
Effects of Long-Term Exposure to 2260 m Altitude on Working Memory and Resting-State Activity in the Prefrontal Cortex: A Large-Sample Cross-Sectional Study

Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
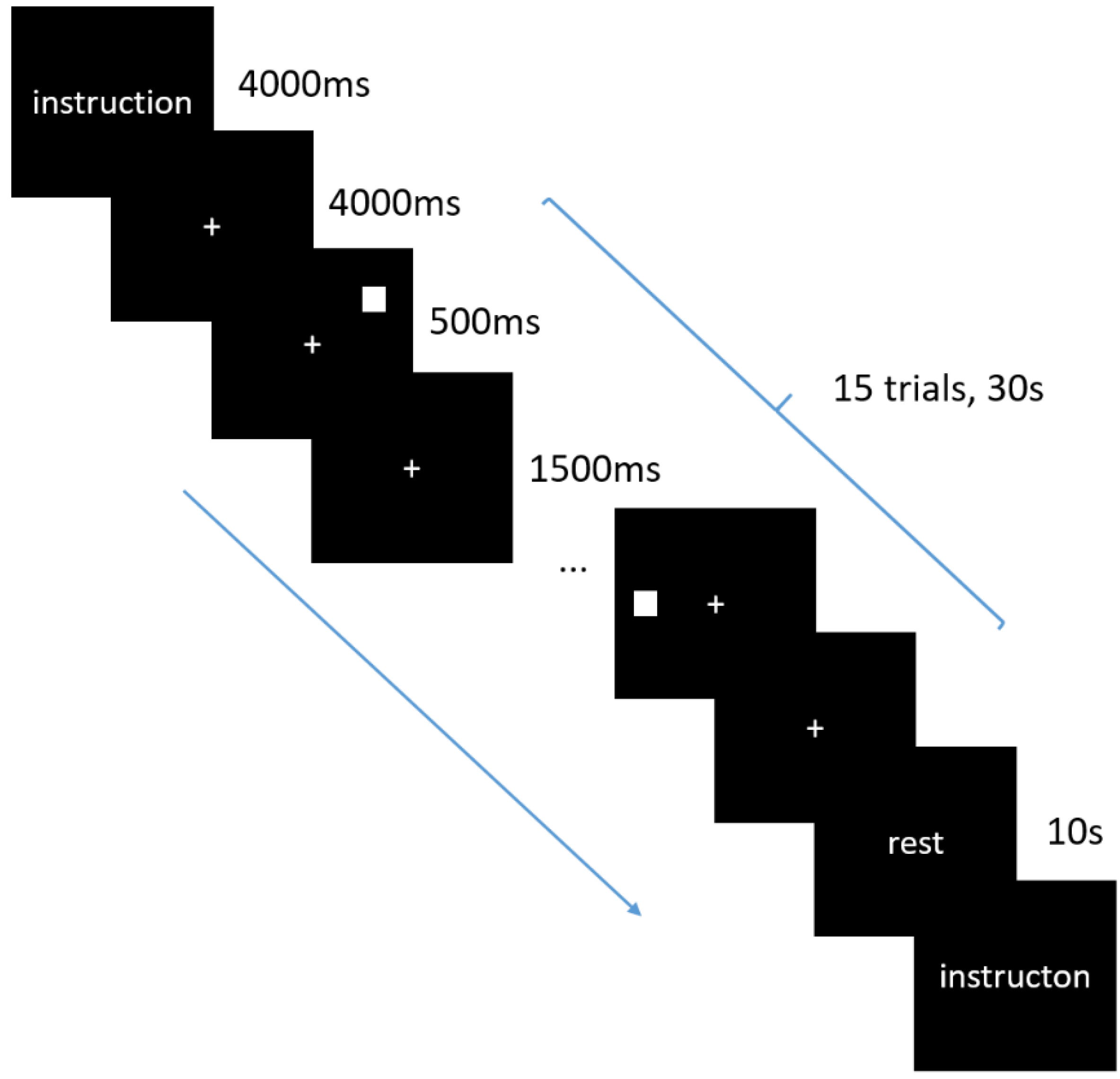
2.2. Physiological and Behavior Tests
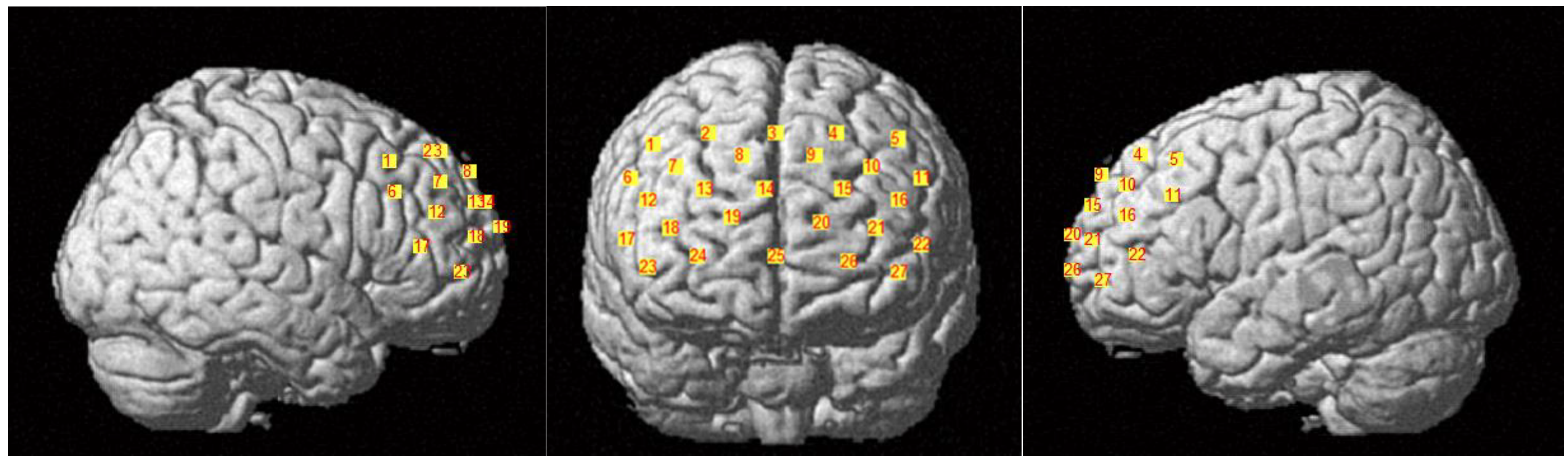
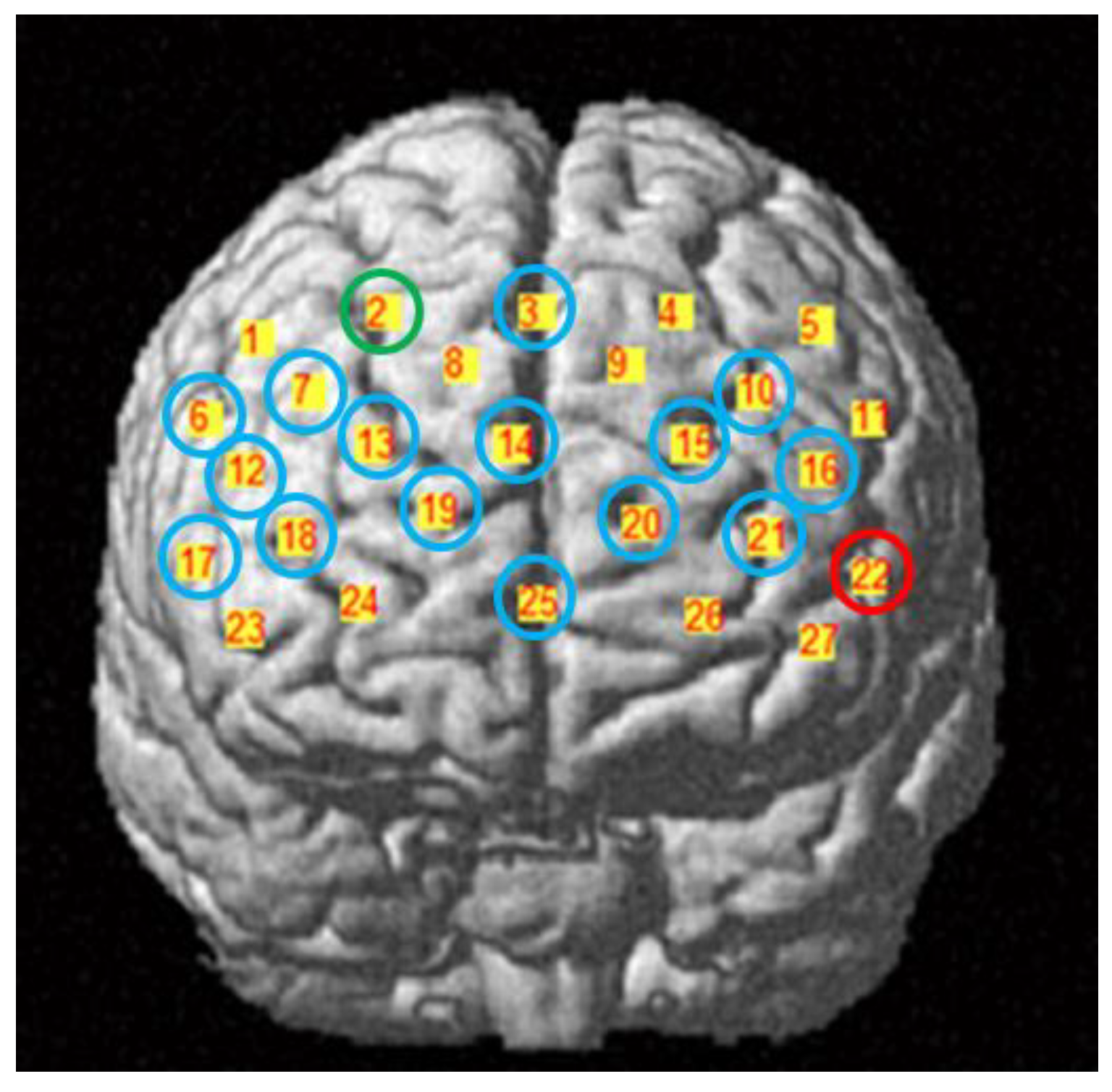
2.3. fNIRS Data Acquisition
2.4. fNIRS Data Preprocessing
2.5. Network Construction
2.6. Network Analysis
2.7. Complexity Analysis
2.8. Statistical Analysis
2.9. Relationship between Behavior Performance and Brain Activity
3. Results
3.1. Behavior Results
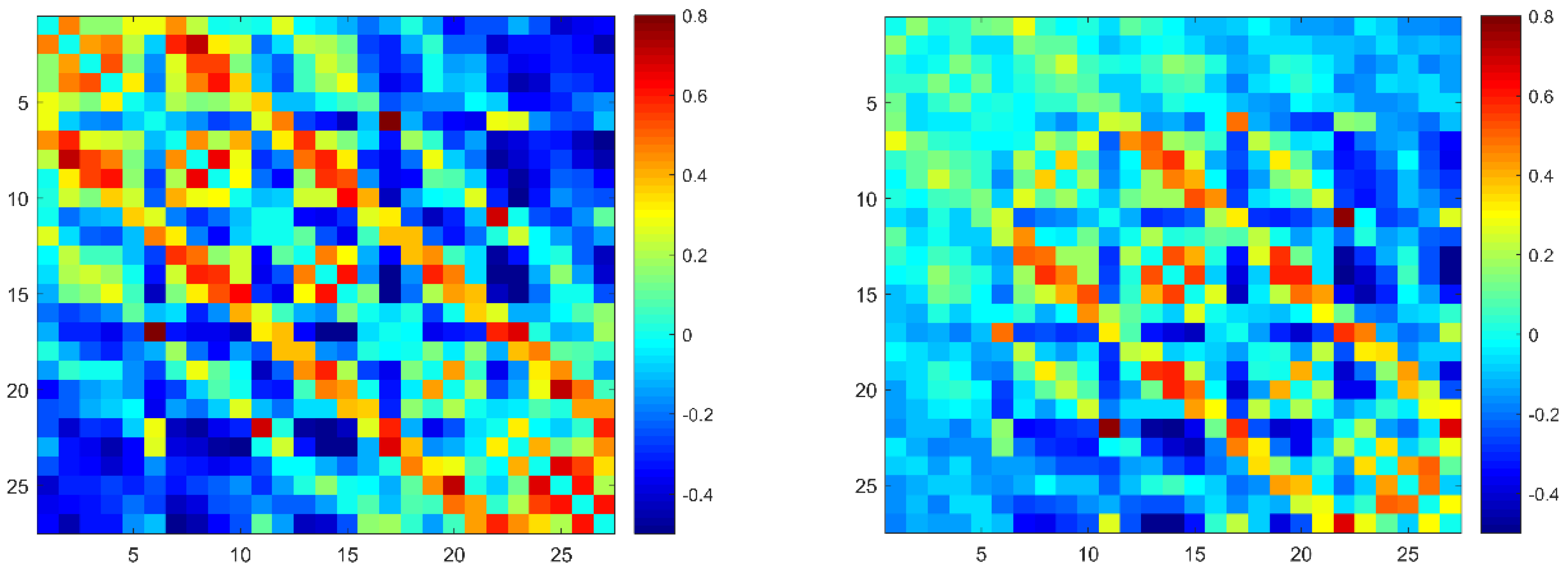
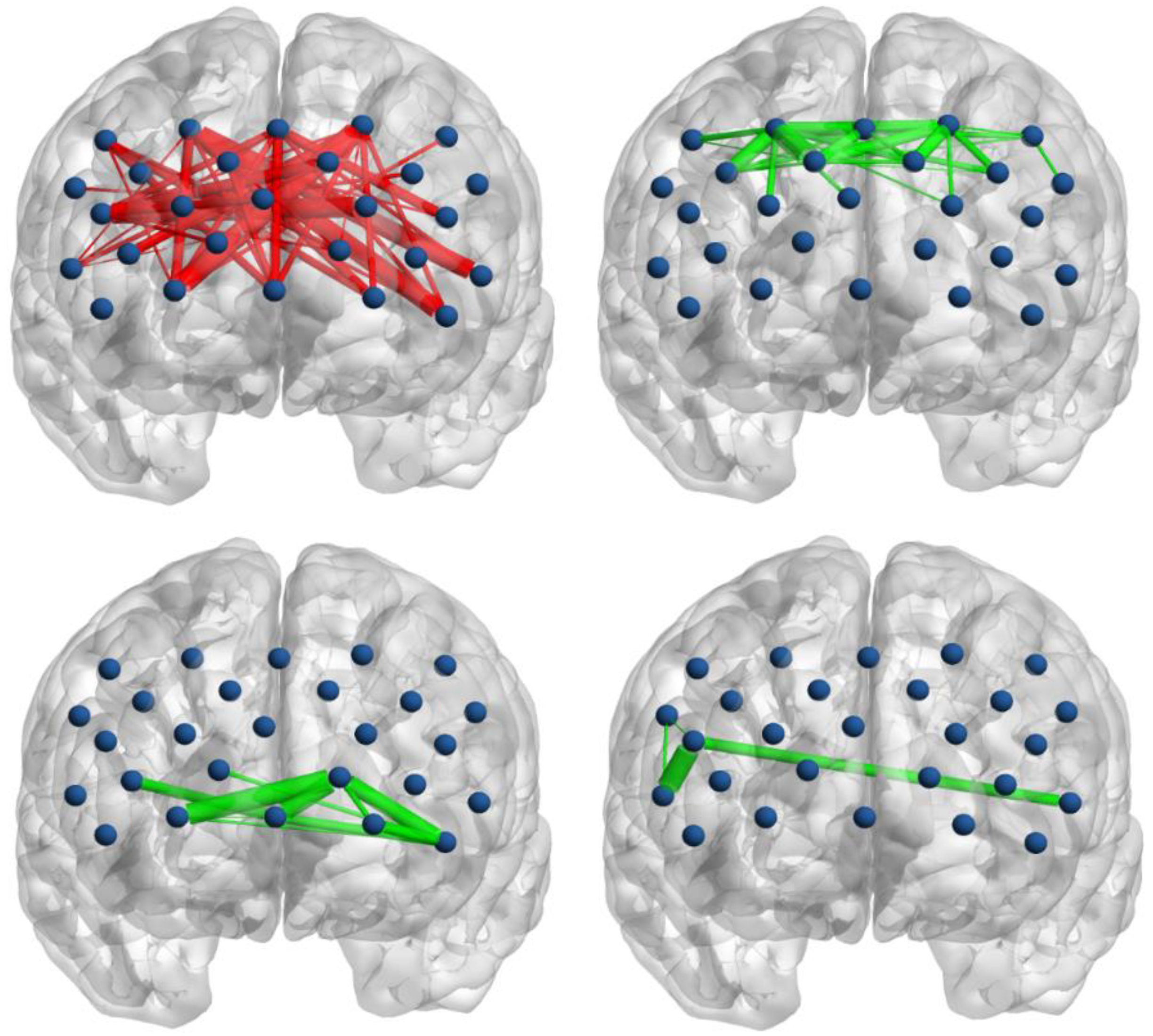
3.2. Functional Connectivity
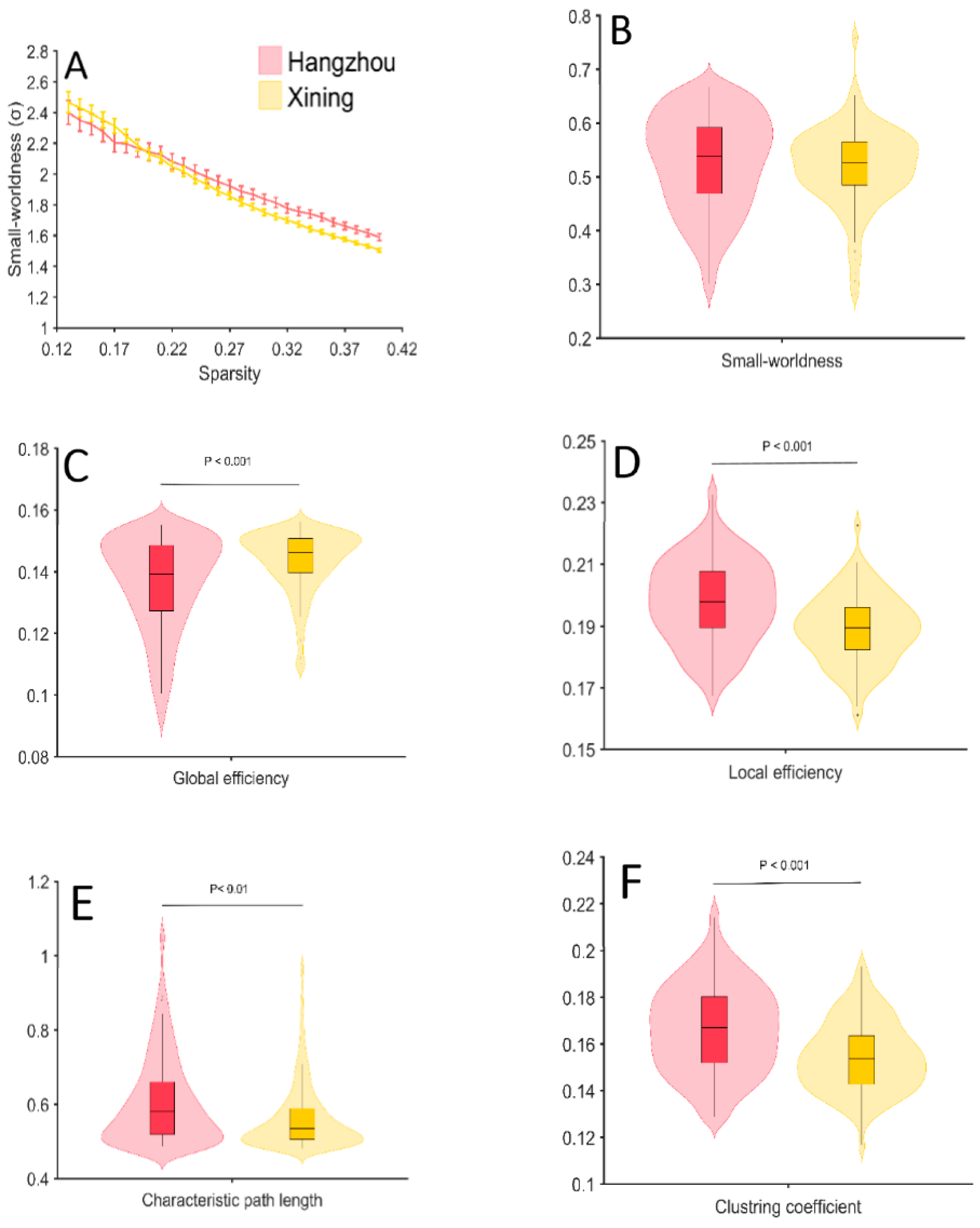
3.3. Global Network Properties
3.4. Regional Nodal Properties
3.5. Complexity
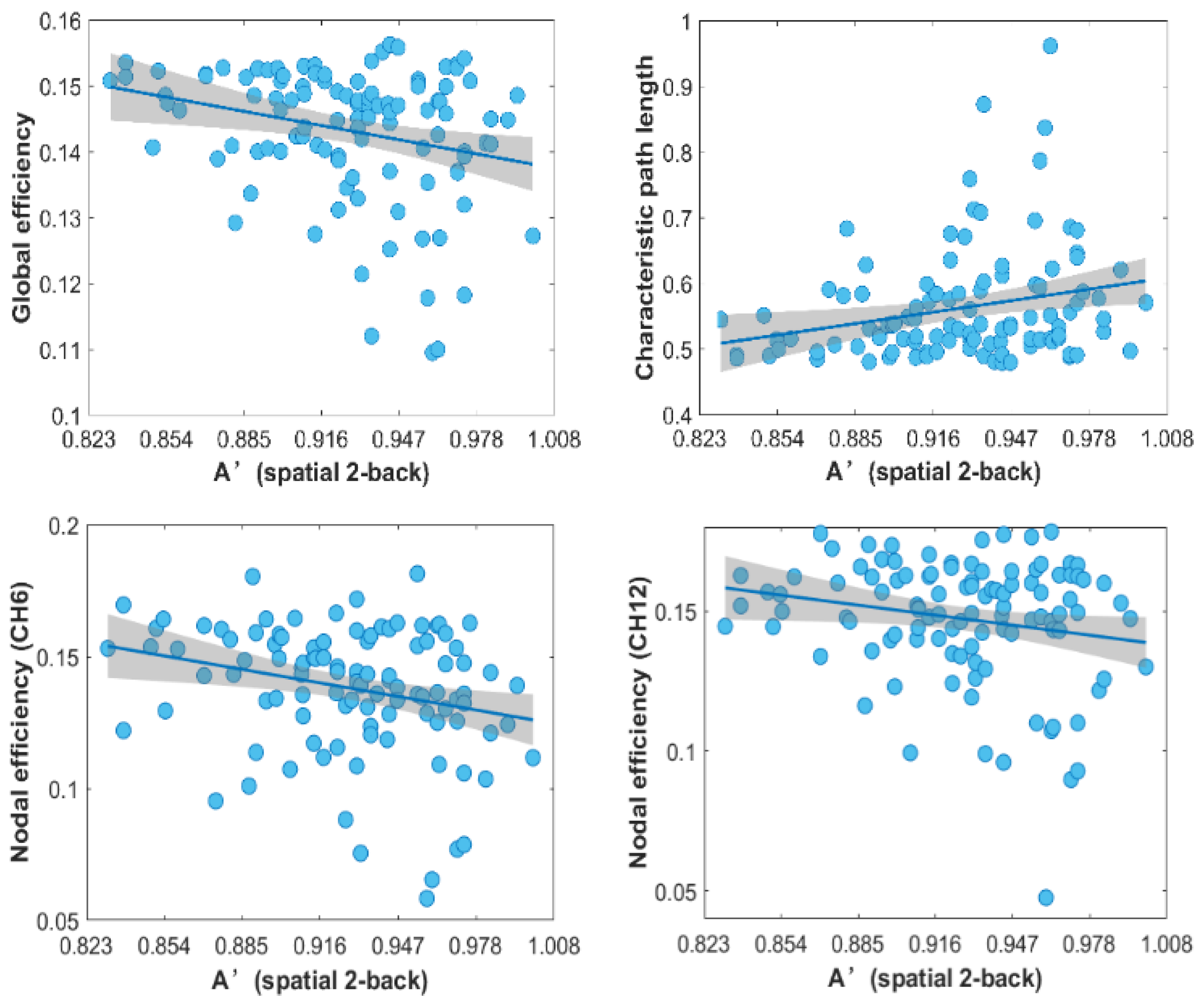
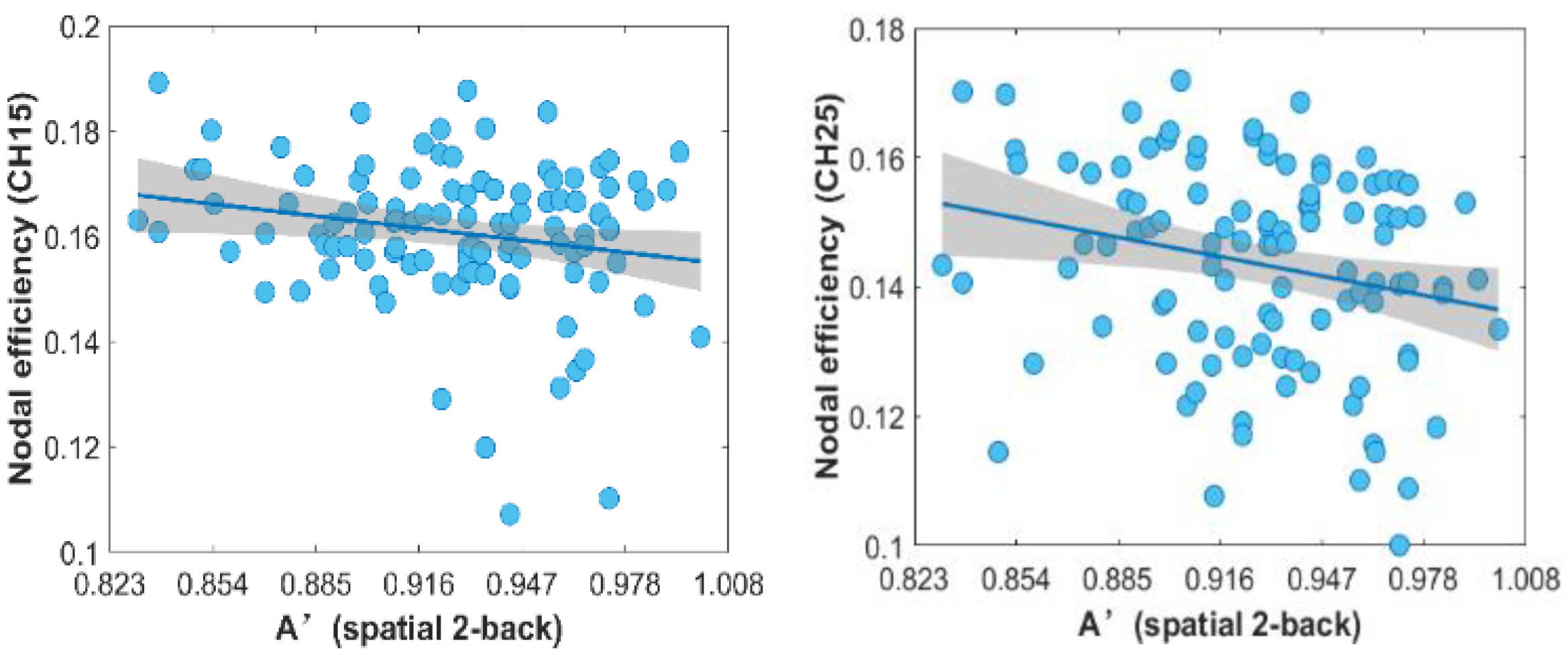
3.6. Correlation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tremblay, J.C.; Ainslie, P.N. Global and country-level estimates of human population at high altitude. Proc. Natl. Acad. Sci. USA 2021, 118, e2102463118. [Google Scholar] [CrossRef] [PubMed]
- Mujika, I.; Sharma, A.P.; Stellingwerff, T. Contemporary Periodization of Altitude Training for Elite Endurance Athletes: A Narrative Review. Sports Med. 2019, 49, 1651–1669. [Google Scholar] [CrossRef] [PubMed]
- Taylor, L.; Watkins, S.L.; Marshall, H.; Dascombe, B.J.; Foster, J. The Impact of Different Environmental Conditions on Cognitive Function: A Focused Review. Front. Physiol. 2016, 6, 372. [Google Scholar] [CrossRef] [PubMed]
- Shaw, D.M.; Cabre, G.; Gant, N. Hypoxic Hypoxia and Brain Function in Military Aviation: Basic Physiology and Applied Perspectives. Front. Physiol. 2021, 12, 698. [Google Scholar] [CrossRef]
