterns of the Resting-State EEG Network. Brain Topogr. 2021, 34, 78–87. [Google Scholar] [CrossRef]
- Bowling, J.T.; Friston, K.J.; Hopfinger, J.B. Top-down versus bottom-up attention differentially modulate frontal-parietal connectivity. Hum. Brain Mapp. 2020, 41, 928–942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cross, N.; Paquola, C.; Pomares, F.B.; Perrault, A.A.; Jegou, A.; Nguyen, A.; Aydin, U.; Bernhardt, B.C.; Grova, C.; Dang-Vu, T.T. Cortical gradients of functional connectivity are robust to state-dependent changes following sleep deprivation. NeuroImage 2021, 226, 117547. [Google Scholar] [CrossRef]
- Uji, M.; Cross, N.; Pomares, F.B.; Perrault, A.A.; Jegou, A.; Nguyen, A.; Aydin, U.; Lina, J.M.; Dang-Vu, T.T.; Grova, C. Data-driven beamforming technique to attenuate ballistocardiogram artefacts in electroencephalography-functional magnetic resonance imaging without detecting cardiac pulses in electrocardiography recordings. Hum. Brain Mapp. 2021, 42, 3993–4021. [Google Scholar] [CrossRef]
- Faul, F.; Erdfelder, E.; Lang, A.-G.; Buchner, A. G*Power 3: A flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behav. Res. Methods 2007, 39, 175–191. [Google Scholar] [CrossRef]
- Nakao, T.; Takezawa, T.; Shiraishi, M.; Miyatani, M. Activation of self-knowledge reduces conflict during occupational choice: An ERP study. Int. J. Neurosci. 2009, 119, 1640–1654. [Google Scholar] [CrossRef]
- Zani, A.; Proverbio, A.M. How voluntary orienting of attention and alerting modulate costs of conflict processing. Sci. Rep. 2017, 7, 46701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delorme, A.; Mullen, T.; Kothe, C.; Akalin Acar, Z.; Bigdely-Shamlo, N.; Vankov, A.; Makeig, S. Eeglab, sift, nft, bcilab, and erica: New tools for advanced EEG processing. Comput. Intell. Neurosci. 2011, 2011, 130714. [Google Scholar] [CrossRef] [Green Version]
- Galvao-Carmona, A.; Gonzalez-Rosa, J.J.; Hidalgo-Munoz, A.R.; Paramo, D.; Benitez, M.L.; Izquierdo, G.; Vazquez-Marrufo, M. Disentangling the attention network test: Behavioral, event related potentials, and neural source analyses. Front. Hum. Neurosci. 2014, 8, 813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doricchi, F.; Pellegrino, M.; Marson, F.; Pinto, M.; Caratelli, L.; Cestari, V.; Rossi-Arnaud, C.; Lasaponara, S. Deconstructing Reorienting of Attention: Cue Predictiveness Modulates the Inhibition of the No-target Side and the Hemispheric Distribution of the P1 Response to Invalid Targets. J. Cogn. Neurosci. 2020, 32, 1046–1060. [Google Scholar] [CrossRef]
- Granger, C.W.J. Investigating Causal Relations by Econometric Models and Cross-Spectral Methods. Econometrica 1969, 37, 424–438. [Google Scholar] [CrossRef]
- Ding, M.; Bressler, S.L.; Yang, W.; Liang, H. Short-window spectral analysis of cortical event-related potentials by adaptive multivariate autoregressive modeling: Data preprocessing, model validation, and variability assessment. Biol. Cybern. 2000, 83, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Blinowska, K.J.; Kus, R.; Kaminski, M. Granger causality and information flow in multivariate processes. Phys. Rev. E Stat. Nonlin. Soft Matter. Phys. 2004, 70(Pt. 1), 050902. [Google Scholar] [CrossRef]
- Lütkepohl, H. New Introduction to Multiple Time Series Analysis, 1st ed.; Springer: Berlin/Heidelberg, Germany, 2005; 764p. [Google Scholar]
- Astolfi, L.; Cincotti, F.; Mattia, D.; De Vico Fallani, F.; Tocci, A.; Colosimo, A.; Salinari, S.; Marciani, M.G.; Hesse, W.; Witte, H.; et al. Tracking the Time-Varying Cortical Connectivity Patterns by Adaptive Multivariate Estimators. IEEE Trans. Biomed. Eng. 2008, 55, 902–913. [Google Scholar] [CrossRef] [PubMed]
