
Cortical Source Analysis of the Face Sensitive N290 ERP Component in Infants at High Risk for Autism


Abstract
:1. Introduction
1.1. The Infant N290
1.2. Cortical Source Analysis
1.3. Head Model Selection
1.4. The Current Study
2. Materials and Methods
2.1. Participants
2.2. Procedure
2.2.1. EEG Recording
2.2.2. Magnetic Resonance Imaging
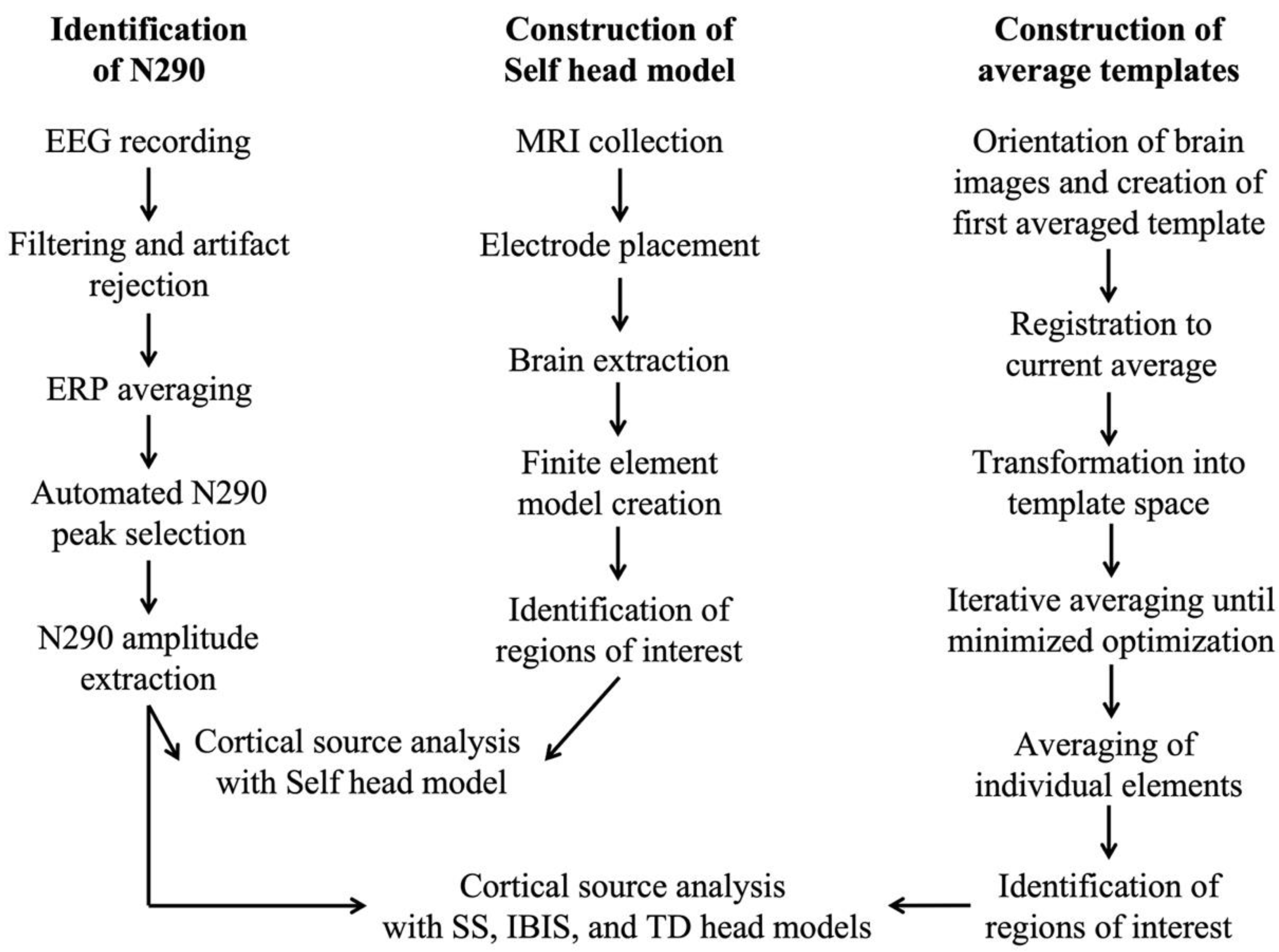
2.3. Data Processing
2.3.1. MRI File Preparation
2.3.2. Construction of Group- and Study-Specific Templates
2.3.3. Electrode Placement
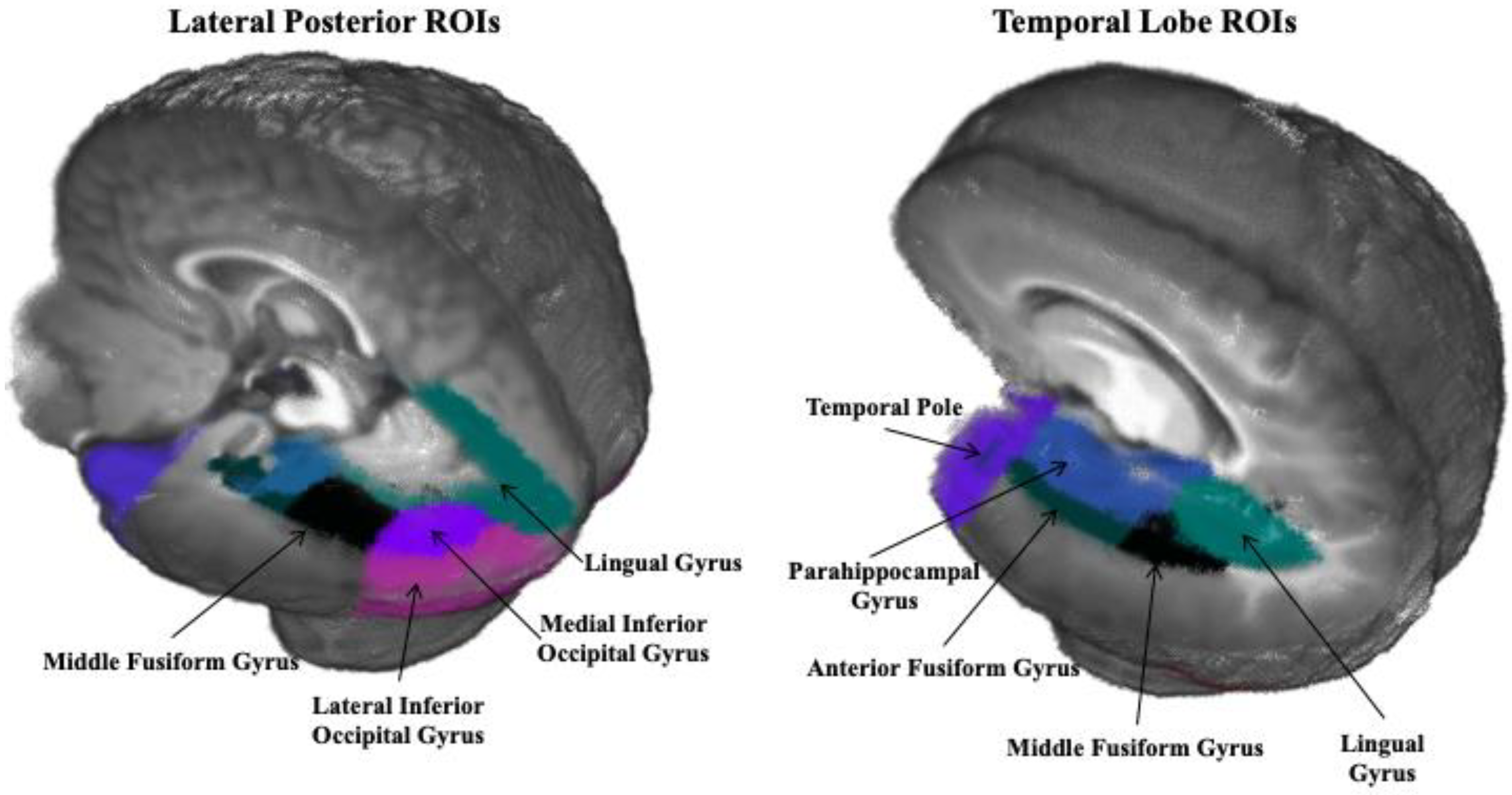
2.3.4. Regions of Interest
2.4. Data Analysis
2.4.1. Head Model Comparison
2.4.2. Cortical Source Analysis
3. Results
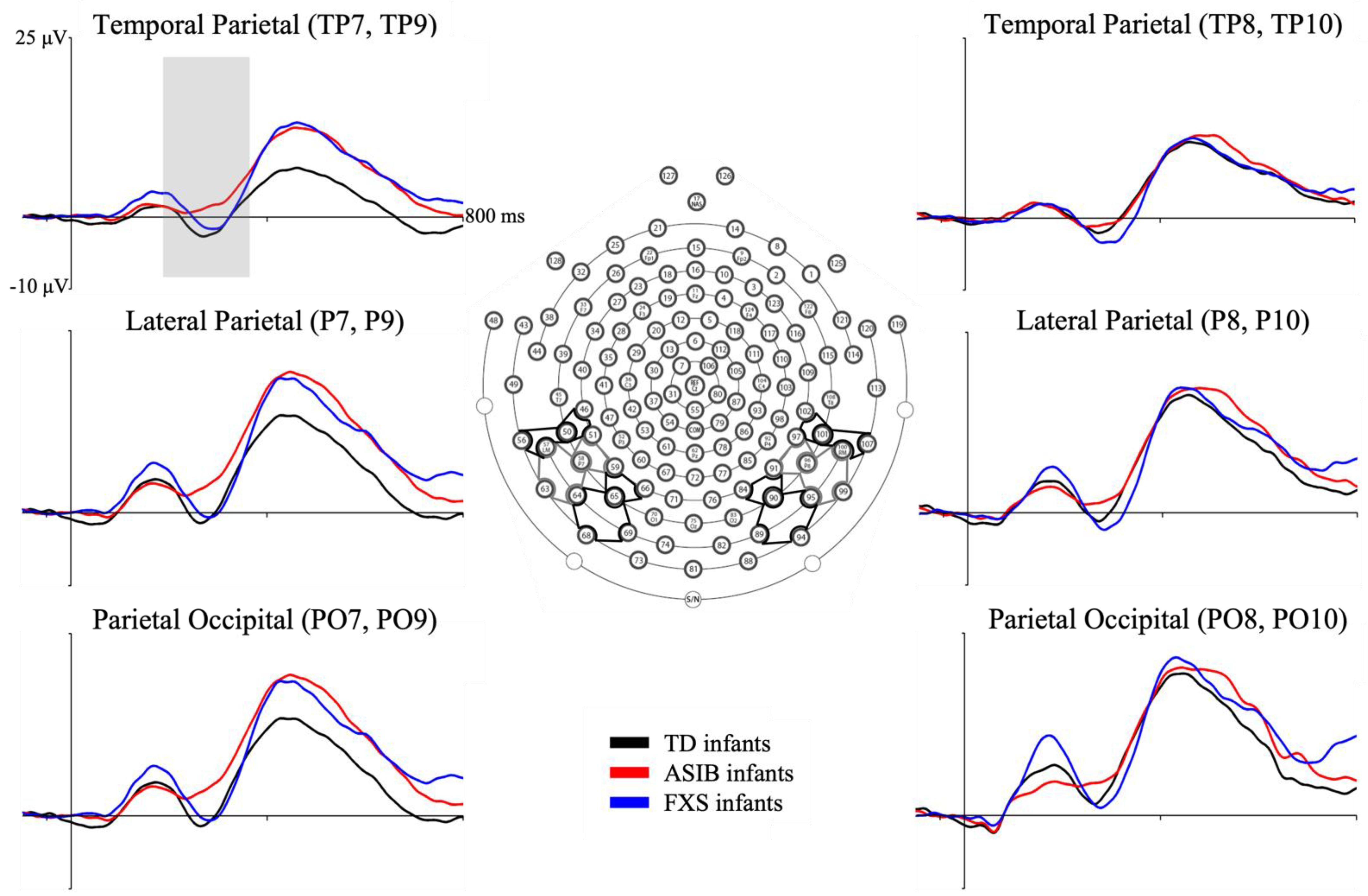
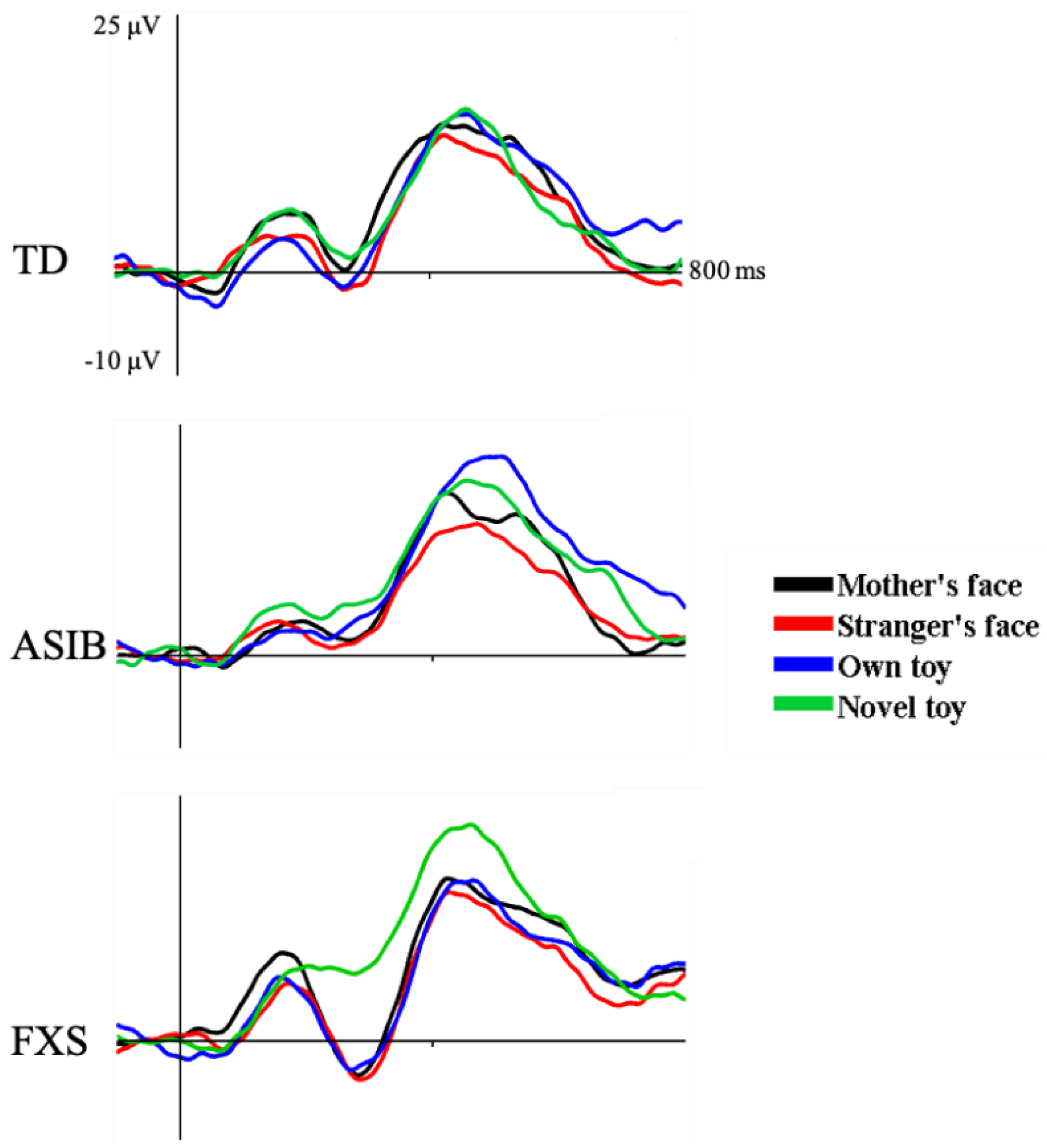
3.1. N290 ERP Component
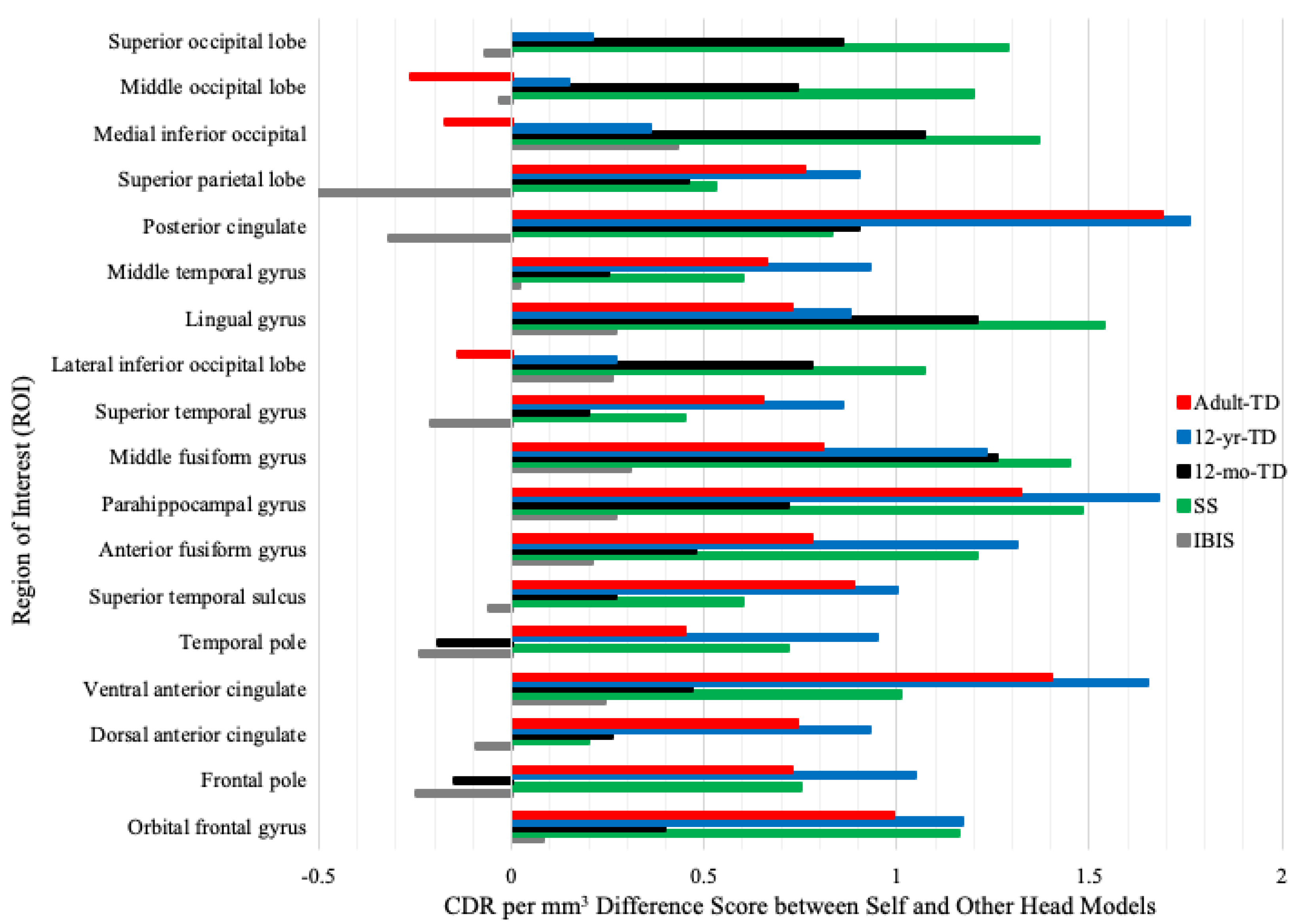
3.2. Head Model Comparison
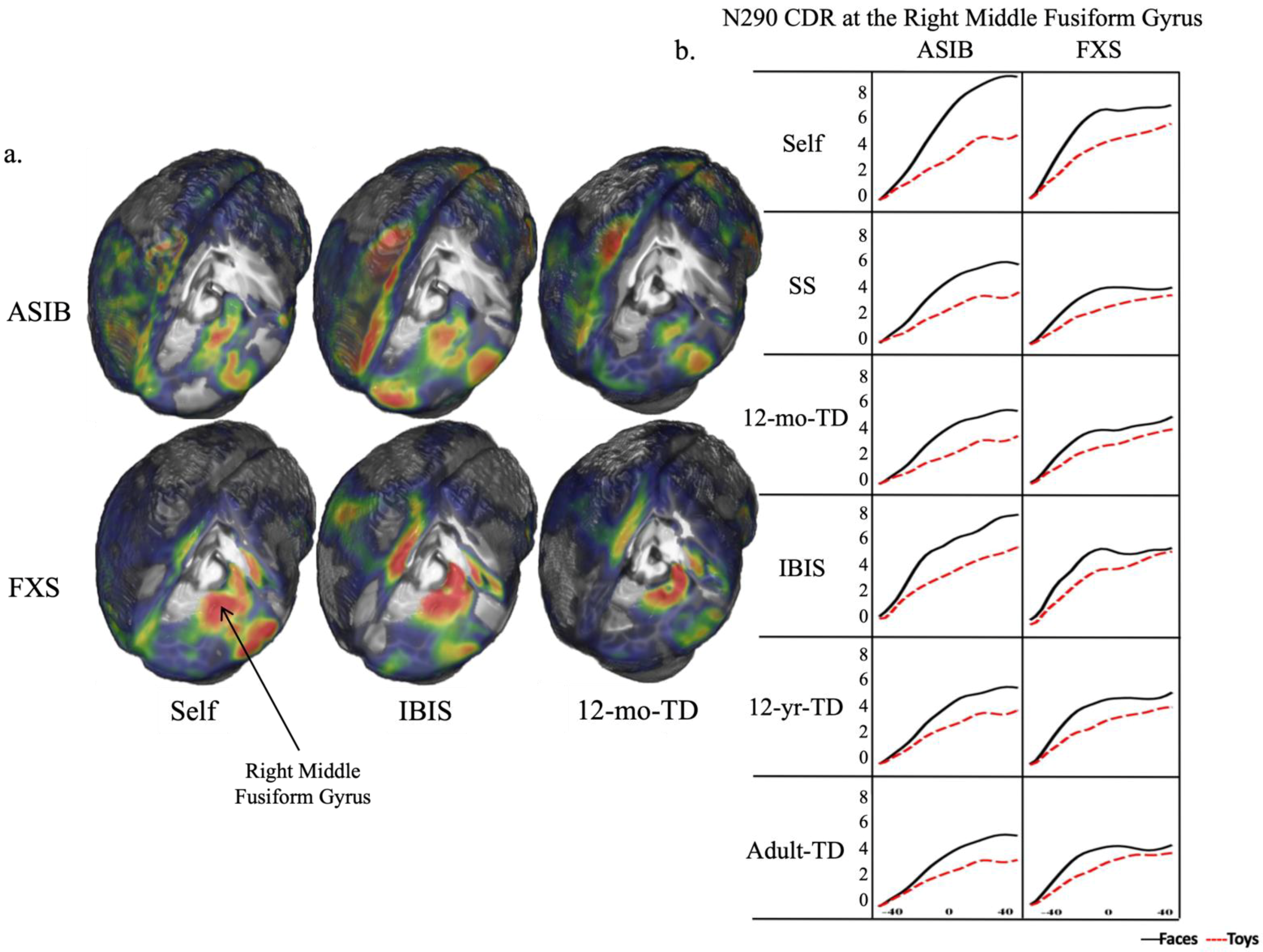
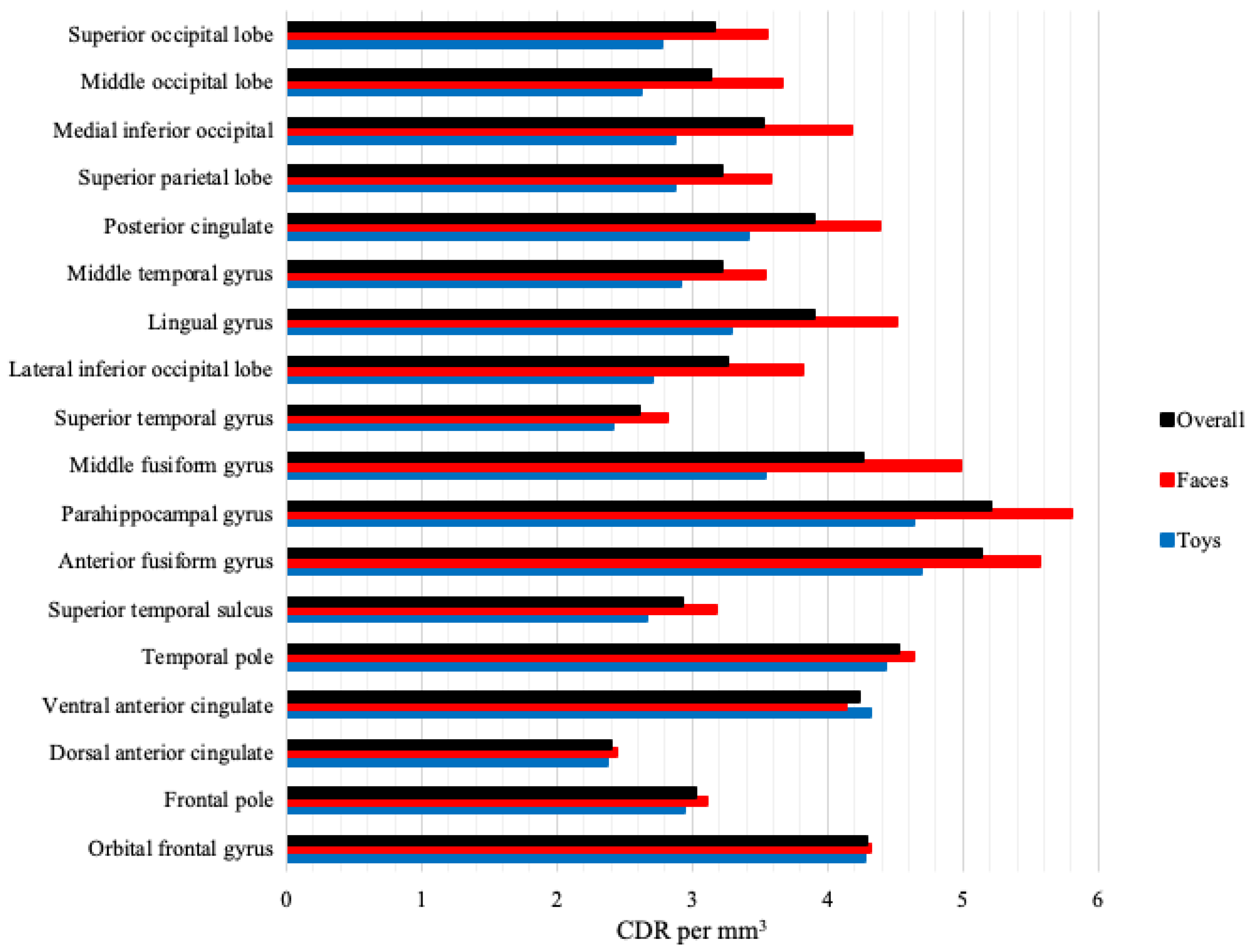
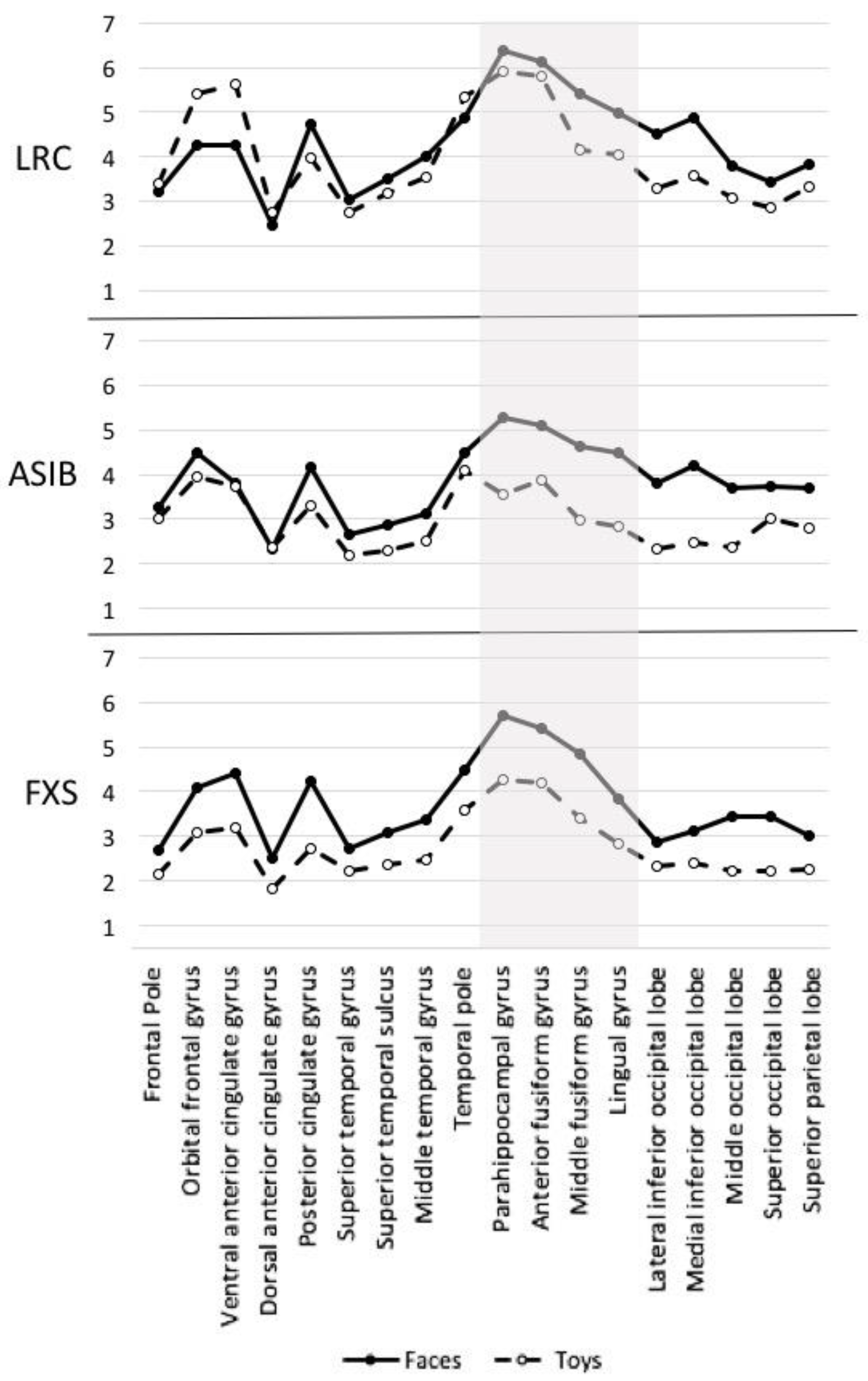
3.3. Cortical Source Analysis
4. Discussion
4.1. Head Model Comparison
4.2. Cortical Source Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ASD | autism spectrum disorder |
| CDR | current density reconstruction |
| ERP | event-related potential |
| FEM | finite element model |
| FXS | fragile X syndrome |
| IBIS | Infant Brain Imaging Study |
| ASIB | infant siblings of children with autism |
| LRC | low-risk control |
| MNI | Montreal Neurological Institute |
| Nc | Negative central |
| ROI | region of interest |
| SS | study-specific |
| TD | typically developing |
References
- Di Lorenzo, R.; Munsters, N.M.; Ward, E.K.; de Jonge, M.; Kemner, C.; van den Boomen, C. Is it fear? Similar brain responses to fearful and neutral faces in infants with a heightened likelihood for autism spectrum disorder. J. Autism Dev. Disord. 2021, 51, 961–972. [Google Scholar] [CrossRef] [PubMed]
- Elsabbagh, M.; Volein, A.; Csibra, G.; Holmboe, K.; Garwood, H.; Tucker, L.; Krljes, S.; Baron-Cohen, S.; Bolton, P.; Charman, T.; et al. Neural correlates of eye gaze processing in the infant broader autism phenotype. Biol. Psychiatry 2009, 65, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Elsabbagh, M.; Mercure, E.; Hudry, K.; Chandler, S.; Pasco, G.; Charman, T.; Pickles, A.; Baron-Cohen, S.; Bolton, P.; Johnson, M.H.; et al. Infant neural sensitivity to dynamic eye gaze is associated with later emerging autism. Curr. Biol. 2012, 22, 338–342. [Google Scholar] [CrossRef] [PubMed]
- Guy, M.W.; Richards, J.E.; Tonnsen, B.L.; Roberts, J.E. Neural correlates of face processing in etiologically-distinct 12-month-old infants at high-risk of autism spectrum disorder. Dev. Cogn. Neurosci. 2018, 29, 61–71. [Google Scholar] [CrossRef]
- Keehn, B.; Vogel-Farley, V.; Tager-Flusberg, H.; Nelson, C.A. Atypical hemispheric specialization for faces in infants at-risk for autism spectrum disorder. Autism Res. 2015, 8, 187–198. [Google Scholar] [CrossRef]
- McCleery, J.P.; Akshoomoff, N.; Dobkins, K.R.; Carver, L.J. Atypical face versus object processing and hemispheric asymmetries in 10-month-old infants at risk for autism. Biol. Psychiatry 2009, 66, 950–957. [Google Scholar] [CrossRef]