- Virues-Ortega, J.; Garrido, E.; Javierre, C.; Kloezeman, K.C. Human behaviour and development under high-altitude conditions. Dev. Sci. 2006, 9, 400–410. [Google Scholar] [CrossRef]
- McMorris, T.; Hale, B.J.; Barwood, M.; Costello, J.; Corbett, J. Effect of acute hypoxia on cognition: A systematic review and meta-regression analysis. Neurosci. Biobehav. Rev. 2017, 74, 225–232. [Google Scholar] [CrossRef]
- Pun, M.; Guadagni, V.; Bettauer, K.M.; Drogos, L.L.; Aitken, J.; Hartmann, S.E.; Furian, M.; Muralt, L.; Lichtblau, M.; Bader, P.R.; et al. Effects on Cognitive Functioning of Acute, Subacute and Repeated Exposures to High Altitude. Front. Physiol. 2018, 9, 1131. [Google Scholar] [CrossRef]
- Altbacker, A.; Takacs, E.; Barkaszi, I.; Kormos, T.; Czigler, I.; Balazs, L. Differential impact of acute hypoxia on event related potentials: Impaired task-irrelevant, but preserved task-relevant processing and response inhibition. Physiol. Behav. 2019, 206, 28–36. [Google Scholar] [CrossRef]
- Thakur, L.; Ray, K.; Anand, J.P.; Panjwani, U. Event related potential (ERP) P300 after 6 months residence at 4115 meter. Indian J. Med. Res. 2011, 134, 113–117. [Google Scholar]
- Legg, S.; Hill, S.; Gilbey, A.; Raman, A.; Schlader, Z.; Mündel, T. Effect of Mild Hypoxia on Working Memory, Complex Logical Reasoning, and Risk Judgment. Int. J. Aviat. Psychol. 2014, 24, 126–140. [Google Scholar] [CrossRef]
- Thropp, J.E.; Buza, P.W. Cumulative Cyclic Exposures to 8000-ft Pressurization Equivalence and Attention Network Responses. Aerosp. Med. Hum. Perform. 2019, 90, 513–523. [Google Scholar] [CrossRef] [PubMed]