- Niso, G.; Bruna, R.; Pereda, E.; Gutierrez, R.; Bajo, R.; Maestu, F.; del-Pozo, F. HERMES: Towards an integrated toolbox to characterize functional and effective brain connectivity. Neuroinformatics 2013, 11, 405–434. [Google Scholar] [CrossRef] [Green Version]
- Abramov, D.M.; Pontes, M.; Pontes, A.T.; Mourao-Junior, C.A.; Vieira, J.; Quero Cunha, C.; Tamborino, T.; Galhanone, P.R.; deAzevedo, L.C.; Lazarev, V.V. Visuospatial information processing load and the ratio between parietal cue and target P3 amplitudes in the Attentional Network Test. Neurosci. Lett. 2017, 647, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Mangun, G.R.; Hillyard, S.A. Modulations of Sensory-Evoked Brain Potentials Indicate Changes in Perceptual Processing During Visual-Spatial Priming. J. Exp. Psychol. Hum. Percept. Perform. 1991, 17, 1057–1074. [Google Scholar] [CrossRef]
- Slagter, H.A.; Prinssen, S.; Reteig, L.C.; Mazaheri, A. Facilitation and inhibition in attention: Functional dissociation of pre-stimulus alpha activity, P1, and N1 components. NeuroImage 2016, 125, 25–35. [Google Scholar] [CrossRef] [Green Version]
- Luna, F.G.; Marino, J.; Roca, J.; Lupianez, J. Executive and arousal vigilance decrement in the context of the attentional networks: The ANTI-Vea task. J. Neurosci. Methods 2018, 306, 77–87. [Google Scholar] [CrossRef]
- Luna, F.G.; Roca, J.; Martin-Arevalo, E.; Lupianez, J. Measuring attention and vigilance in the laboratory vs. online: The split-half reliability of the ANTI-Vea. Behav. Res. Methods 2020, 53, 1124–1147. [Google Scholar] [CrossRef]
- Bullmore, E.; Sporns, O. Complex brain networks: Graph theoretical analysis of structural and functional systems. Nat. Rev. Neurosci. 2009, 10, 186–198. [Google Scholar] [CrossRef]
- Sharp, D.J.; Scott, G.; Leech, R. Network dysfunction after traumatic brain injury. Nat. Rev. Neurol. 2014, 10, 156–166. [Google Scholar] [CrossRef]
- Tian, L.; Wang, J.; Yan, C.; He, Y. Hemisphere- and gender-related differences in small-world brain networks: A resting-state functional MRI study. NeuroImage 2011, 54, 191–202. [Google Scholar] [CrossRef]
- Bassett, D.S.; Bullmore, E. Small-World Brain Networks. Neuroscientist 2016, 12, 512–523. [Google Scholar] [CrossRef] [PubMed]
- Deiber, M.P.; Ibanez, V.; Missonnier, P.; Rodriguez, C.; Giannakopoulos, P. Age-associated modulations of cerebral oscillatory patterns related to attention control. NeuroImage 2013, 82, 531–546. [Google Scholar] [CrossRef]
- Proskovec, A.L.; Heinrichs-Graham, E.; Wiesman, A.I.; McDermott, T.J.; Wilson, T.W. Oscillatory dynamics in the dorsal and ventral attention networks during the reorienting of attention. Hum. Brain Mapp. 2018, 39, 2177–2190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dosenbach, N.U.; Visscher, K.M.; Palmer, E.D.; Miezin, F.M.; Wenger, K.K.; Kang, H.C.; Burgund, E.D.; Grimes, A.L.; Schlaggar, B.L.; Petersen, S.E. A core system for the implementation of task sets. Neuron 2006, 50, 799–812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trujillo, L.T.; Kornguth, S.; Schnyer, D.M. An ERP examination of the different effects of sleep deprivation on exogenously cued and endogenously cued attention. Sleep 2009, 32, 1285–1297. [Google Scholar] [CrossRef] [Green Version]
- Wascher, E.; Hoffmann, S.; Sanger, J.; Grosjean, M. Visuo-spatial processing and the N1 component of the ERP. Psychophysiology 2009, 46, 1270–1277. [Google Scholar] [CrossRef]
- Vogel, E.K.; Luck, S.J. The visual N1 component as an index of a discrimination process. Psychophysiology 2000, 37, 190–203. [Google Scholar] [CrossRef]
- Doniger, G.M.; Foxe, J.J.; Schroeder, C.E.; Murray, M.M.; Higgins, B.A.; Javitt, D.C. Visual perceptual learning in human object recognition areas: A repetition priming study using high-density electrical mapping. NeuroImage 2001, 13, 305–313. [Google Scholar] [CrossRef] [PubMed]