- Messinger, D.; Young, G.S.; Ozonoff, S.; Dobkins, K.; Carter, A.; Zwaigenbaum, L.; Landa, R.J.; Charman, T.; Stone, W.L.; Constantino, J.N.; et al. Beyond autism: A baby siblings research consortium study of high-risk children at three years of age. J. Am. Acad. Child Adolesc. Psychiatry 2013, 52, 300–308. [Google Scholar] [CrossRef]
- Ozonoff, S.; Young, G.S.; Carter, A.; Messinger, D.; Yirmiya, N.; Zwaigenbaum, L.; Bryson, S.; Carver, L.J.; Constantino, J.N.; Dobkins, K.; et al. Recurrence risk for autism spectrum disorders: A Baby Siblings Research Consortium study. Pediatrics 2011, 128, e488–e495. [Google Scholar] [CrossRef]
- Szatmari, P.; Chawarska, K.; Dawson, G.; Georgiades, S.; Landa, R.; Lord, C.; Messinger, D.S.; Thurm, A.; Halladay, A. Prospective longitudinal studies of infant siblings of children with autism: Lessons learned and future directions. J. Am. Acad. Child Adolesc. Psychiatry 2016, 55, 179–187. [Google Scholar] [CrossRef]
- Roberts, J.E.; Bradshaw, J.; Will, E.A.; Hogan, A.L.; McQuillen, S.; Hills, K. Emergence and rate of autism in fragile X syndrome across the first years of life. Dev. Psychopathol. 2020, 32, 1335–1352. [Google Scholar] [CrossRef]
- Shaw, K.A.; Maenner, M.J.; Baio, J.; Washington, A.; Christensen, D.L.; Wiggins, L.D.; Pettygrove, S.; Andrews, J.G.; White, T.; Robinson Rosenberg, C.; et al. Early identification of autism spectrum disorder among children aged 4 years—Early Autism and Developmental Disabilities Monitoring Network, six sites, United States 2020. MMWR Surveill. Summ. 2020, 69, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Conte, S.; Richards, J.E.; Guy, M.W.; Xie, W.; Roberts, J.E. Face-sensitive brain responses in the first year of life. NeuroImage 2020, 211, 116602. [Google Scholar] [CrossRef]
- Guy, M.W.; Zieber, N.; Richards, J.E. The cortical development of specialized face processing in infancy. Child Dev. 2016, 87, 1581–1600. [Google Scholar] [CrossRef] [PubMed]
- Halit, H.; de Haan, M.; Johnson, M.H. Cortical specialisation for face processing: Face-sensitive event-related potential components in 3- and 12-month-old infants. NeuroImage 2003, 19, 1180–1193. [Google Scholar] [CrossRef]
- Grelotti, D.J.; Gauthier, I.; Schultz, R.T. Social interest and the development of cortical face specialization: What autism teaches us about face processing. Dev. Psychobiol. 2002, 40, 213–225. [Google Scholar] [CrossRef]
- Hubl, D.; Bölte, S.; Feineis-Matthews, S.; Lanfermann, H.; Federspiel, A.; Strik, W.; Poustka, F.; Dierks, T. Functional imbalance of visual pathways indicates alternative face processing strategies in autism. Neurology 2003, 61, 1232–1237. [Google Scholar] [CrossRef]
- McPartland, J.; Dawson, G.; Webb, S.J.; Panagiotides, H.; Carver, L.J. Event-related brain potentials reveal anomalies in temporal processing of faces in autism spectrum disorder. J. Child Psychol. Psychiatry 2004, 45, 1235–1245. [Google Scholar] [CrossRef]