- Hewett, K.J.; Curry, I.P.; Rath, E.; Collins, S.M. Subtle Cognitive Effects of Moderate Hypoxia; Army Aeromedical Research Lab.: Fort Rucker, AL, USA, 2009. [Google Scholar]
- Bouak, F.; Vartanian, O.; Hofer, K.; Cheung, B. Acute Mild Hypoxic Hypoxia Effects on Cognitive and Simulated Aircraft Pilot Performance. Aerosp. Med. Hum. Perform. 2018, 89, 526–535. [Google Scholar] [CrossRef] [PubMed]
- Parker, P.J.; Manley, A.J.; Shand, R.; O’Hara, J.R.; Mellor, A. Working Memory Capacity and Surgical Performance While Exposed to Mild Hypoxic Hypoxemia. Aerosp. Med. Hum. Perform. 2017, 88, 918–923. [Google Scholar] [CrossRef]
- Komiyama, T.; Sudo, M.; Higaki, Y.; Kiyonaga, A.; Tanaka, H.; Ando, S. Does moderate hypoxia alter working memory and executive function during prolonged exercise? Physiol. Behav. 2015, 139, 290–296. [Google Scholar] [CrossRef] [PubMed]
- Schlaepfer, T.E.; Bärtsch, P.; Fisch, H.U.J.C.S. Paradoxical effects of mild hypoxia and moderate altitude on human visual perception. Clin. Sci. 1992, 83, 633–636. [Google Scholar] [CrossRef] [PubMed]
- Petrassi, F.A.; Hodkinson, P.D.; Walters, P.L.; Gaydos, S.J. Hypoxic Hypoxia at Moderate Altitudes: Review of the State of the Science. Aviat. Space Environ. Med. 2012, 83, 975–984. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.X.; Liu, H.C.; Yan, X.D.; Weng, X.C. Minimal Effects on Human Memory Following Long-Term Living at Moderate Altitude. High Alt. Med. Biol. 2011, 12, 37–43. [Google Scholar] [CrossRef]
- Wei, X.; Ni, X.; Zhao, S.; Chi, A. Influence of Exposure at Different Altitudes on the Executive Function of Plateau Soldiers-Evidence from ERPs and Neural Oscillations. Front. Physiol. 2021, 12, 632058. [Google Scholar] [CrossRef]