- O’Connell, R.G.; Schneider, D.; Hester, R.; Mattingley, J.B.; Bellgrove, M.A. Attentional load asymmetrically affects early electrophysiological indices of visual orienting. Cereb Cortex 2011, 21, 1056–1065. [Google Scholar] [CrossRef] [Green Version]
- Kilintari, M.; Bufacchi, R.J.; Novembre, G.; Guo, Y.; Haggard, P.; Iannetti, G.D. High-precision voluntary movements are largely independent of preceding vertex potentials elicited by sudden sensory events. J. Physiol. 2018, 596, 3655–3673. [Google Scholar] [CrossRef]
- Fleck, J.I.; Payne, L.; Halko, C.; Purcell, M. Should we pay attention to eye movements? The impact of bilateral eye movements on behavioral and neural responses during the Attention Network Test. Brain Cogn. 2019, 132, 56–71. [Google Scholar] [CrossRef] [PubMed]
- Senderecka, M. Emotional enhancement of error detection-The role of perceptual processing and inhibition monitoring in failed auditory stop trials. Cogn. Affect. Behav. Neurosci. 2018, 18, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Polich, J. Updating P300: An integrative theory of P3a and P3b. Clin. Neurophysiol. 2007, 118, 2128–2148. [Google Scholar] [CrossRef] [Green Version]
- Anzolin, A.; Mattia, D.b.; Toppi, J.; Pichiorri, F.; Riccio, A.; Astolfi, L. Brain connectivity networks at the basis of human attention components: An EEG study. In Proceedings of the Annual International Conference of the IEEE Engineering in Medicine and Biology Society, EMBS; 2017; pp. 3953–3956. [Google Scholar]
- Wang, C.; Wang, X.; Zhu, M.; Pi, Y.; Wang, X.; Wan, F.; Chen, S.; Li, G. Spectrum power and brain functional connectivity of different EEG frequency bands in attention network tests. Annu. Int. Conf. IEEE Eng. Med. Biol. Soc. 2021, 2021, 224–227. [Google Scholar] [CrossRef]
- Kempermann, G.; Gast, D.; Gage, F.H. Neuroplasticity in old age: Sustained fivefold induction of hippocampal neurogenesis by long-term environmental enrichment. Ann. Neurol. 2002, 52, 135–143. [Google Scholar] [CrossRef]
- Posner, M.I. Measuring alertness. Ann. N. Y. Acad. Sci. 2008, 1129, 193–199. [Google Scholar] [CrossRef]
- Robertson, I.H. A right hemisphere role in cognitive reserve. Neurobiol. Aging 2014, 35, 1375–1385. [Google Scholar] [CrossRef]
- He, Y.; Evans, A. Graph theoretical modeling of brain connectivity. Curr. Opin. Neurol. 2010, 23, 341–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, B.; Astolfi, L.; Valdes-Sosa, P.A.; Marinazzo, D.; Palva, S.; Benar, C.G.; Michel, C.M.; Koenig, T. Electrophysiological Brain Connectivity: Theory and Implementation. IEEE Trans. Bio-Med. Eng. 2019, 66, 2115–2137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Posner, M.I.; Sheese, B.E.; Odludas, Y.; Tang, Y. Analyzing and shaping human attentional networks. Neural. Netw. 2006, 19, 1422–1429. [Google Scholar] [CrossRef] [PubMed]
- Sturm, W.; Willmes, K. On the functional neuroanatomy of intrinsic and phasic alertness. NeuroImage 2001, 14 (Pt. 2), S76–S84. [Google Scholar] [CrossRef]
- Thiebaut de Schotten, M.; Dell’Acqua, F.; Forkel, S.J.; Simmons, A.; Vergani, F.; Murphy, D.G.; Catani, M. A lateralized brain network for visuospatial attention. Nat. Neurosci. 2011, 14, 1245–1246. [Google Scholar] [CrossRef]
- Middag-van Spanje, M.; Duecker, F.; Gallotto, S.; de Graaf, T.A.; van Heugten, C.; Sack, A.T.; Schuhmann, T. Transcranial magnetic stimulation over posterior parietal cortex modulates alerting and executive control processes in attention. Eur. J. Neurosci. 2022, 56, 5853–5868. [Google Scholar] [CrossRef]
