
Decrease in the P2 Amplitude of Object Working Memory after 8 h-Recovery Sleep Following 36 h-Total Sleep Deprivation: An ERP Study


Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Experimental Task
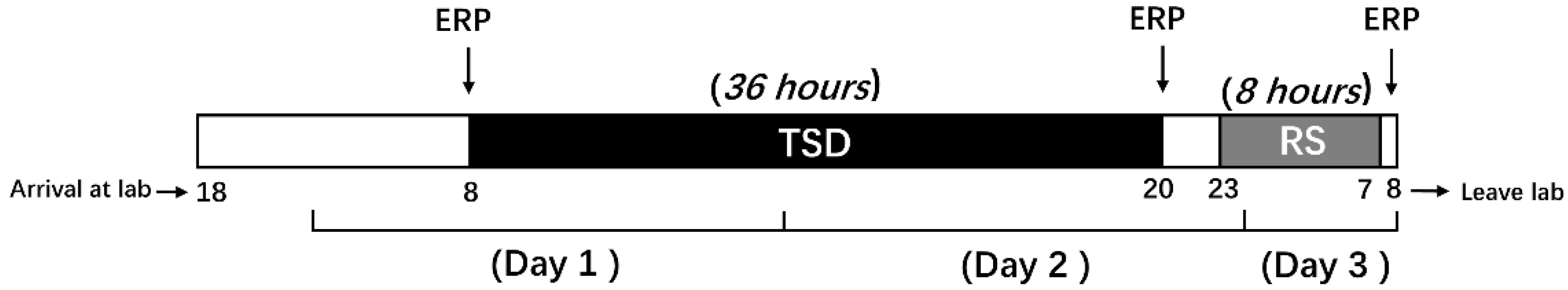
2.3. Experimental Procedures
2.4. EEG Data Collection and Recordings
2.5. Data Analysis
3. Results
3.1. Behavior Performance
3.2. Amplitude
3.3. Latency
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Periasamy, S.; Hsu, D.-Z.; Fu, Y.-H.; Liu, M.-Y. Sleep deprivation-induced multi-organ injury: Role of oxidative stress and inflammation. Excli J. 2015, 14, 672–683. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, S.; Liu, C.-Y.; Hayashi, Y. Sleep in vertebrate and invertebrate animals, and insights into the function and evolution of sleep. Neurosci. Res. 2017, 118, 3–12. [Google Scholar] [CrossRef]
- Kotronoulas, G.; Stamatakis, A.; Stylianopoulou, F. Hormones, hormonal agents, and neuropeptides involved in the neuroendocrine regulation of sleep in humans. Hormones 2009, 8, 232–248. [Google Scholar] [CrossRef] [PubMed]
- AlDabal, L. Metabolic, endocrine, and immune consequences of sleep deprivation. Open Respir. Med. J. 2011, 5, 31–43. [Google Scholar] [CrossRef] [PubMed]
- Killgore, W.D.S. Effects of sleep deprivation on cognition. Prog. Brain Res. 2010, 185, 105–129. [Google Scholar] [CrossRef]
- Pan, J. A White Paper on “Chinese People Sleeping during COVID-19” Was Released; Chinese Sleep Research Society: Harbin, China, 2020; Volume 004. [Google Scholar]
- Baddeley, A.D.; Hitch, G. Working memory. Psychol. Learn. Motiv. 1974, 8, 47–89. [Google Scholar] [CrossRef]
- Choo, W.-C.; Lee, W.-W.; Venkatraman, V.; Sheu, F.-S.; Chee, M.W. Dissociation of cortical regions modulated by both working memory load and sleep deprivation and by sleep deprivation alone. NeuroImage 2005, 25, 579–587. [Google Scholar] [CrossRef]
- Diamond, A. Executive functions. Annu. Rev. Psychol. 2013, 64, 135–168. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Shao, Y.; Liu, Z.; Li, C.; Chen, Y.; Zhou, Q. Decreased Information Replacement of Working Memory After Sleep Deprivation: Evidence from an Event-Related Potential Study. Front. Neurosci. 2019, 13, 408. [Google Scholar] [CrossRef]
- Peng, Z.; Dai, C.; Cai, X.; Zeng, L.; Li, J.; Xie, S.; Wang, H.; Yang, T.; Shao, Y.; Wang, Y. Total Sleep Deprivation Impairs Lateralization of Spatial Working Memory in Young Men. Front. Neurosci. 2020, 14, 562035. [Google Scholar] [CrossRef]
- Chee, M.W.L.; Choo, W.C. Functional imaging of working memory after 24 hr of total sleep deprivation. J. Neurosci. 2004, 24, 4560–4567. [Google Scholar] [CrossRef] [PubMed]
- Habeck, C.; Rakitin, B.C.; Moeller, J.; Scarmeas, N.; Zarahn, E.; Brown, T.; Stern, Y. An event-related fMRI study of the neurobehavioral impact of sleep deprivation on performance of a delayed-match-to-sample task. Cogn. Brain Res. 2004, 18, 306–321. [Google Scholar] [CrossRef] [PubMed]