- Shephard, E.; Milosavljevic, B.; Mason, L.; Elsabbagh, M.; Tye, C.; Gliga, T.; Jones, E.J.H.; Charman, T.; Johnson, M.H.; the BASIS Team. Neural and behavioural indices of face processing in siblings of children with autism spectrum disorder (ASD): A longitudinal study from infancy to mid-childhood. Cortex 2020, 127, 162–179. [Google Scholar] [CrossRef]
- de Haan, M.; Johnson, M.H.; Halit, H. Development of face-sensitive event-related potentials during infancy: A review. Int. J. Psychophysiol. 2003, 51, 45–58. [Google Scholar] [CrossRef]
- Halit, H.; Csibra, G.; Volein, A.; Johnson, M.H. Face-sensitive cortical processing in early infancy. J. Child Psychol. Psychiatry 2004, 45, 1228–1234. [Google Scholar] [CrossRef]
- Hoehl, S.; Peykarjou, S. The early development of face processing—What makes faces special? Neurosci. Bull. 2012, 28, 765–788. [Google Scholar] [CrossRef] [PubMed]
- Key, A.P.F.; Stone, W.L. Processing of novel and familiar faces in infants at average and high risk for autism. Dev. Cogn. Neurosci. 2012, 2, 244–255. [Google Scholar] [CrossRef] [PubMed]
- Luyster, R.J.; Powell, C.; Tager-Flusberg, H.; Nelson, C.A. Neural measures of social attention across the first years of life: Characterizing typical development and markers of autism risk. Dev. Cogn. Neurosci. 2014, 8, 131–143. [Google Scholar] [CrossRef] [PubMed]
- de Haan, M.; Nelson, C.A. Recognition of the mother’s face by six-month-old infants: A neurobehavioral study. Child Dev. 1997, 68, 187–210. [Google Scholar] [CrossRef] [PubMed]
- de Haan, M.; Nelson, C.A. Brain activity differentiates face and object processing in 6-month-old infants. Dev. Psychol. 1999, 35, 1113–1121. [Google Scholar] [CrossRef]
- Luyster, R.J.; Wagner, J.B.; Vogel-Farley, V.; Tager-Flusberg, H.; Nelson, C.A. Neural correlates of familiar and unfamiliar face processing in infants at risk for autism spectrum disorders. Brain Topogr. 2011, 24, 220–228. [Google Scholar] [CrossRef]
- Côte, V.; Lalancette, È.; Knoth, I.S.; Côte, L.; Agbogba, K.; Vannasing, P.; Major, P.; Barlaam, F.; Michaud, J.; Lippé, S. Distinct patterns of repetition suppression in fragile X syndrome, down syndrome, tuberous sclerosis complex and mutations in SYNGAP1. Brain Res. 2021, 1751, 147205. [Google Scholar] [CrossRef]
- Fu, X.; Richards, J.E. Evaluating head models for cortical source localization of the face-sensitive N290 component in infants. Brain Topogr. 2022, 35, 398–415. [Google Scholar] [CrossRef]
- Johnson, M.H.; Griffin, R.; Csibra, G.; Halit, H.; Farroni, T.; de Haan, M.; Tucker, L.A.; Baron-Cohen, S.; Richards, J. The emergence of the social brain network: Evidence from typical and atypical development. Dev. Psychopathol. 2005, 17, 599–619. [Google Scholar] [CrossRef]
- Bentin, S.; Allison, T.; Puce, A.; Perez, E.; McCarthy, G. Electrophysiological studies of face perception in humans. J. Cognit. Neurosci. 1996, 8, 551–565. [Google Scholar] [CrossRef] [Green Version]
- Caldara, R.; Thut, G.; Servoir, P.; Michel, C.M.; Bovet, P.; Renault, B. Face versus non-face object perception and the ‘other-race’ effect: A spatio-temporal event-related potential study. Clin. Neurophysiol. 2003, 114, 515–528. [Google Scholar] [CrossRef]