- Bunzeck, N.; Schutze, H.; Duzel, E. Category-specific organization of prefrontal response-facilitation during priming. Neuropsychologia 2006, 44, 1765–1776. [Google Scholar] [CrossRef]
- Burgess, P.W.; Dumontheil, I.; Gilbert, S.J. The gateway hypothesis of rostral prefrontal cortex (area 10) function. Trends Cogn. Sci. 2007, 11, 290–298. [Google Scholar] [CrossRef]
- Gomez, C.M.; Flores, A.; Ledesma, A. Fronto-parietal networks activation during the contingent negative variation period. Brain Res. Bull. 2007, 73, 40–47. [Google Scholar] [CrossRef]
- Praamstra, P.; Boutsen, L.; Humphreys, G.W. Frontoparietal control of spatial attention and motor intention in human EEG. J. Neurophysiol. 2005, 94, 764–774. [Google Scholar] [CrossRef]
- Kanwisher, N.; Wojciulik, E. Visual attention: Insights from brain imaging. Nat. Rev. Neurosci. 2000, 1, 91–100. [Google Scholar] [CrossRef]
- Chennu, S.; Noreika, V.; Gueorguiev, D.; Blenkmann, A.; Kochen, S.; Ibanez, A.; Owen, A.M.; Bekinschtein, T.A. Expectation and attention in hierarchical auditory prediction. J. Neurosci. 2013, 33, 11194–11205. [Google Scholar] [CrossRef] [Green Version]
- Chen, A.; Zhang, Z.; Cao, C.; Lu, J.; Wu, S.; Ma, S.; Feng, Y.; Wang, S.; Xu, G.; Song, J. Altered Attention Network in Paratroopers Exposed to Repetitive Subconcussion: Evidence Based on Behavioral and Event-Related Potential Results. J. Neurotrauma. 2021, 38, 3306–3314. [Google Scholar] [CrossRef] [PubMed]
- Fiebelkorn, I.C.; Kastner, S. Functional Specialization in the Attention Network. Annu. Rev. Psychol. 2020, 71, 221–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, G.; Rushby, J.A.; De Blasio, F.M. Neurophysiological correlates of visuospatial attention and the social dynamics of gaze processing. Cogn. Affect. Behav. Neurosci. 2019, 19, 1218–1230. [Google Scholar] [CrossRef] [PubMed]
- Corbetta, M.; Shulman, G.L. Control of goal-directed and stimulus-driven attention in the brain. Nat. Rev. Neurosci. 2002, 3, 201–215. [Google Scholar] [CrossRef]
- Posner, M.I. Cognitive Neuroscience of Attention; The Guilford Press: New York, NY, USA, 2012. [Google Scholar]
- Fiebelkorn, I.C.; Pinsk, M.A.; Kastner, S. A Dynamic Interplay within the Frontoparietal Network Underlies Rhythmic Spatial Attention. Neuron 2018, 99, 842–853 e8. [Google Scholar] [CrossRef] [Green Version]
- Dosenbach, N.U.; Fair, D.A.; Cohen, A.L.; Schlaggar, B.L.; Petersen, S.E. A dual-networks architecture of top-down control. Trends Cogn. Sci. 2008, 12, 99–105. [Google Scholar] [CrossRef] [Green Version]
- Poch, C.; Carretie, L.; Campo, P. A dual mechanism underlying alpha lateralization in attentional orienting to mental representation. Biol. Psychol. 2017, 128, 63–70. [Google Scholar] [CrossRef]
- Foxe, J.J.; Snyder, A.C. The Role of Alpha-Band Brain Oscillations as a Sensory Suppression Mechanism during Selective Attention. Front. Psychol. 2011, 2, 154. [Google Scholar] [CrossRef] [Green Version]
- Marshall, T.R.; O’Shea, J.; Jensen, O.; Bergmann, T.O. Frontal eye fields control attentional modulation of alpha and gamma oscillations in contralateral occipitoparietal cortex. J. Neurosci. 2015, 35, 1638–1647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thut, G.; Nietzel, A.; Brandt, S.A.; Pascual-Leone, A. Alpha-band electroencephalographic activity over occipital cortex indexes visuospatial attention bias and predicts visual target detection. J. Neurosci. 2006, 26, 9494–9502. [Google Scholar] [CrossRef] [Green Version]
- Posner, M.I.; Rothbart, M.K.; Voelker, P. Developing brain networks of attention. Curr. Opin. Pediatr. 2016, 28, 720–724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Senkowski, D.; Talsma, D.; Herrmann, C.S.; Woldorff, M.G. Multisensory processing and oscillatory gamma responses: Effects of spatial selective attention. Exp. Brain Res. 2005, 166, 411–426. [Google Scholar] [CrossRef]
