

Brief Maternal Separation Promotes Resilience to Anxiety-like and Depressive-like Behaviors in Female C57BL/6J Offspring with Imiquimod-Induced Psoriasis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods and Animals
2.1. Animals
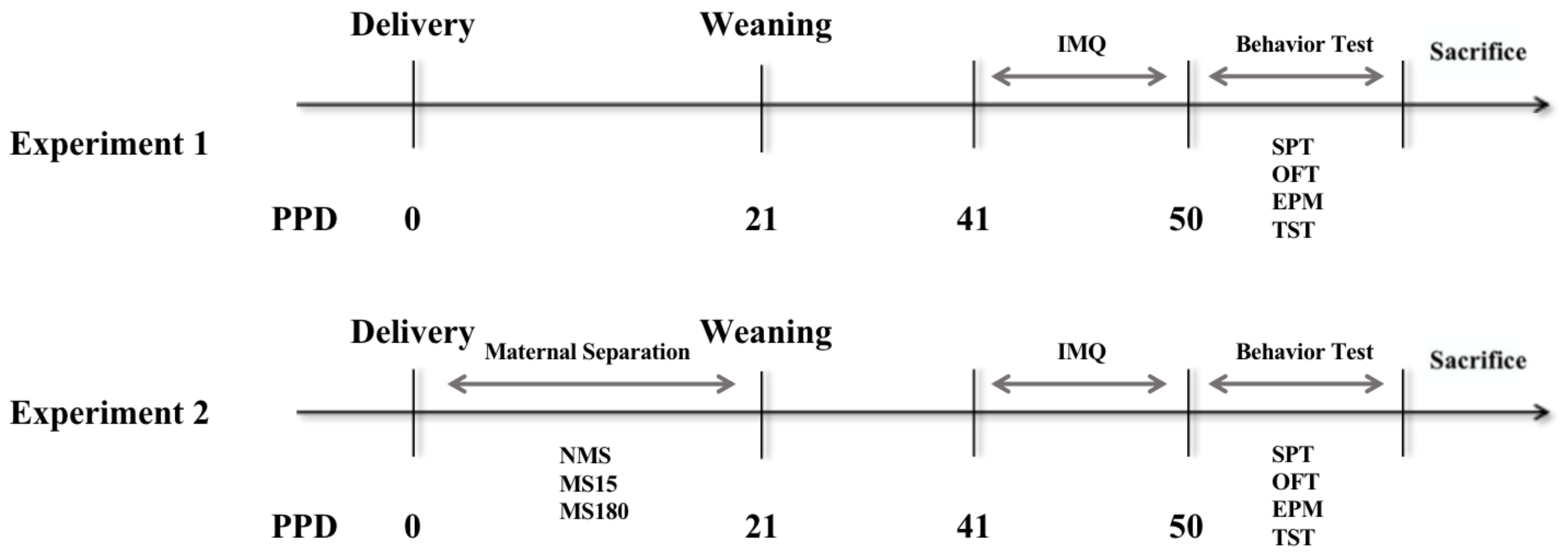
2.2. Experimental Design
2.3. Maternal Separation (MS)
2.4. Sucrose Preference Test (SPT)
2.5. Open Field Test (OFT)
2.6. Elevated Plus Maze (EPM) Test
2.7. Tail Suspension Test (TST)
2.8. Psoriasis Area and Severity Index Score
2.9. Tissue Collection
2.10. Western Blot Analysis
2.11. Immunofluorescence
2.12. Histopathological Examination and Immunohistochemical Staining
2.13. Statistical Analysis
3. Results
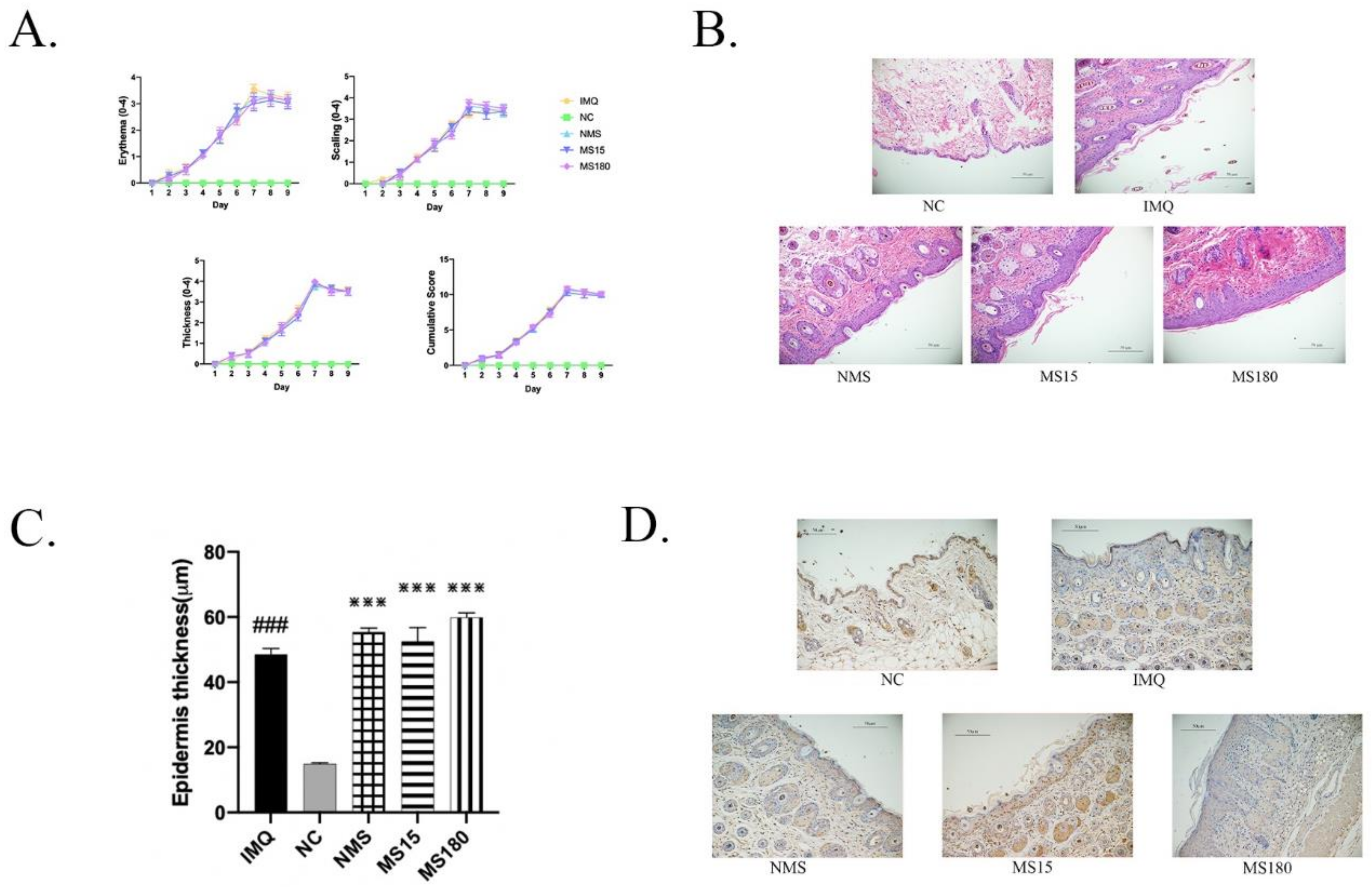
3.1. The Skin Lesions of Psoriasis-like Female Mice
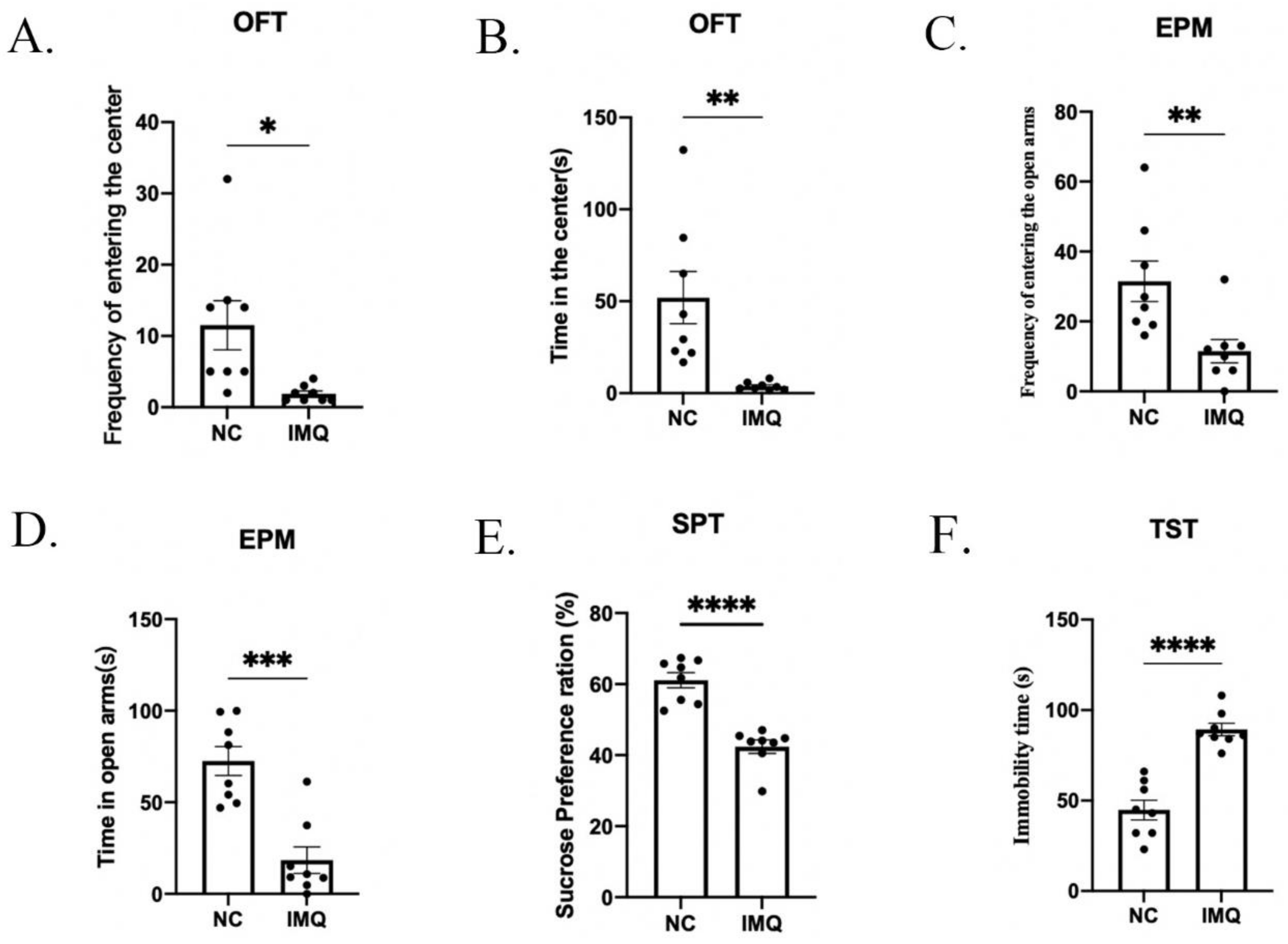
3.2. Anxiety and Depressive-like Behaviors in Psoriasis-like Female Mice
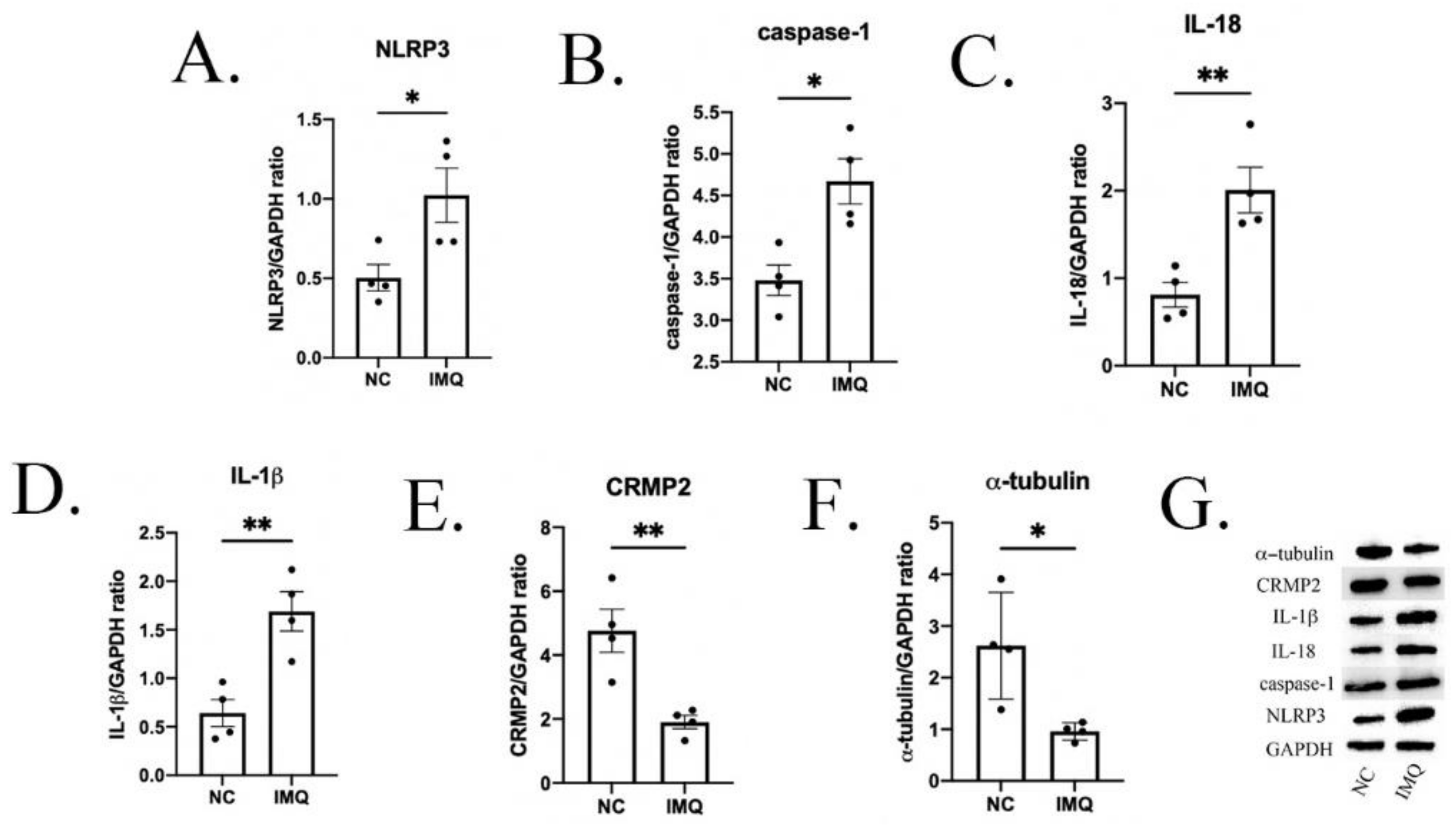
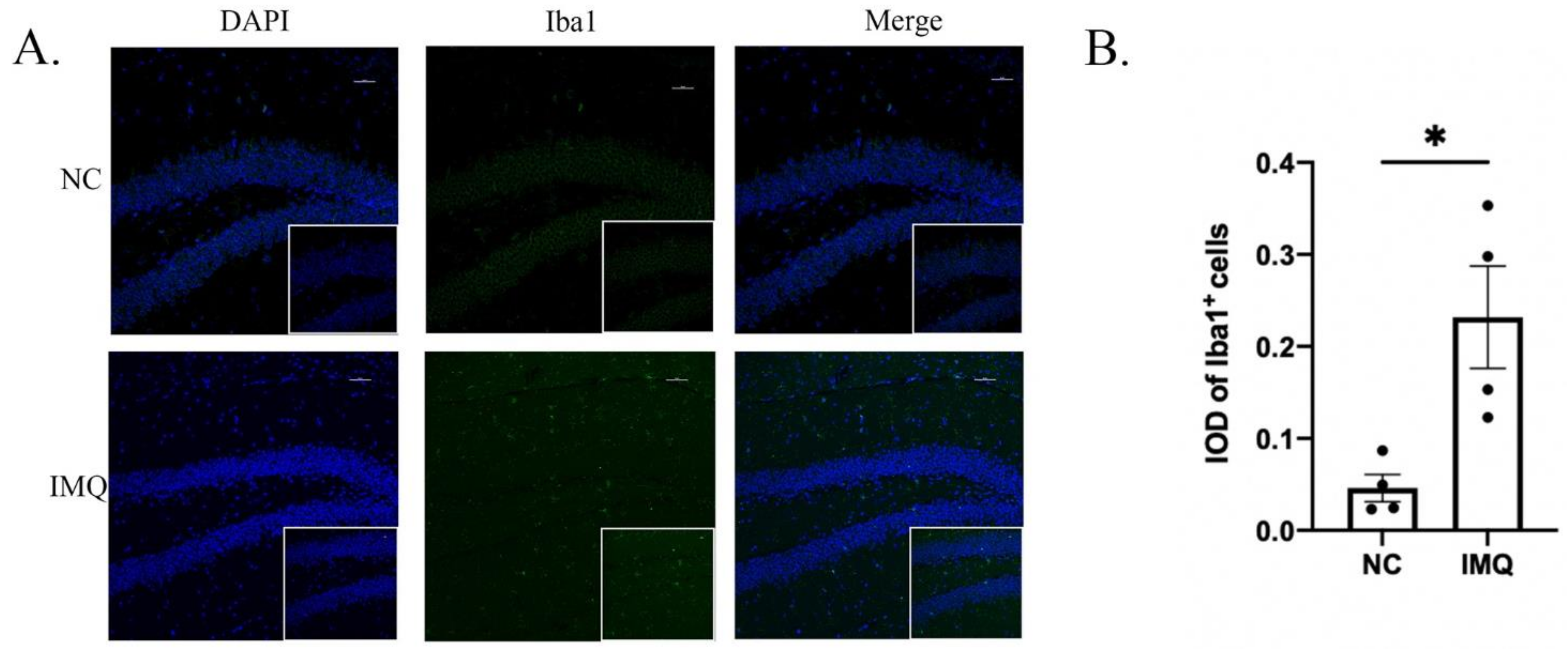
3.3. Neuroinflammation Changes in Psoriasis-like Female Mice
3.4. Neuroplasticity Changes in Psoriasis-like Female Mice
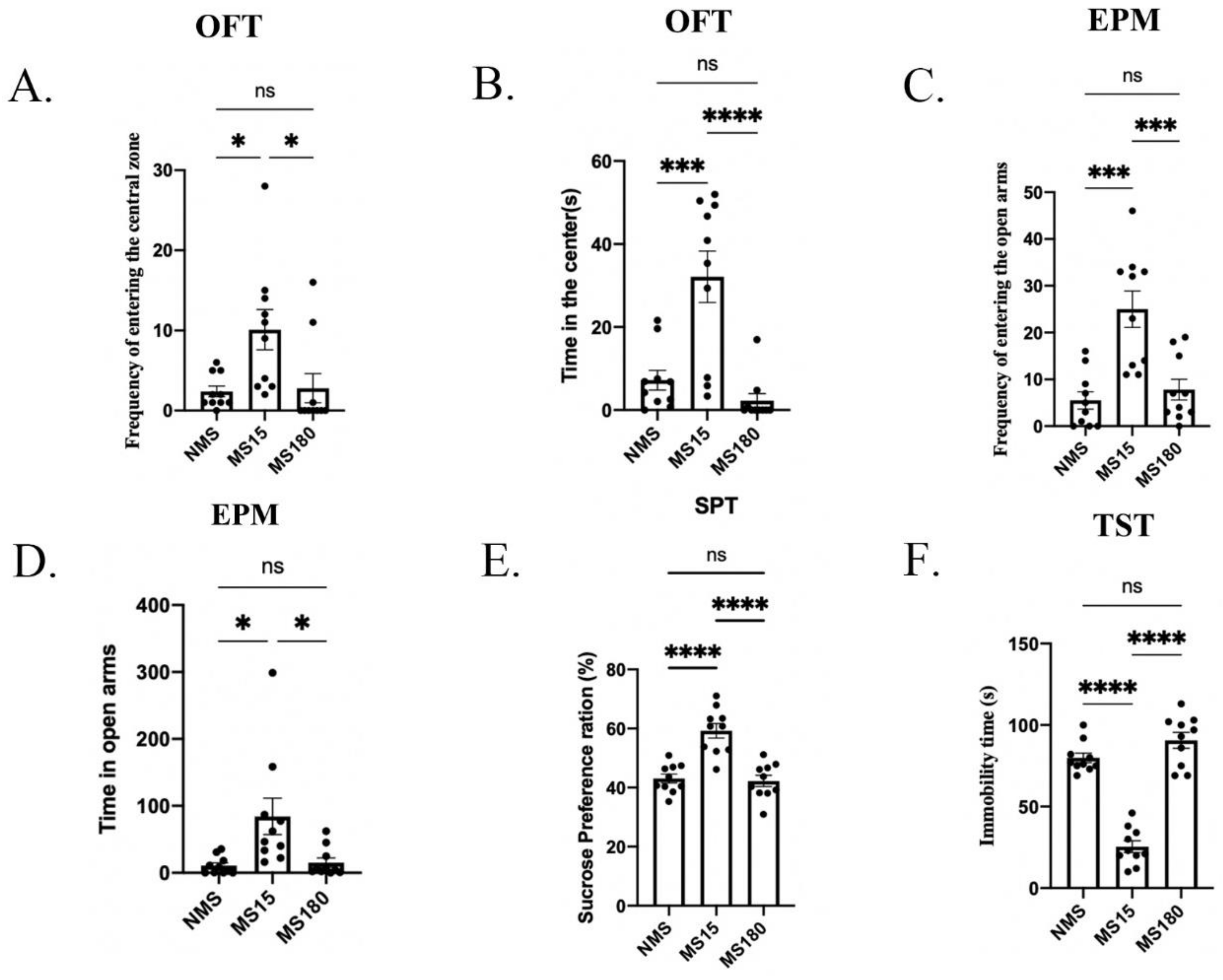
3.5. Brief MS Promoted Resilience to Behavior Deficits in Psoriasis-like Female Offspring
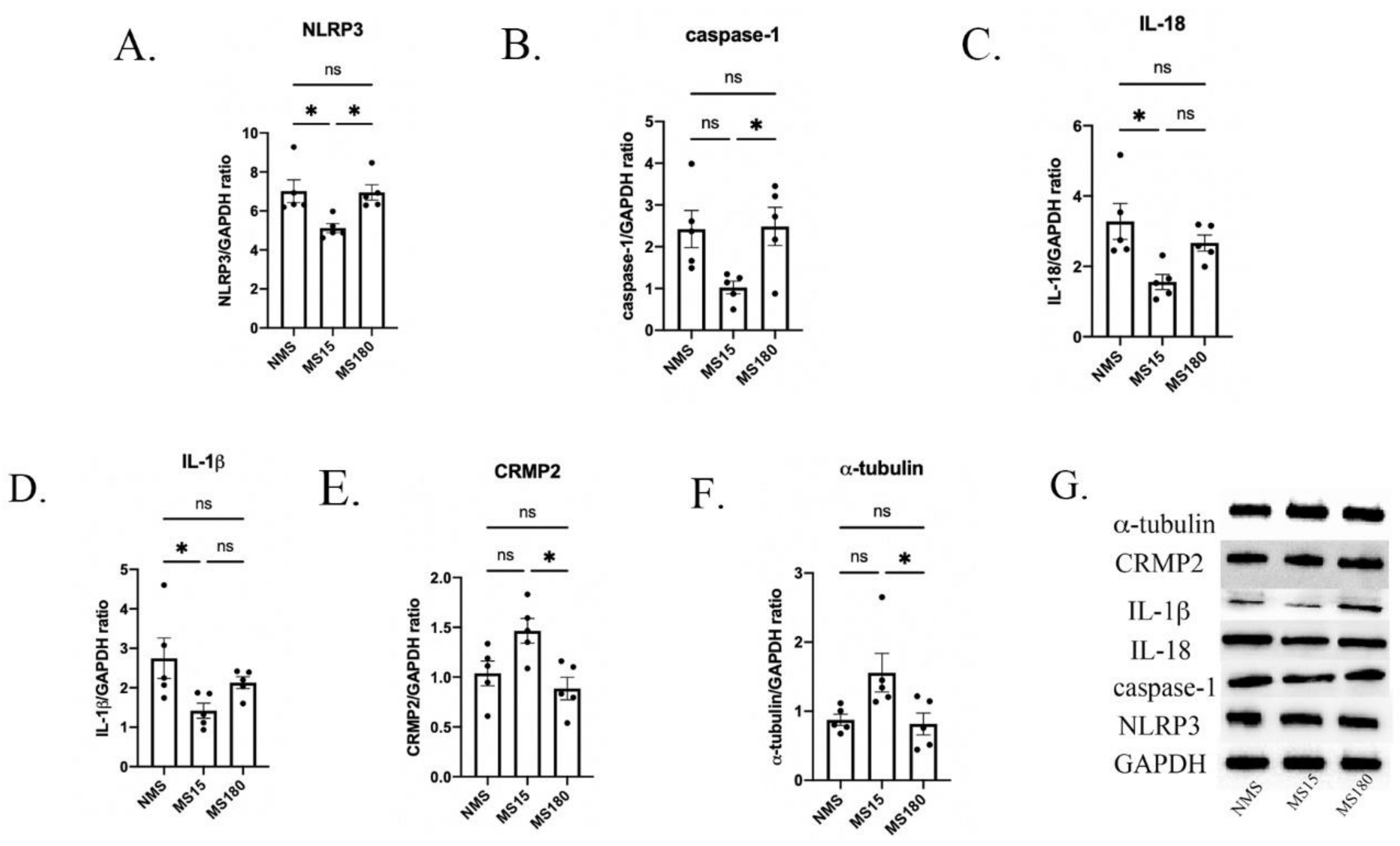
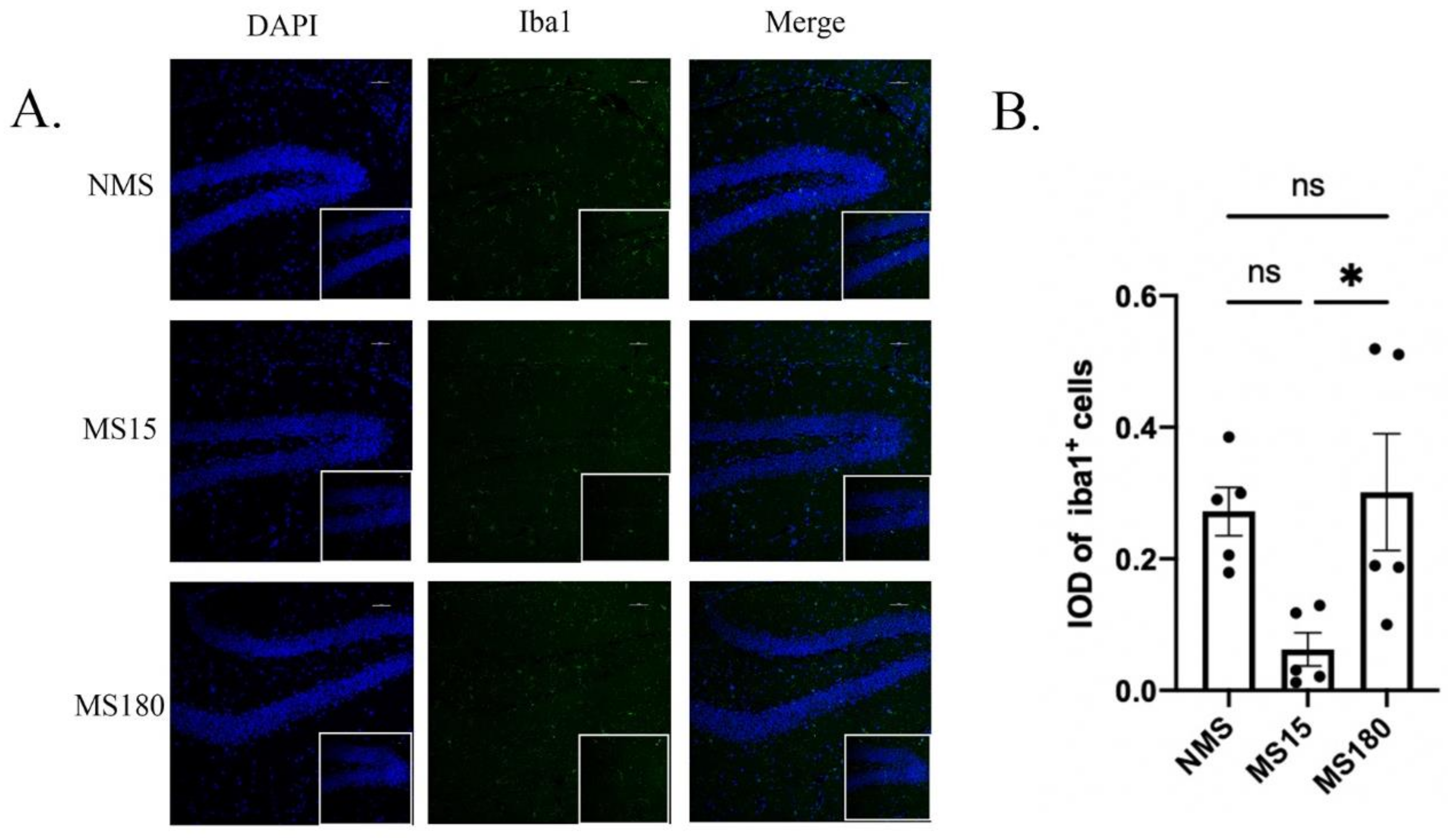
3.6. MS15 Protocol Inhibited Activation of Neuroinflammation in the Hippocampus of Female Mice Resilience to Behavior Deficits
3.7. MS15 Protocol Inhibited Hippocampal Neuroplastic Injury in Female Mice Resilience to Behavior Deficits
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bell, K.A.; Balogh, E.A.; Feldman, S.R. An update on the impact of depression on the treatment of psoriasis. Expert Opin. Pharm. 2021, 22, 695–703. [Google Scholar] [CrossRef] [PubMed]
- Stewart, T.J.; Tong, W.; Whitfeld, M.J. The associations between psychological stress and psoriasis: A systematic review. Int. J. Derm. 2018, 57, 1275–1282. [Google Scholar] [CrossRef] [PubMed]