- Carmel, S.; Bentin, S. Domain specificity versus expertise: Factors influencing distinct processing of faces. Cognition 2002, 83, 29. [Google Scholar] [CrossRef]
- Itier, R.J.; Latinus, M.; Taylor, M.J. Face, eye and object early processing: What is the face specificity? NeuroImage 2006, 29, 667–676. [Google Scholar] [CrossRef] [PubMed]
- Itier, R.J.; Taylor, M.J. Face recognition memory and configural processing: A developmental ERP study using upright, inverted, and contrast-reversed faces. J. Cognit. Neurosci. 2004, 16, 487–502. [Google Scholar] [CrossRef]
- Itier, R.J.; Taylor, M.J. Source analysis of the N170 to faces and objects. Neuroreport 2004, 15, 1261–1265. [Google Scholar] [CrossRef]
- Rossion, B.; Gauthier, I.; Tarr, M.J.; Despland, P.; Bruyer, R.; Linotte, S.; Crommelinck, M. The N170 occipito-temporal component is delayed and enhanced to inverted faces but not to inverted objects: An electrophysiological account of face-specific processes in the human brain. Neuroreport 2000, 11, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Gao, C.; Conte, S.; Richards, J.E.; Xie, W.; Hanayik, T. The neural sources of N170: Understanding timing of activation in face-selective areas. Psychophysiology 2019, 56, e13336. [Google Scholar] [CrossRef]
- Deffke, I.; Sander, T.; Heidenreich, J.; Sommer, W.; Curio, G.; Trahms, L.; Lueschow, A. MEG/EEG sources of the 170-ms response to faces are co-localized in the fusiform gyrus. NeuroImage 2007, 35, 1495–1501. [Google Scholar] [CrossRef]
- Rossion, B.; Joyce, C.A.; Cottrell, G.W.; Tarr, M.J. Early lateralization and orientation tuning for face, word, and object processing in the visual cortex. NeuroImage 2003, 20, 1609–1624. [Google Scholar] [CrossRef]
- Shibata, T.; Nishijo, H.; Tamura, R.; Miyamoto, K.; Eifuku, S.; Endo, S.; Ono, T. Generators of visual evoked potentials for faces and eyes in the human brain as determined by dipole localization. Brain Topogr. 2002, 15, 51–63. [Google Scholar] [CrossRef]
- Corrigan, N.M.; Richards, T.; Webb, S.J.; Murias, M.; Merkle, K.; Kleinhans, N.M.; Johnson, L.C.; Poliakov, A.; Aylward, E.; Dawson, G. An investigation of the relationship between fMRI and ERP source localized measurements of brain activity during face processing. Brain Topogr. 2009, 22, 83–96. [Google Scholar] [CrossRef] [PubMed]
- Horovitz, S.G.; Rossion, B.; Skudlarski, P.; Gore, J.C. Parametric design and correlational analyses help integrating fMRI and electrophysiological data during face processing. NeuroImage 2004, 22, 1587–1595. [Google Scholar] [CrossRef]
- Sadeh, B.; Podlipsky, I.; Zhdanov, A.; Yovel, G. Event-related potential and functional MRI measures of face-selectivity are highly correlated: A simultaneous ERP-fMRI investigation. Hum. Brain Mapp. 2010, 31, 1490–1501. [Google Scholar] [CrossRef]
- Garrett, A.S.; Menon, V.; MacKenzie, K.; Reiss, A.L. Here’s looking at you, kid: Neural systems underlying face and gaze processing in fragile X syndrome. Arch. Gen. Psychiatry 2004, 61, 281–288. [Google Scholar] [CrossRef]