- Almklov, E.L.; Drummond, S.P.A.; Orff, H.; Alhassoon, O.M. The effects of sleep deprivation on brain functioning in older adults. Behav. Sleep Med. 2015, 13, 324–345. [Google Scholar] [CrossRef] [PubMed]
- Cabeza, R.; Dolcos, F.; Graham, R.; Nyberg, L. Similarities and differences in the neural correlates of episodic memory retrieval and working memory. NeuroImage 2002, 16, 317–330. [Google Scholar] [CrossRef]
- Ullsperger, M.; von Cramon, D. Subprocesses of performance monitoring: A dissociation of error processing and response competition revealed by event-related fMRI and ERPs. NeuroImage 2001, 14, 1387–1401. [Google Scholar] [CrossRef]
- Lythe, K.E.; Williams, S.C.; Anderson, C.; Libri, V.; Mehta, M.A. Frontal and parietal activity after sleep deprivation is dependent on task difficulty and can be predicted by the fMRI response after normal sleep. Behav. Brain Res. 2012, 233, 62–70. [Google Scholar] [CrossRef]
- Chengyang, L.; Daqing, H.; Jianlin, Q.; Haisheng, C.; Qingqing, M.; Jin, W.; Jiajia, L.; Enmao, Y.; Yongcong, S.; Xi, Z. Short-term memory deficits correlate with hippocampal-thalamic functional connectivity alterations following acute sleep restriction. Brain Imaging Behav. 2017, 11, 954–963. [Google Scholar] [CrossRef]
- Lei, Y.; Shao, Y.; Wang, L.; Zhai, T.; Zou, F.; Ye, E.; Jin, X.; Li, W.; Qi, J.; Yang, Z. Large-Scale Brain Network Coupling Predicts Total Sleep Deprivation Effects on Cognitive Capacity. PLoS ONE 2015, 10, e0133959. [Google Scholar] [CrossRef] [PubMed]
- Picton, T.W.; Lins, O.G.; Scherg, M. The recording and analysis of event-related potentials. In Handbook of Neuropsychology; Elseiver: Amsterdam, The Netherlands, 1995; Volume 10, pp. 3–73. [Google Scholar]
- Mecklinger, A.; Pfeifer, E. Event-related potentials reveal topographical and temporal distinct neuronal activation patterns for spatial and object working memory. Cogn. Brain Res. 1996, 4, 211–224. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-J.; Kim, L.; Kim, Y.-K.; Suh, K.-Y.; Han, J.; Park, M.-K.; Park, K.-W.; Lee, D.-H. Auditory event-related potentials and psychological changes during sleep deprivation. Neuropsychobiology 2004, 50, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Brauer, K.A. Sleep Deprivation and Recovery: The Effects of P300 Three and Six Hours Post Recovery. Ph.D. Thesis, Missouri State University, Springfield, MI, USA, 2016. Volume 2953. [Google Scholar]
- Woldorff, M.G.; Hackley, S.A.; Hillyard, S.A. The effects of channel-selective attention on the mismatch negativity wave elicited by deviant tones. Psychophysiology 1991, 28, 30–42. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.C.; Gillin, J.C.; Buchsbaum, M.S.; Chen, P.; Keator, D.B.; Wu, N.K.; Darnall, L.A.; Fallon, J.H.; Bunney, W.E. Frontal lobe metabolic decreases with sleep deprivation not totally reversed by recovery sleep. Neuropsychopharmacology 2006, 31, 2783–2792. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.-C.; Whitehurst, L.N.; Naji, M.; Mednick, S.C. Autonomic Activity during a Daytime Nap Facilitates Working Memory Improvement. J. Cogn. Neurosci. 2020, 32, 1963–1974. [Google Scholar] [CrossRef] [PubMed]