- Rabeony, H.; Pohin, M.; Vasseur, P.; Petit-Paris, I.; Jégou, J.F.; Favot, L.; Frouin, E.; Boutet, M.A.; Blanchard, F.; Togbe, D.; et al. IMQ-induced skin inflammation in mice is dependent on IL-1R1 and MyD88 signaling but independent of the NLRP3 inflammasome. Eur. J. Immunol. 2015, 45, 2847–2857. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Liu, Y.; Guo, X.; Meng, Y.; Qi, C.; Zhao, J.; Di, T.; Zhang, L.; Guo, X.; Wang, Y.; et al. Depressive-like behaviors in mice with Imiquimod-induced psoriasis. Int. Immunopharmacol. 2020, 89 Pt B, 107057. [Google Scholar] [CrossRef]
- Troubat, R.; Barone, P.; Leman, S.; Desmidt, T.; Cressant, A.; Atanasova, B.; Brizard, B.; El Hage, W.; Surget, A.; Belzung, C.; et al. Neuroinflammation and depression: A review. Eur. J. Neurosci. 2021, 53, 151–171. [Google Scholar] [CrossRef]
- Liu, W.; Ge, T.; Leng, Y.; Pan, Z.; Fan, J.; Yang, W.; Cui, R. The Role of Neural Plasticity in Depression: From Hippocampus to Prefrontal Cortex. Neural. Plast. 2017, 6871089. [Google Scholar] [CrossRef]
- Di Filippo, M.; Sarchielli, P.; Picconi, B.; Calabresi, P. Neuroinflammation and synaptic plasticity: Theoretical basis for a novel, immune-centred, therapeutic approach to neurological disorders. Trends Pharm. Sci. 2008, 29, 402–412. [Google Scholar] [CrossRef]
- Zheng, Z.H.; Tu, J.L.; Li, X.H.; Hua, Q.; Liu, W.Z.; Liu, Y.; Pan, B.X.; Hu, P.; Zhang, W.H. Neuroinflammation induces anxiety- and depressive-like behavior by modulating neuronal plasticity in the basolateral amygdala. Brain Behav. Immun. 2021, 91, 505–518. [Google Scholar] [CrossRef]
- Xiang, D.; Xiao, J.; Sun, S.; Fu, L.; Yao, L.; Wang, G.; Liu, Z. Differential Regulation of DNA Methylation at the CRMP2 Promoter Region. Between the Hippocampus and Prefrontal Cortex in a CUMS Depression Model. Front. Psychiatry 2020, 11, 141. [Google Scholar] [CrossRef]
- Li, W.; Liu, X.; Qiao, H. Downregulation of hippocampal SIRT6 activates AKT/CRMP2 signaling and ameliorates chronic stress-induced depression-like behavior in mice. Acta Pharm. Sin. 2020, 41, 1557–1567. [Google Scholar] [CrossRef]
- Wei, Y.; Wang, G.; Chen, J.; Xiao, L.; Wu, Z.; He, J.; Zhang, N. Maternal deprivation induces cytoskeletal alterations and depressive-like behavior in adult male rats by regulating the AKT/GSK3β/CRMP2 signaling pathway. Physiol. Behav. 2021, 242, 113625. [Google Scholar] [CrossRef]
- Robakis, T.K.; Watson-Lin, K.; Wroolie, T.E.; Myoraku, A.; Nasca, C.; Bigio, B.; McEwen, B.; Rasgon, N.L. Early life adversity blunts responses to pioglitazone in depressed, overweight adults. Eur. Psychiatry 2019, 55, 4–9. [Google Scholar] [CrossRef] [PubMed]