- Reynolds, G.D.; Richards, J.E. Cortical Source Localization of Infant Cognition. Dev. Neuropsychol. 2009, 34, 312–329. [Google Scholar] [CrossRef]
- Mazziotta, J.C.; Toga, A.W.; Evans, A.; Fox, P.; Lancaster, J. A probablistic atlas of the human brain: Theory and rationale for its development. NeuroImage 1995, 2, 89–101. [Google Scholar] [CrossRef]
- Mazziotta, J.; Toga, A.; Evans, A.; Fox, P.; Lancaster, J.; Zilles, K.; Woods, R.; Paus, T.; Simpson, G.; Pike, B.; et al. A probabilistic atlas and reference system for the human brain: International Consortium for Brain Mapping (ICBM). Philos. Trans. R Soc. Lond. B-Biol. Sci. 2001, 356, 1293–1322. [Google Scholar] [CrossRef]
- Conte, S.; Richards, J.E. Cortical source analysis of event-related potentials: A developmental approach. Dev. Cogn. Neurosci. 2022, 54, 101092. [Google Scholar] [CrossRef]
- Richards, J.E.; Xie, W. Brains for all the ages: Structural neurodevelopment in infants and children from a life-span perspective. In Advances in Child Development and Behavior; Benson, J.B., Ed.; Elsevier: Amsterdam, The Netherlands, 2015; Volume 48, pp. 1–52. [Google Scholar] [CrossRef]
- Muzik, O.; Chugani, D.C.; Juhasz, C.; Shen, C.G.; Chugani, H.T. Statistical parametric mapping: Assessment of application in children. NeuroImage 2000, 12, 538–549. [Google Scholar] [CrossRef]
- Joshi, S.; Davis, B.; Jomier, M.; Gerig, G. Unbiased diffeomorphic atlas construction for computational anatomy. NeuroImage 2004, 23, S151–S160. [Google Scholar] [CrossRef]
- Prastawa, M.; Gilmore, J.H.; Lin, W.; Gerig, G. Automatic segmentation of MR images of the developing newborn brain. Med. Image Anal. 2005, 9, 457–466. [Google Scholar] [CrossRef]
- Sanchez, C.; Richards, J.; Almli, C.R. Neurodevelopmental MRI brain templates for children from 2 weeks to 4 years of age. Dev. Psychol. 2011, 54, 77–91. [Google Scholar] [CrossRef]
- Sanchez, C.; Richards, J.; Almli, C.R. Age-specific MRI templates for pediatric neuroimaging. Dev. Neuropsychol. 2012, 37, 379–399. [Google Scholar] [CrossRef]
- Wilke, M.; Schmithorst, V.J.; Holland, S.K. Assessment of spatial normalization of whole-brain magnetic resonance images in children. Hum. Brain Mapp. 2002, 17, 48–60. [Google Scholar] [CrossRef]
- Fillmore, P.T.; Richards, J.E.; Phillips-Meek, M.C.; Cryer, A.; Stevens, M. Stereotaxic magnetic resonance imaging brain atlases for infants from 3 to 12 months. Dev. Neurosci. 2015, 37, 515–532. [Google Scholar] [CrossRef]
- Fonov, V.; Evans, A.C.; Botteron, K.; Almli, C.R.; McKinstry, R.C.; Collins, D.L. Unbiased average age-appropriate atlases for pediatric studies. NeuroImage 2011, 54, 313–327. [Google Scholar] [CrossRef] [PubMed]
- Richards, J.E.; Boswell, C.; Stevens, M.; Vendemia, J.M.C. Evaluating methods for constructing average high-density electrode positions. Brain Topogr. 2015, 28, 70–86. [Google Scholar] [CrossRef]
- Courchesne, E.; Carper, R.; Akshoomoff, N. Evidence of brain overgrowth in the first year of life in autism. JAMA 2003, 290, 337–344. [Google Scholar] [CrossRef]