- Chen, X.; Liu, J.; Wang, J.; Xin, Z.; Zhang, Q.; Zhang, W.; Xi, Y.; Zhu, Y.; Li, C.; Li, J.; et al. Altered resting-state networks may explain the executive impairment in young health immigrants into high-altitude area. Brain Imaging Behav. 2020, 15, 147–156. [Google Scholar] [CrossRef]
- Zhang, X.; Kang, T.; Liu, Y.; Yuan, F.; Li, M.; Lin, J.; Zhang, J.J. Resting-State Neuronal Activity and Functional Connectivity Changes in the Visual Cortex after High Altitude Exposure: A Longitudinal Study. Brain Sci. 2022, 12, 724. [Google Scholar]
- Davranche, K.; Casini, L.; Arnal, P.J.; Rupp, T.; Perrey, S.; Verges, S. Cognitive functions and cerebral oxygenation changes during acute and prolonged hypoxic exposure. Physiol. Behav. 2016, 164, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Lawley, J.S.; Macdonald, J.H.; Oliver, S.J.; Mullins, P.G. Unexpected reductions in regional cerebral perfusion during prolonged hypoxia. J. Physiol. 2017, 595, 935–947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedman, N.P.; Robbins, T.W. The role of prefrontal cortex in cognitive control and executive function. Neuropsychopharmacology 2022, 47, 72–89. [Google Scholar] [CrossRef] [PubMed]
- Racz, F.S.; Mukli, P.; Nagy, Z.; Eke, A. Increased prefrontal cortex connectivity during cognitive challenge assessed by fNIRS imaging. Biomed. Opt. Express 2017, 8, 3842–3855. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Liang, M.; Wang, L.; Tian, L.; Zhang, X.; Li, K.; Jiang, T. Altered functional connectivity in early Alzheimer’s disease: A resting-state fMRI study. Hum. Brain Mapp. 2007, 28, 967–978. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Zhang, J.; Gong, Q.; Weng, X. Adaptive influence of long term high altitude residence on spatial working memory: An fMRI study. Brain Cogn. 2011, 77, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Huppert, T.J.; Diamond, S.G.; Franceschini, M.A.; Boas, D.A. HomER: A review of time-series analysis methods for near-infrared spectroscopy of the brain. Appl. Opt. 2009, 48, D280–D298. [Google Scholar] [CrossRef]
- Long, Y.; Zheng, L.; Zhao, H.; Zhou, S.; Zhai, Y.; Lu, C. Interpersonal Neural Synchronization during Interpersonal Touch Underlies Affiliative Pair Bonding between Romantic Couples. Cereb. Cortex 2021, 31, 1647–1659. [Google Scholar] [CrossRef]
- Hoshi, Y. Functional near-infrared spectroscopy: Current status and future prospects. J. Biomed. Opt. 2007, 12, 062106. [Google Scholar] [CrossRef]
- Wang, J.; Wang, X.; Xia, M.; Liao, X.; Evans, A.; He, Y. GRETNA: A graph theoretical network analysis toolbox for imaging connectomics. Front. Hum. Neurosci. 2015, 9, 386. [Google Scholar] [CrossRef]
- Hou, X.; Zhang, Z.; Zhao, C.; Duan, L.; Gong, Y.; Li, Z.; Zhu, C. NIRS-KIT: A MATLAB toolbox for both resting-state and task fNIRS data analysis. Neurophotonics 2021, 8, 010802. [Google Scholar] [CrossRef] [PubMed]
- Achard, S.; Salvador, R.; Whitcher, B.; Suckling, J.; Bullmore, E.D. A resilient, low-frequency, small-world human brain functional network with highly connected association cortical hubs. J. Neurosci. 2006, 26, 63–72. [Google Scholar] [CrossRef]
- Wang, J.; Wang, L.; Zang, Y.; Yang, H.; Tang, H.; Gong, Q.; Chen, Z.; Zhu, C.; He, Y. Parcellation-dependent small-world brain functional networks: A resting-state fMRI study. Hum. Brain Mapp. 2009, 30, 1511–1523. [Google Scholar] [CrossRef] [PubMed]
- Cai, L.; Dong, Q.; Niu, H. The development of functional network organization in early childhood and early adolescence: A resting-state fNIRS study. Dev. Cogn. Neurosci. 2018, 30, 223–235. [Google Scholar] [CrossRef] [PubMed]