- Shen, S.; Chen, Z.; Qin, X.; Zhang, M.; Dai, Q. Remote and adjacent psychological predictors of early-adulthood resilience: Role of early-life trauma, extraversion, life-events, depression, and social-support. PLoS ONE 2021, 16, e0251859. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, J.M. Childhood mistreatment and adolescent and young adult depression. Soc. Sci. Med. 2009, 68, 799–806. [Google Scholar] [CrossRef]
- Peng, A.; Qiu, X.; Ji, S.; Hu, D.; Dong, B.; Song, T.; Huang, C.; Chen, L. The impact of childhood parental loss on risk for depression and anxiety in adulthood: A community-based study in Southwest China. J. Affect. Disord. 2022, 298 Pt A, 104–109. [Google Scholar] [CrossRef]
- McKay, M.T.; Cannon, M.; Chambers, D.; Conroy, R.M.; Coughlan, H.; Dodd, P.; Healy, C.; O’Donnell, L.; Clarke, M.C. Childhood trauma and adult mental disorder: A systematic review and meta-analysis of longitudinal cohort studies. Acta Psychiatry Scand. 2021, 143, 189–205. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Wu, Z.; Wang, G.; Xiao, L.; Wang, H.; Sun, L.; Xie, Y. Long-term maternal separation potentiates depressive-like behaviours and neuroinflammation in adult male C57/BL6J mice. Pharm. Biochem. Behav. 2020, 196, 172953. [Google Scholar] [CrossRef] [PubMed]
- Calpe-López, C.; Martínez-Caballero, M.A.; García-Pardo, M.P.; Aguilar, M.A. Brief Maternal Separation Inoculates Against the Effects of Social Stress on Depression-Like Behavior and Cocaine Reward in Mice. Front. Pharm. 2022, 13, 825522. [Google Scholar] [CrossRef]
- Vetulani, J. Early maternal separation: A rodent model of depression and a prevailing human condition. Pharm. Rep. 2013, 65, 1451–1461. [Google Scholar] [CrossRef]
- Plotsky, P.M.; Thrivikraman, K.V.; Nemeroff, C.B.; Caldji, C.; Sharma, S.; Meaney, M.J. Long-term consequences of neonatal rearing on central corticotropin-releasing factor systems in adult male rat offspring. Neuropsychopharmacology 2005, 30, 2192–2204. [Google Scholar] [CrossRef] [Green Version]