- Dementieva, Y.A.; Vance, D.D.; Donnelly, S.L.; Elston, L.A.; Wolpert, C.M.; Ravan, S.A.; DeLong, G.R.; Abramson, R.K.; Wright, H.H.; Cuccaro, M.L. Accelerated head growth in early development of individuals with autism. Pediatr. Neurol. 2005, 32, 102–108. [Google Scholar] [CrossRef]
- Dissanayake, C.; Bui, Q.M.; Huggins, R.; Loesch, D.Z. Growth in stature and head circumference in high-functioning autism and Asperger disorder during the first 3 years of life. Dev. Psychopathol. 2006, 18, 381–393. [Google Scholar] [CrossRef]
- Hazlett, H.C.; Poe, M.; Gerig, G.; Smith, R.G.; Provenzale, J.; Ross, A.; Gilmore, J.; Piven, J. Magnetic resonance imaging and head circumference study of brain size in autism: Birth through age 2 years. Arch. Gen. Psychiatry 2005, 62, 1366–1376. [Google Scholar] [CrossRef] [PubMed]
- Hazlett, H.; Gu, H.; Munsell, B.C.; Kim, S.H.; Styner, M.; Wolff, J.J.; Elison, J.T.; Swanson, M.R.; Zhu, H.; Botterton, K.N.; et al. Early brain development in infants at high risk for autism spectrum disorder. Nature 2017, 542, 348–353. [Google Scholar] [CrossRef] [PubMed]
- Hazlett, H.C.; Poe, M.D.; Lightbody, A.A.; Styner, M.; MacFall, J.R.; Reiss, A.L.; Piven, J. Trajectories of early brain volume development in fragile X syndrome and autism. J. Am. Acad. Child Adolesc. Psychiatry 2012, 51, 921–933. [Google Scholar] [CrossRef] [PubMed]
- Hallez, H.; Vanrumste, B.; Grech, R.; Muscat, J.; De Clercq, W.; Vergult, A.; D’Asseler, Y.; Camilleri, K.P.; Fabri, S.G.; Van Huffel, S.; et al. Review on solving the forward problem in EEG source analysis. J. Neuroeng. Rehabil. 2007, 4, 46. [Google Scholar] [CrossRef] [PubMed]
- Michel, C.M.; Murray, M.M.; Lantz, G.; Gonzalez, S.; Spinelli, L.; de Peralta, R.G. EEG source imaging. Clin. Neurophysiol. 2004, 115, 2195–2222. [Google Scholar] [CrossRef]
- Jenkinson, M.; Pechaud, M.; Smith, S. BET2: MR-based estimation of brain, skull and scalp surfaces. In Proceedings of the Eleventh Annual Meeting of the Organization for Human Brain Mapping, Toronto, ON, Canada, 12–16 June 2005; p. 716. [Google Scholar]
- Conte, S.; Richards, J.E. The influence of head model conductor on the source localization of auditory evoked potentials. Brain Topogr. 2021, 34, 793–812. [Google Scholar] [CrossRef] [PubMed]
- Jenkinson, M.; Smith, S. A global optimisation method for robust affine registration of brain images. Med. Image Anal. 2001, 5, 143–156. [Google Scholar] [CrossRef]
- Avants, B.B.; Epstein, C.L.; Grossman, M.; Gee, J.C. Symmetric diffeomorphic image registration with cross-correlation: Evaluating automated labeling of elderly and neurodegenerative brain. Med. Image Anal. 2008, 12, 26–41. [Google Scholar] [CrossRef]
- Avants, B.B.; Tustison, N.J.; Song, G.; Cook, P.A.; Klein, A.; Gee, J.C. A reproducible evaluation of ANTs similarity metric performance in brain image registration. NeuroImage 2011, 54, 2033–2044. [Google Scholar] [CrossRef] [Green Version]
- Guy, M.W.; Conte, S.; Bursalioglu, A.; Richards, J.E. Peak selection and latency jitter correction in developmental event-related potentials. Dev. Psychobiol. 2021, 63, e22193. [Google Scholar] [CrossRef]