- Koch, C.; Massimini, M.; Boly, M.; Tononi, G. Neural correlates of consciousness: Progress and problems. Nat. Rev. Neurosci. 2016, 17, 307–321. [Google Scholar] [CrossRef] [PubMed]
- Leicht, G.; Kirsch, V.; Giegling, I.; Karch, S.; Hantschk, I.; Moller, H.J.; Pogarell, O.; Hegerl, U.; Rujescu, D.; Mulert, C. Reduced early auditory evoked gamma-band response in patients with schizophrenia. Biol. Psychiatry 2010, 67, 224–231. [Google Scholar] [CrossRef]
- Tononi, G. An information integration theory of consciousness. BMC Neurosci. 2004, 5, 42. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Z.; Chen, H.; Ding, Z.; Li, Z.; Song, Y.; Li, X. The Modulating Effect of Top-down Attention on the Optimal Pre-target Onset Oscillatory States of Bottom-up Attention. Neuroscience 2021, 466, 186–195. [Google Scholar] [CrossRef]
- Xuan, B.; Mackie, M.A.; Spagna, A.; Wu, T.; Tian, Y.; Hof, P.R.; Fan, J. The activation of interactive attentional networks. NeuroImage 2016, 129, 308–319. [Google Scholar] [CrossRef] [Green Version]
- Matsui, T.; Hattori, Y.; Tsumura, K.; Aoki, R.; Takeda, M.; Nakahara, K.; Jimura, K. Executive control by fronto-parietal activity explains counterintuitive decision behavior in complex value-based decision-making. NeuroImage 2022, 249, 118892. [Google Scholar] [CrossRef]
- Billeke, P.; Ossandon, T.; Perrone-Bertolotti, M.; Kahane, P.; Bastin, J.; Jerbi, K.; Lachaux, J.P.; Fuentealba, P. Human Anterior Insula Encodes Performance Feedback and Relays Prediction Error to the Medial Prefrontal Cortex. Cereb. Cortex 2020, 30, 4011–4025. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Liu, X.; Guise, K.G.; Knight, R.T.; Ghajar, J.; Fan, J. Effective connectivity of the fronto-parietal network during attentional control. J. Cogn. Neurosci. 2010, 22, 543–553. [Google Scholar] [CrossRef]
- Nee, D.E.; Wager, T.D.; Jonides, J. Interference resolution: Insights from a meta-analysis of neuroimaging tasks. Cogn. Affect. Behav. Neurosci. 2007, 7, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, M.X. Error-related medial frontal theta activity predicts cingulate-related structural connectivity. NeuroImage 2011, 55, 1373–1383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, M.X.; Ridderinkhof, K.R. EEG source reconstruction reveals frontal-parietal dynamics of spatial conflict processing. PLoS ONE 2013, 8, e57293. [Google Scholar] [CrossRef]
- Cohen, M.X.; Donner, T.H. Midfrontal conflict-related theta-band power reflects neural oscillations that predict behavior. J. Neurophysiol. 2013, 110, 2752–2763. [Google Scholar] [CrossRef] [Green Version]
- Compton, R.J.; Arnstein, D.; Freedman, G.; Dainer-Best, J.; Liss, A. Cognitive control in the intertrial interval: Evidence from EEG alpha power. Psychophysiology 2011, 48, 583–590. [Google Scholar] [CrossRef]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duan, K.; Xie, S.; Zhang, X.; Xie, X.; Cui, Y.; Liu, R.; Xu, J. Exploring the Temporal Patterns of Dynamic Information Flow during Attention Network Test (ANT). Brain Sci. 2023, 13, 247. https://doi.org/10.3390/brainsci13020247
Duan K, Xie S, Zhang X, Xie X, Cui Y, Liu R, Xu J. Exploring the Temporal Patterns of Dynamic Information Flow during Attention Network Test (ANT). Brain Sciences. 2023; 13(2):247. https://doi.org/10.3390/brainsci13020247
Chicago/Turabian StyleDuan, Keyi, Songyun Xie, Xin Zhang, Xinzhou Xie, Yujie Cui, Ruizhen Liu, and Jian Xu. 2023. "Exploring the Temporal Patterns of Dynamic Information Flow during Attention Network Test (ANT)" Brain Sciences 13, no. 2: 247. https://doi.org/10.3390/brainsci13020247