- Wei, Y.; Wang, G.; Wang, H.; He, J.; Zhang, N.; Wu, Z.; Xiao, L.; Yang, C. Sex-dependent impact of different degrees of maternal separation experience on OFT behavioral performances after adult chronic unpredictable mild stress exposure in rats. Physiol. Behav. 2018, 194, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Su, W.J.; Chen, Y.; Wu, T.Y.; Gong, H.; Shen, X.L.; Wang, Y.X.; Sun, X.J.; Jiang, C.L. Effects of hydrogen-rich water on depressive-like behavior in mice. Sci. Rep. 2016, 6, 23742. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, L.; Liu, Y.Z.; Shen, X.L.; Wu, T.Y.; Zhang, T.; Wang, W.; Wang, Y.X.; Jiang, C.L. NLRP3 Inflammasome Mediates Chronic Mild Stress-Induced Depression in Mice via Neuroinflammation. Int. J. Neuropsychopharmacol. 2015, 18, pyv006. [Google Scholar] [CrossRef] [PubMed]
- Kraeuter, A.K.; Guest, P.C.; Sarnyai, Z. The Open Field Test. for Measuring Locomotor Activity and Anxiety-Like Behavior. Methods Mol. Biol. 2019, 1916, 99–103. [Google Scholar]
- Kraeuter, A.K.; Guest, P.C.; Sarnyai, Z. The Elevated Plus Maze Test. for Measuring Anxiety-Like Behavior in Rodents. Methods Mol. Biol. 2019, 1916, 69–74. [Google Scholar]
- Cryan, J.F.; Mombereau, C.; Vassout, A. The tail suspension test as a model for assessing antidepressant activity: Review of pharmacological and genetic studies in mice. Neurosci. Biobehav. Rev. 2005, 29, 571–625. [Google Scholar] [CrossRef]
- Stein, D.J.; Vasconcelos, M.F.; Albrechet-Souza, L.; Ceresér, K.; de Almeida, R. Microglial Over-Activation by Social Defeat Stress Contributes to Anxiety-and Depressive-Like Behaviors. Front. Behav. Neurosci. 2017, 11, 207. [Google Scholar] [CrossRef]
- Xu, Y.; Sheng, H.; Bao, Q.; Wang, Y.; Lu, J.; Ni, X. NLRP3 inflammasome activation mediates estrogen deficiency-induced depression- and anxiety-like behavior and hippocampal inflammation in mice. Brain Behav. Immun. 2016, 56, 175–186. [Google Scholar] [CrossRef]
- Zhang, Y.; Huang, R.; Cheng, M.; Wang, L.; Chao, J.; Li, J.; Zheng, P.; Xie, P.; Zhang, Z.; Yao, H. Gut microbiota from NLRP3-deficient mice ameliorates depressive-like behaviors by regulating astrocyte dysfunction via circHIPK2. Microbiome 2019, 7, 116. [Google Scholar] [CrossRef]