- Darvas, F.; Schmitt, U.; Louis, A.K.; Fuchs, M.; Knoll, G.; Buchner, H. Spatiotemporal current density reconstruction (stCDR) from EEG/MET data. Brain Topogr. 2001, 13, 195–207. [Google Scholar] [CrossRef] [PubMed]
- Pascual-Marqui, R.D.; Lehmann, D.; Koukkou, M.; Kochi, K.; Anderer, P.; Saletu, B.; Kinoshita, T. Assessing interactions in the brain with exact low-resolution electromagnetic tomography. Phil. Trans. Math. Phys. Eng. Sci. 2011, 369, 3768–3784. [Google Scholar] [CrossRef] [PubMed]
- Hazlett, H.C.; Poe, M.D.; Lightbody, A.A.; Gerig, G.; MacFall, J.R.; Ross, A.K.; Provenzale, J.; Martin, A.; Reiss, A.L.; Piven, J. Teasing apart the heterogeneity of autism: Same behavior, different brains in toddlers with fragile X syndrome and autism. J. Neurodev. Disord. 2009, 1, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Hoeft, F.; Lightbody, A.A.; Hazlett, H.C.; Patnaik, S.; Piven, J.; Reiss, A.L. Morphometric spatial patterns differentiating boys with fragile X syndrome, typically developing boys, and developmentally delayed boys aged 1 to 3 years. JAMA Psychiatry 2008, 65, 1087–1097. [Google Scholar] [CrossRef]
- Hoeft, F.; Walter, E.; Lightbody, A.A.; Hazlett, H.C.; Chang, C.; Piven, J.; Reiss, A.L. Neuroanatomical differences in toddler boys with fragile X syndrome and idiopathic autism. JAMA Psychiatry 2011, 68, 295–305. [Google Scholar] [CrossRef]
- Chen, H.; Uddin, L.Q.; Guo, X.; Wang, J.; Wang, R.; Wang, X.; Duan, X.; Chen, H. Parsing brain structural heterogeneity in males with autism spectrum disorder reveals distinct clinical subtypes. Hum. Brain Mapp. 2019, 40, 628–637. [Google Scholar] [CrossRef]
- Martinez-Murcia, F.J.; Lai, M.-C.; Gorriz, J.M.; Ramirez, J.; Young, A.M.H.; Deoni, S.C.L.; Ecker, C.; Lombardo, M.V.; MRC AIMS Consortium; Baron-Cohen, S.; et al. On the brain structure heterogeneity of autism: Parsing out acquisition side effects with significance-weighted principal component analysis. Hum. Brain Mapp. 2016, 38, 1208–1223. [Google Scholar] [CrossRef]
- Zabihi, M.; Oldehinkel, M.; Wolfers, T.; Frouin, V.; Goyard, D.; Loth, E.; Charman, T.; Tillmann, J.; Banaschewski, T.; Dumas, G.; et al. Dissecting the heterogeneous cortical anatomy of autism spectrum disorder using normative models. Biol. Psychiatry 2019, 4, 567–578. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Segment | Conductivity (S/m) |
|---|---|
| Scalp | 0.3500 |
| Skull | 0.0132 |
| Cerebrospinal fluid | 1.7900 |
| White matter | 0.2000 |
| Gray matter | 0.3300 |
| Dura | 0.3300 |
| Muscles | 0.3500 |
| Eyes | 0.5000 |
| Nasal cavity | 0.0048 |
| CDR per mm3 | ||||
|---|---|---|---|---|
| Head Model | Description | Mean | Faces | Toys |
| Self | Created from the participant’s own MRI | 3.39 | 3.97 | 2.80 |
| IBIS-ASIB/IBIS-FXS | Created from 53 (ASIBs) or 24 (FXS) MRIs collected as part of the IBIS study | −0.05 | −0.08 | −0.01 |
| SS-ASIB/SS-FXS | Created from 8 (ASIBs) or 11 (FXS) MRIs collected as part of the current study | −1.02 | −0.63 | −0.81 |
| 12-mo-TD | Created from 169 MRIs collected for the Neurodevelopmental MRI Database | −0.59 | −0.76 | −0.42 |