- Buysse, D.J.; Reynolds, C.F., III; Monk, T.H.; Berman, S.R.; Kupfer, D.J. The Pittsburgh sleep quality index: A new instrument for psychiatric practice and research. Psychiatry Res. 1989, 28, 193–213. [Google Scholar] [CrossRef] [PubMed]
- Vogel, E.K.; Luck, S.J. The visual N1 component as an index of a discrimination process. Psychophysiology 2000, 37, 190–203. [Google Scholar] [CrossRef] [PubMed]
- Schutter, D.J.; de Haan, E.H.; van Honk, J. Functionally dissociated aspects in anterior and posterior electrocortical processing of facial threat. Int. J. Psychophysiol. 2004, 53, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Luck, S.J.; Hillyard, S.A. Electrophysiological correlates of feature analysis during visual search. Psychophysiology 1994, 31, 291–308. [Google Scholar] [CrossRef]
- Smith, M.E.; McEvoy, L.K.; Gevins, A. The impact of moderate sleep loss on neurophysiologic signals during working-memory task performance. Sleep 2002, 25, 56–66. [Google Scholar] [CrossRef]
- Drummond, S.P.; Brown, G.G.; Salamat, J.S.; Gillin, J.C. Increasing task difficulty facilitates the cerebral compensatory response to total sleep deprivation. Sleep 2004, 27, 445–451. [Google Scholar] [CrossRef]
- Campbell, K.B.; Colrain, I.M. Event-related potential measures of the inhibition of information processing: II. The sleep onset period. Int. J. Psychophysiol. 2002, 46, 197–214. [Google Scholar] [CrossRef]
- Baddeley, A. Working memory. Curr. Biol. 2010, 20, R136–R140. [Google Scholar] [CrossRef]
- Kusztor, A.; Raud, L.; Juel, B.E.; Nilsen, A.S.; Storm, J.F.; Huster, R.J. Sleep deprivation differentially affects subcomponents of cognitive control. Sleep 2019, 42, zsz016. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Ye, E.; Qi, J.; Wang, L.; Lei, Y.; Chen, P.; Mi, G.; Zou, F.; Shao, Y.; Yang, Z. Recovery Sleep Reverses Impaired Response Inhibition due to Sleep Restriction: Evidence from a Visual Event Related Potentials Study. PLoS ONE 2015, 10, e0142361. [Google Scholar] [CrossRef] [PubMed]
- Suwazono, S.; Machado, L.; Knight, R.T. Predictive value of novel stimuli modifies visual event-related potentials and behavior. Clin. Neurophysiol. 2000, 111, 29–39. [Google Scholar] [CrossRef]
- Johnson, R.; Donchin, E. P300 and stimulus categorization: Two plus one is not so different from one plus one. Psychophysiology 1980, 17, 167–178. [Google Scholar] [CrossRef] [PubMed]
- Koslowsky, M.; Babkoff, H. Meta-analysis of the relationship between total sleep deprivation and performance. Chrono-Int. 1992, 9, 132–136. [Google Scholar] [CrossRef]