- Arioz, B.I.; Tastan, B.; Tarakcioglu, E.; Tufekci, K.U.; Olcum, M.; Ersoy, N.; Bagriyanik, A.; Genc, K.; Genc, S. Melatonin Attenuates LPS-Induced Acute Depressive-Like Behaviors and Microglial NLRP3 Inflammasome Activation Through the SIRT1/Nrf2 Pathway. Front. Immunol. 2019, 10, 1511. [Google Scholar] [CrossRef]
- Xie, J.; Bi, B.; Qin, Y.; Dong, W.; Zhong, J.; Li, M.; Cheng, Y.; Xu, J.; Wang, H. Inhibition of phosphodiesterase-4 suppresses HMGB1/RAGE signaling pathway and NLRP3 inflammasome activation in mice exposed to chronic unpredictable mild stress. Brain Behav. Immun. 2021, 92, 67–77. [Google Scholar] [CrossRef] [PubMed]
- Bassett, B.; Subramaniyam, S.; Fan, Y.; Varney, S.; Pan, H.; Carneiro, A.; Chung, C.Y. Minocycline alleviates depression-like symptoms by rescuing decrease in neurogenesis in dorsal hippocampus via blocking microglia activation/phagocytosis. Brain Behav. Immun. 2021, 91, 519–530. [Google Scholar] [CrossRef] [PubMed]
- Wohleb, E.S.; Terwilliger, R.; Duman, C.H.; Duman, R.S. Microglia-Mediated Neuronal Remodeling and Depressive-like Behavior. Biol. Psychiatry 2018, 83, 38–49. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Sethi, R.; Mangala, L.S.; Taylor, C.; Goldsmith, J.; Wang, M.; Masuda, K.; Carrami, E.M.; Mannion, D.; Miranda, F.; et al. Tuning microtubule dynamics to enhance cancer therapy by modulating FER-mediated CRMP2 phosphorylation. Nat. Commun. 2018, 9, 476. [Google Scholar] [CrossRef]
- Wilson, S.M.; Moutal, A.; Melemedjian, O.K.; Wang, Y.; Ju, W.; François-Moutal, L.; Khanna, M.; Khanna, R. The functionalized amino acid (S)-Lacosamide subverts CRMP2-mediated tubulin polymerization to prevent constitutive and activity-dependent increase in neurite outgrowth. Front. Cell Neurosci. 2014, 8, 196. [Google Scholar] [CrossRef]
- Wu, Z.; Wang, G.; Wang, H.; Xiao, L.; Wei, Y.; Yang, C. Fluoxetine exposure for more than 2 days decreases the neuronal plasticity mediated by CRMP2 in differentiated PC12 cells. Brain Res. Bull. 2020, 158, 99–107. [Google Scholar] [CrossRef]
- Fang, W.; Gao, G.; Zhao, H.; Xia, Y.; Guo, X.; Li, N.; Li, Y.; Yang, Y.; Chen, L.; Wang, Q.; et al. Role of the Akt/GSK-3β/CRMP-2 pathway in axon degeneration of dopaminergic neurons resulting from MPP+ toxicity. Brain Res. 2015, 1602, 9–19. [Google Scholar] [CrossRef]