| 12-yr-TD | Created from 139 MRIs collected for the Neurodevelopmental MRI Database | −0.91 | −1.12 | −0.68 |
| Adult-TD | Created from 632 MRIs collected for the Neurodevelopmental MRI Database | −0.60 | −0.77 | −0.41 |
| ASIB | FXS | ||||||
|---|---|---|---|---|---|---|---|
| Head Model | Head Model | ||||||
| ROI | Self | SS | IBIS | ROI | Self | SS | IBIS |
| Parahippocampal gyrus | 4.65 | 3.53 | 4.54 | Parahippocampal gyrus | 5.19 | 3.45 | 4.80 |
| Anterior fusiform | 4.42 | 3.77 | 4.49 | Anterior fusiform | 4.99 | 3.37 | 4.57 |
| Lingual gyrus | 4.35 | 2.56 | 4.24 | Middle fusiform | 4.33 | 2.81 | 3.74 |
| Middle fusiform | 4.08 | 2.71 | 4.14 | Temporal pole | 4.24 | 3.00 | 4.46 |
| Posterior cingulate | 3.78 | 2.80 | 4.13 | Ventral anterior cingulate | 3.94 | 2.51 | 3.45 |
| Medial inferior occipital | 3.63 | 2.11 | 3.46 | Orbital frontal gyrus | 3.88 | 2.40 | 3.61 |
| Superior occipital lobe | 3.53 | 2.44 | 3.88 | Lingual gyrus | 3.63 | 2.27 | 3.24 |
| Temporal pole | 3.50 | 3.50 | 3.78 | Posterior cingulate | 3.46 | 2.75 | 3.76 |
| Orbital frontal gyrus | 3.26 | 2.55 | 3.44 | Medial inferior occipital | 3.03 | 1.77 | 2.41 |
| Superior parietal lobe | 3.20 | 2.94 | 3.91 | Middle occipital lobe | 2.97 | 1.52 | 2.69 |
| Lateral inferior occipital | 3.05 | 2.02 | 3.05 | Middle temporal gyrus | 2.82 | 2.08 | 2.75 |
| Middle occipital lobe | 3.05 | 2.18 | 3.50 | Superior occipital lobe | 2.82 | 1.39 | 2.70 |
| Ventral anterior cingulate | 3.03 | 2.59 | 3.13 | Lateral inferior occipital | 2.80 | 1.70 | 2.35 |
| Middle temporal gyrus | 2.65 | 2.25 | 2.70 | Superior temporal sulcus | 2.60 | 1.94 | 2.68 |
| Superior temporal sulcus | 2.53 | 2.02 | 2.57 | Frontal pole | 2.57 | 1.46 | 2.61 |
| Superior temporal gyrus | 2.53 | 2.02 | 2.57 | Superior parietal lobe | 2.53 | 1.79 | 2.89 |
| Frontal pole | 3.50 | 3.50 | 3.78 | Superior temporal gyrus | 2.37 | 1.85 | 2.61 |
| Dorsal anterior cingulate | 3.03 | 2.59 | 3.13 | Dorsal anterior cingulate | 2.08 | 1.93 | 2.08 |
| Correlation with Self Head Model (SD) | |||
|---|---|---|---|
| Head Model | Overall | ASIB | FXS |
| IBIS-ASIB/IBIS-FXS | .84 (.12) | .88 (.07) | .81 (.14) |
| SS-ASIB/SS-FXS | .81 (.13) | .81 (.17) | .81 (.10) |
| 12-mo-TD | .81 (.15) | .78 (.18) | .82 (.12) |
| 12-yr-TD | .70 (.12) | .77 (.12) | .66 (.26) |
| Adult-TD | .74 (.18) | .78 (.12) | .72 (.22) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guy, M.W.; Richards, J.E.; Roberts, J.E. Cortical Source Analysis of the Face Sensitive N290 ERP Component in Infants at High Risk for Autism. Brain Sci. 2022, 12, 1129. https://doi.org/10.3390/brainsci12091129
Guy MW, Richards JE, Roberts JE. Cortical Source Analysis of the Face Sensitive N290 ERP Component in Infants at High Risk for Autism. Brain Sciences. 2022; 12(9):1129. https://doi.org/10.3390/brainsci12091129
Chicago/Turabian StyleGuy, Maggie W., John E. Richards, and Jane E. Roberts. 2022. "Cortical Source Analysis of the Face Sensitive N290 ERP Component in Infants at High Risk for Autism" Brain Sciences 12, no. 9: 1129. https://doi.org/10.3390/brainsci12091129