- Rubinov, M.; Sporns, O. Complex network measures of brain connectivity: Uses and interpretations. Neuroimage 2010, 52, 1059–1069. [Google Scholar] [CrossRef]
- Hager, B.; Yang, A.C.; Brady, R.; Meda, S.; Clementz, B.; Pearlson, G.D.; Sweeney, J.A.; Tamminga, C.; Keshavan, M. Neural complexity as a potential translational biomarker for psychosis. J. Affect. Disord. 2017, 216, 89–99. [Google Scholar] [CrossRef]
- Snodgrass, J.G.; Corwin, J. Pragmatics of measuring recognition memory: Applications to dementia and amnesia. J. Exp. Psychol. Gen. 1988, 117, 34–50. [Google Scholar] [CrossRef]
- Xia, M.; Wang, J.; He, Y. BrainNet Viewer: A network visualization tool for human brain connectomics. PLoS ONE 2013, 8, e68910. [Google Scholar] [CrossRef]
- Ma, H.; Zhang, D.; Li, X.; Ma, H.; Wang, N.; Wang, Y. Long-term exposure to high altitude attenuates verbal and spatial working memory: Evidence from an event-related potential study. Brain Behav. 2019, 9, e01256. [Google Scholar] [CrossRef]
- Badre, D. Cognitive control, hierarchy, and the rostro–caudal organization of the frontal lobes. Trends Cogn. Sci. 2008, 12, 193–200. [Google Scholar] [CrossRef]
- Miller, E.K.; Cohen, J.D. An integrative theory of prefrontal cortex function. Annu. Rev. Neurosci. 2001, 24, 167–202. [Google Scholar] [CrossRef] [PubMed]
- Badre, D.; Nee, D.E. Frontal Cortex and the Hierarchical Control of Behavior. Trends Cogn. Sci. 2018, 22, 170–188. [Google Scholar] [CrossRef] [PubMed]
- Latora, V.; Marchiori, M. Efficient behavior of small-world networks. Phys. Rev. Lett. 2001, 87, 198701. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hangzhou (n = 91) | Xining (n = 117) | p (t/χ2) | |