- Liu, N.; Wang, Y.; An, A.Y.; Banker, C.; Qian, Y.H.; O’Donnell, J.M. Single housing-induced effects on cognitive impairment and depression-like behavior in male and female mice involve neuroplasticity-related signaling. Eur. J. Neurosci. 2020, 52, 2694–2704. [Google Scholar] [CrossRef]
- Sun, L.; Wang, G.; Wu, Z.; Xie, Y.; Zhou, L.; Xiao, L.; Wang, H. Swimming exercise reduces the vulnerability to stress and contributes to the AKT/GSK3β/CRMP2 pathway and microtubule dynamics mediated protective effects on neuroplasticity in male C57BL/6 mice. Pharm. Biochem. Behav. 2021, 211, 173285. [Google Scholar] [CrossRef]
- Xu, X.; Xiao, X.; Yan, Y.; Zhang, T. Activation of liver X receptors prevents emotional and cognitive dysfunction by suppressing microglial M1-polarization and restoring synaptic plasticity in the hippocampus of mice. Brain Behav. Immun. 2021, 94, 111–124. [Google Scholar] [CrossRef]
- Xie, Y.; Wu, Z.; Sun, L.; Zhou, L.; Xiao, L.; Wang, H.; Wang, G. Swimming exercise reverses chronic unpredictable mild stress-induced depression-like behaviors and alleviates neuroinflammation and collapsing response mediator protein-2-mediated neuroplasticity injury in adult male mice. Neuroreport 2022, 33, 272–282. [Google Scholar] [CrossRef] [PubMed]
- O’Hara, M.W.; McCabe, J.E. Postpartum depression: Current status and future directions. Annu. Rev. Clin. Psychol. 2013, 9, 379–407. [Google Scholar] [CrossRef] [PubMed]
- Zajdel, J.; Zager, A.; Blomqvist, A.; Engblom, D.; Shionoya, K. Acute maternal separation potentiates the gene expression and corticosterone response induced by inflammation. Brain Behav. Immun. 2019, 77, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Busillo, J.M.; Azzam, K.M.; Cidlowski, J.A. Glucocorticoids sensitize the innate immune system through regulation of the NLRP3 inflammasome. J. Biol. Chem. 2011, 286, 38703–38713. [Google Scholar] [CrossRef] [PubMed]
- Delpech, J.C.; Wei, L.; Hao, J.; Yu, X.; Madore, C.; Butovsky, O.; Kaffman, A. Early life stress perturbs the maturation of microglia in the developing hippocampus. Brain Behav. Immun. 2016, 57, 79–93. [Google Scholar] [CrossRef]
- Walker, F.R.; Nilsson, M.; Jones, K. Acute and chronic stress-induced disturbances of microglial plasticity, phenotype and function. Curr. Drug Targets 2013, 14, 1262–1276. [Google Scholar] [CrossRef]