- Verleger, R. On the utility of P3 latency as an index of mental chronometry. Psychophysiology 1997, 34, 131–156. [Google Scholar] [CrossRef] [PubMed]
- Kerkhof, G.; Korving, H.; Geest, H.-V.; Rietveld, W. Diurnal differences between morning-type and evening-type subjects in self-rated alertness, body temperature and the visual and auditory evoked potential. Neurosci. Lett. 1980, 16, 11–15. [Google Scholar] [CrossRef] [PubMed]
- Montplaisir, J. Depression and biological rhythm: Therapeutic effects of sleep deprivation. L’union medicale du Can. 1981, 110, 272–276. [Google Scholar]
- Lavie, P. Sleep-wake as a biological rhythm. Annu. Rev. Psychol. 2001, 52, 277–303. [Google Scholar] [CrossRef] [PubMed]
- Turner, T.H.; Drummond, S.P.A.; Salamat, J.S.; Brown, G.G. Effects of 42 hr of total sleep deprivation on component processes of verbal working memory. Neuropsychology 2007, 21, 787–795. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Nocturnal Sleep | Sleep Deprivation |
|---|---|---|
| Number | 20 | 20 |
| Gender | Male | Male |
| Age (y) | 23.55 ± 2.33 | 23.11 ± 2.40 |
| PSQI | 3.54 ± 0.93 | 3.66 ± 0.50 |
| NS-BS | NS-SD0 | |
|---|---|---|
| Mean Reaction time | 519.83 ± 117.88 | 538.57 ± 134.62 |
| Correct rate | 0.89 ± 0.07 | 0.89 ± 0.06 |
| Correct number/s | 1.83 ± 0.47 | 1.79 ± 0.49 |
| BS | 36 h-TSD | 8 h-RS | |
|---|---|---|---|
| Mean Reaction time | 522.19 ± 88.44 | 543.85 ± 107.88 | 526.07 ± 80.19 |
| Correct rate | 0.88 ± 0.09 | 0.79 ± 0.13 * | 0.87 ± 0.07 |
| Correct number/s | 1.76 ± 0.44 | 1.53 ± 0.45 * | 1.69 ± 0.34 |
| Amplitude | Latency | |||
|---|---|---|---|---|
| N1 | P2 | N1 | P2 | |
| NS-BS | −2.05 ± 2.82 | 8.07 ± 2.15 | 98.13 ± 18.98 | 183.63 ± 20.95 |
| NS-SD0 | −1.95 ± 2.23 | 7.40 ± 2.90 | 94.66 ± 17.12 | 179.87 ± 26.47 |
| Amplitude | Latency | |||
|---|---|---|---|---|
| N1 | P2 | N1 | P2 | |
| BS | −2.89 ± 1.62 | 8.63 ± 4.99 | 92.88 ± 20.43 | 182.28 ± 23.26 |
| 36 h-TSD | −4.40 ± 3.63 | 11.91 ± 6.50 | 93.41 ± 12.01 | 160.88 ± 15.72 * |
| 8 h-RS | −3.25 ± 2.69 | 7.15 ± 4.90 | 97.12 ± 20.46 | 165.20 ± 14.29 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shao, Y.; Peng, Z.; Xu, L.; Lian, J.; An, X.; Cheng, M.-Y. Decrease in the P2 Amplitude of Object Working Memory after 8 h-Recovery Sleep Following 36 h-Total Sleep Deprivation: An ERP Study. Brain Sci. 2023, 13, 1470. https://doi.org/10.3390/brainsci13101470
Shao Y, Peng Z, Xu L, Lian J, An X, Cheng M-Y. Decrease in the P2 Amplitude of Object Working Memory after 8 h-Recovery Sleep Following 36 h-Total Sleep Deprivation: An ERP Study. Brain Sciences. 2023; 13(10):1470. https://doi.org/10.3390/brainsci13101470
Chicago/Turabian StyleShao, Yongcong, Ziyi Peng, Lin Xu, Jie Lian, Xin An, and Ming-Yang Cheng. 2023. "Decrease in the P2 Amplitude of Object Working Memory after 8 h-Recovery Sleep Following 36 h-Total Sleep Deprivation: An ERP Study" Brain Sciences 13, no. 10: 1470. https://doi.org/10.3390/brainsci13101470