|---|---|---|---|
| Age | 20.22 ± 0.48 | 20.41 ± 0.90 | >0.05 |
| Female/Male | 46/45 | 65/52 | >0.05 |
| RPMs | 60.34 ± 29.29 | 57.61 ± 27.23 | >0.05 |
| SpO2 | 98.68 ± 0.99 | 96.76 ± 1.26 | <0.001 |
| HR | 80.55 ± 11.10 | 77.12 ± 9.43 | <0.05 |
| Channel | MNI | Brain Regions (Percentage of Overlap) | Brodmann Area | ||
|---|---|---|---|---|---|
| X | Y | Z | |||
| 1 | 47 | 27 | 47 | Dorsolateral prefrontal cortex (0.63) | 9 |
| 2 | 25 | 42 | 50 | Includes frontal eye fields (0.79) | 8 |
| 3 | 2 | 47 | 50 | Includes frontal eye fields (0.75) | 8 |
| 4 | −20 | 43 | 51 | Includes frontal eye fields (0.83) | 8 |
| 5 | −42 | 28 | 48 | Includes frontal eye fields (0.61) | 8 |
| 6 | 55 | 27 | 34 | Dorsolateral prefrontal cortex (0.90) | 46 |
| 7 | 38 | 45 | 38 | Dorsolateral prefrontal cortex (0.69) | 9 |
| 8 | 15 | 57 | 41 | Dorsolateral prefrontal cortex (0.83) | 9 |
| 9 | −11 | 58 | 41 | Dorsolateral prefrontal cortex (0.85) | 9 |
| 10 | −33 | 48 | 38 | Dorsolateral prefrontal cortex (0.80) | 9 |
| 11 | −50 | 30 | 35 | Dorsolateral prefrontal cortex (0.51) | 46 |
| 12 | 48 | 45 | 27 | Frontopolar area (0.72) | 10 |
| 13 | 27 | 60 | 29 | Frontopolar area (0.92) | 10 |
| 14 | 6 | 65 | 31 | Frontopolar area (0.90) | 10 |
| 15 | −21 | 62 | 29 | Frontopolar area (0.84) | 10 |
| 16 | −42 | 48 | 26 | Dorsolateral prefrontal cortex (0.66) | 46 |
| 17 | 56 | 39 | 12 | Dorsolateral prefrontal cortex (0.39) | 46 |
| 18 | 41 | 60 | 16 | Frontopolar area (1) | 10 |
| 19 | 18 | 71 | 19 | Frontopolar area (1) | 10 |
| 20 | −14 | 71 | 17 | Frontopolar area (1) | 10 |
| 21 | −34 | 62 | 15 | Frontopolar area (0.97) | 10 |
| 22 | −51 | 44 | 10 | Dorsolateral prefrontal cortex (0.56) | 46 |