- Malcon, L.; Wearick-Silva, L.E.; Zaparte, A.; Orso, R.; Luft, C.; Tractenberg, S.G.; Donadio, M.; de Oliveira, J.R.; Grassi-Oliveira, R. Maternal separation induces long-term oxidative stress alterations and increases anxiety-like behavior of male Balb/cJ mice. Exp. Brain Res. 2020, 238, 2097–2107. [Google Scholar] [CrossRef]
- Reshetnikov, V.V.; Kovner, A.V.; Lepeshko, A.A.; Pavlov, K.S.; Grinkevich, L.N.; Bondar, N.P. Stress early in life leads to cognitive impairments, reduced numbers of CA3 neurons and altered maternal behavior in adult female mice. Genes Brain Behav. 2020, 19, e12541. [Google Scholar] [CrossRef]
- Liu, W.; Zhang, A.; He, H.; Zhao, X.; Tao, F.; Sun, Y. Inflammatory burden in adolescents with prolonged parent-child separation. Brain Behav. Immun. 2021, 98, 257–262. [Google Scholar] [CrossRef]
- Brierley, S.M.; Linden, D.R. Neuroplasticity and dysfunction after gastrointestinal inflammation. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 611–627. [Google Scholar] [CrossRef]
- Levy, M.; Boulle, F.; Emerit, M.B.; Poilbout, C.; Steinbusch, H.; Van den Hove, D.; Kenis, G.; Lanfumey, L. 5-HTT independent effects of fluoxetine on neuroplasticity. Sci. Rep. 2019, 9, 6311. [Google Scholar] [CrossRef]
- Wu, Z.; Wang, G.; Xiao, L.; Wei, Y.; Wang, H.; Zhou, L.; Sun, L. Resilience in the LPS-induced acute depressive-like behaviors: Increase of CRMP2 neuroprotection and microtubule dynamics in hippocampus. Brain Res. Bull. 2020, 162, 261–270. [Google Scholar] [CrossRef] [PubMed]
- Bale, T.L.; Baram, T.Z.; Brown, A.S.; Goldstein, J.M.; Insel, T.R.; McCarthy, M.M.; Nemeroff, C.B.; Reyes, T.M.; Simerly, R.B.; Susser, E.S.; et al. Early life programming and neurodevelopmental disorders. Biol. Psychiatry 2010, 68, 314–319. [Google Scholar] [CrossRef] [PubMed]
- Rincón-Cortés, M.; Sullivan, R.M. Early life trauma and attachment: Immediate and enduring effects on neurobehavioral and stress axis development. Front. Endocrinol. 2014, 5, 33. [Google Scholar]
- Nguyen, P.T.; Dorman, L.C.; Pan, S.; Vainchtein, I.D.; Han, R.T.; Nakao-Inoue, H.; Taloma, S.E.; Barron, J.J.; Molofsky, A.B.; Kheirbek, M.A.; et al. Microglial Remodeling of the Extracellular Matrix Promotes Synapse Plasticity. Cell 2020, 182, 388–403.e15. [Google Scholar] [CrossRef]