| 23 | 48 | 55 | 1 | Frontopolar area (0.88) | 10 |
| 24 | 29 | 69 | 6 | Frontopolar area (1) | 10 |
| 25 | 3 | 70 | 6 | Frontopolar area (1) | 10 |
| 26 | −23 | 71 | 4 | Frontopolar area (1) | 10 |
| 27 | −42 | 59 | −1 | Frontopolar area (0.78) | 10 |
| Group | 0-Back | 1-Back | 2-Back | |
|---|---|---|---|---|
| A’ (unit) | Hangzhou | 0.948 ± 0.004 | 0.941 ± 0.004 | 0.929 ± 0.005 |
| A’ (unit) | Xining | 0.941 ± 0.003 | 0.936 ± 0.003 | 0.923 ± 0.004 |
| hitRT (ms) | Hangzhou | 471.48 ± 14.05 | 550.84 ± 16.91 | 749.96 ± 24.68 |
| hitRT (ms) | Xining | 460.56 ± 10.80 | 545.38 ± 12.99 | 696.81 ± 18.95 |
| rejRT (ms) | Hangzhou | 515.99 ± 14.31 | 612.06 ± 19.02 | 846.00 ± 25.80 |
| rejRT (ms) | Xining | 504.09 ± 11.00 | 603.47 ± 14.61 | 792.51 ± 19.81 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, X.; Zhou, A.; Li, J.; Chen, B.; Zhou, X.; Ma, H.; Lu, C.; Weng, X. Effects of Long-Term Exposure to 2260 m Altitude on Working Memory and Resting-State Activity in the Prefrontal Cortex: A Large-Sample Cross-Sectional Study. Brain Sci. 2022, 12, 1148. https://doi.org/10.3390/brainsci12091148
Chen X, Zhou A, Li J, Chen B, Zhou X, Ma H, Lu C, Weng X. Effects of Long-Term Exposure to 2260 m Altitude on Working Memory and Resting-State Activity in the Prefrontal Cortex: A Large-Sample Cross-Sectional Study. Brain Sciences. 2022; 12(9):1148. https://doi.org/10.3390/brainsci12091148
Chicago/Turabian StyleChen, Xin, Aibao Zhou, Junle Li, Bing Chen, Xin Zhou, Hailin Ma, Chunming Lu, and Xuchu Weng. 2022. "Effects of Long-Term Exposure to 2260 m Altitude on Working Memory and Resting-State Activity in the Prefrontal Cortex: A Large-Sample Cross-Sectional Study" Brain Sciences 12, no. 9: 1148. https://doi.org/10.3390/brainsci12091148