- Singhal, G.; Baune, B.T. Microglia: An Interface between the Loss of Neuroplasticity and Depression. Front. Cell Neurosci. 2017, 11, 270. [Google Scholar] [CrossRef]
- Largo-Barrientos, P.; Apóstolo, N.; Creemers, E.; Callaerts-Vegh, Z.; Swerts, J.; Davies, C.; McInnes, J.; Wierda, K.; De Strooper, B.; Spires-Jones, T.; et al. Lowering Synaptogyrin-3 expression rescues Tau-induced memory defects and synaptic loss in the presence of microglial activation. Neuron 2021, 109, 767–777.e5. [Google Scholar] [CrossRef]
- Lonnemann, N.; Hosseini, S.; Marchetti, C.; Skouras, D.B.; Stefanoni, D.; D’Alessandro, A.; Dinarello, C.A.; Korte, M. The NLRP3 inflammasome inhibitor OLT1177 rescues cognitive impairment in a mouse model of Alzheimer’s disease. Proc. Natl. Acad. Sci. USA 2020, 117, 32145–32154. [Google Scholar] [CrossRef]
- Chen, K.; Hu, Q.; Xie, Z.; Yang, G. Monocyte NLRP3-IL-1β Hyperactivation Mediates Neuronal and Synaptic Dysfunction in Perioperative Neurocognitive Disorder. Adv. Sci. 2022, 9, 2104106. [Google Scholar] [CrossRef]
- Nagai, J.; Owada, K.; Kitamura, Y.; Goshima, Y.; Ohshima, T. Inhibition of CRMP2 phosphorylation repairs CNS by regulating neurotrophic and inhibitory responses. Exp. Neurol. 2016, 277, 283–295. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, L.; Wu, Z.; Li, Y.; Xiao, L.; Wang, H.; Wang, G. Brief Maternal Separation Promotes Resilience to Anxiety-like and Depressive-like Behaviors in Female C57BL/6J Offspring with Imiquimod-Induced Psoriasis. Brain Sci. 2022, 12, 1250. https://doi.org/10.3390/brainsci12091250
Zhou L, Wu Z, Li Y, Xiao L, Wang H, Wang G. Brief Maternal Separation Promotes Resilience to Anxiety-like and Depressive-like Behaviors in Female C57BL/6J Offspring with Imiquimod-Induced Psoriasis. Brain Sciences. 2022; 12(9):1250. https://doi.org/10.3390/brainsci12091250
Chicago/Turabian StyleZhou, Lin, Zuotian Wu, Yixin Li, Ling Xiao, Huiling Wang, and Gaohua Wang. 2022. "Brief Maternal Separation Promotes Resilience to Anxiety-like and Depressive-like Behaviors in Female C57BL/6J Offspring with Imiquimod-Induced Psoriasis" Brain Sciences 12, no. 9: 1250. https://doi.org/10.3390/brainsci12091250