
Adaptive Cognitive Control in Prematurely Born Children: An HD-EEG Investigation

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Experimental Procedures
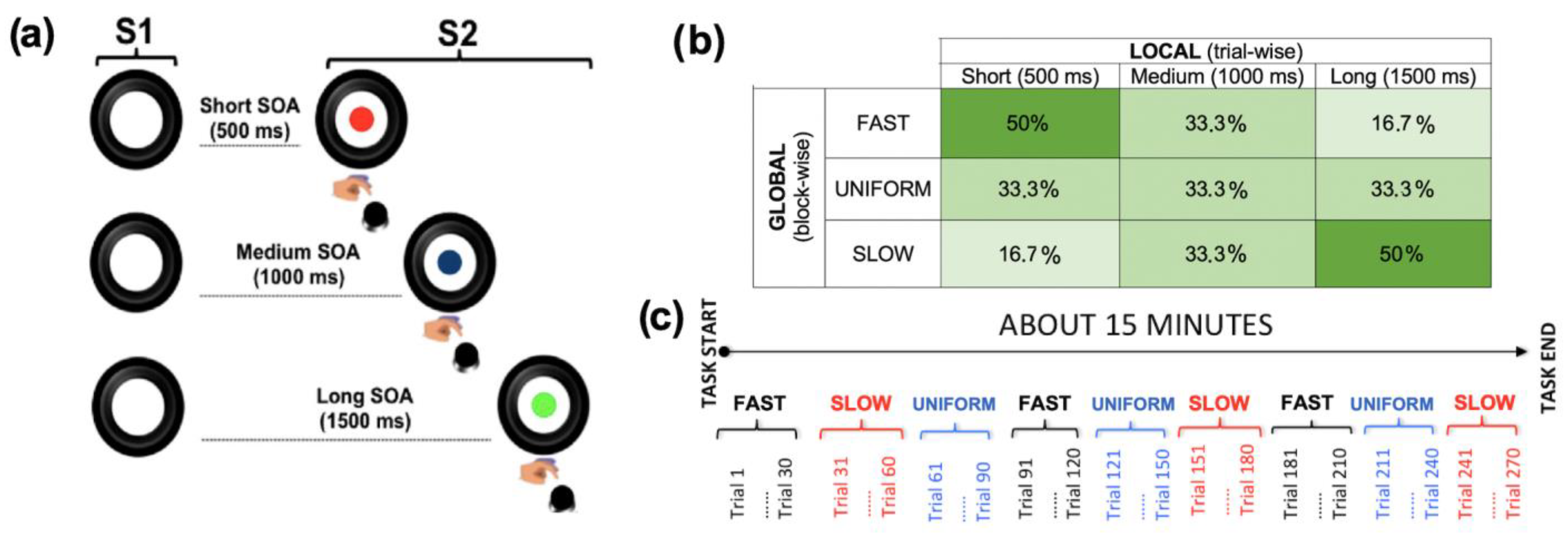
2.3. Experimental Task
2.3.1. Local Predictive Context
2.3.2. Global Predictive Context
Uniform (U) Blocks
Fast Blocks (Short-Biased)
Slow Blocks (Long-Biased)
2.4. Experimental Design
2.5. Behavioural Data Analysis
2.6. EEG Recording and Preprocessing
2.7. EEG Statistical Analysis
2.8. Cortical Source Modelling
3. Results
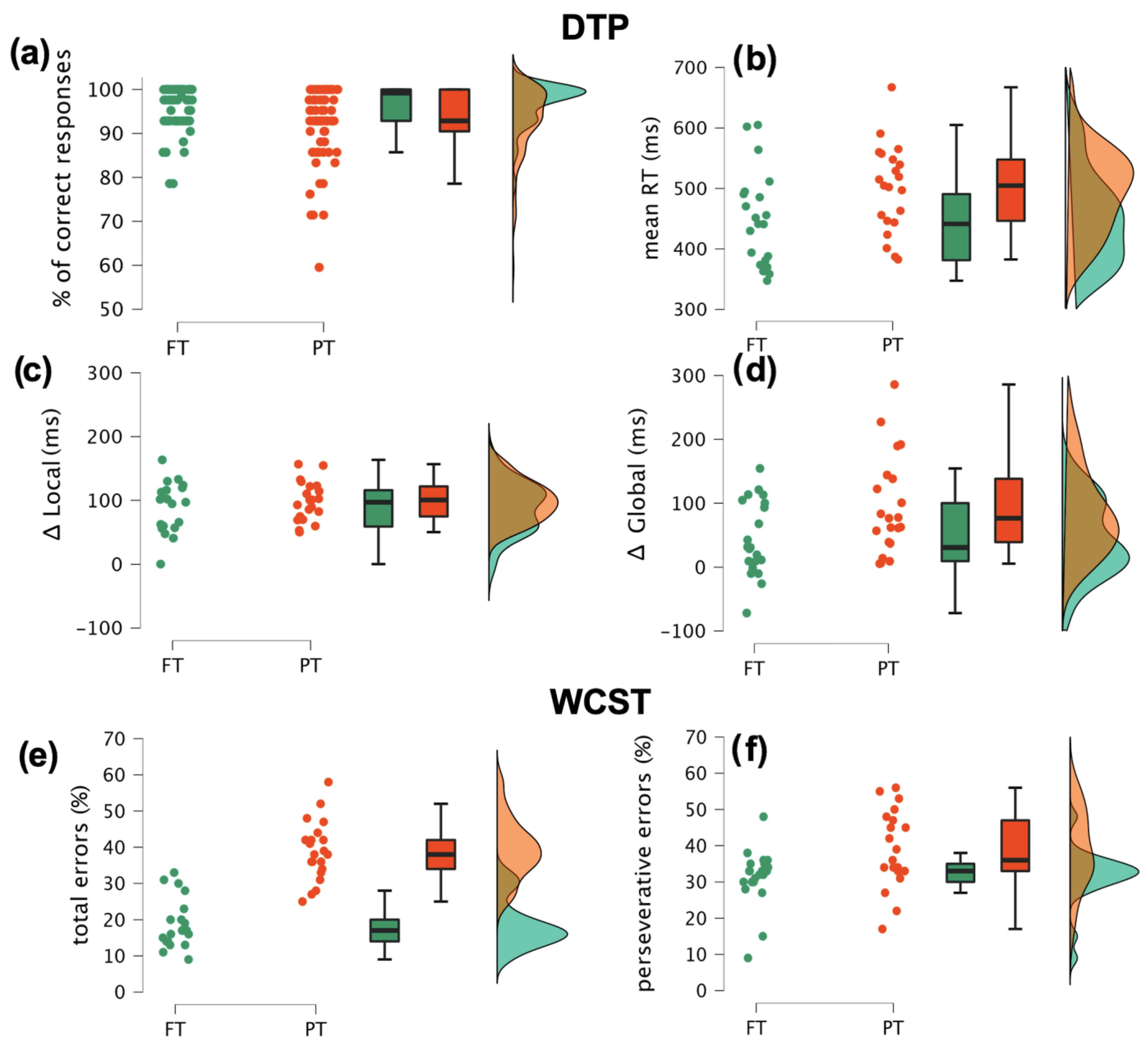
3.1. Behavioural Results
3.1.1. Dynamic Temporal Prediction Task
3.1.2. Wisconsin Card Sorting Test
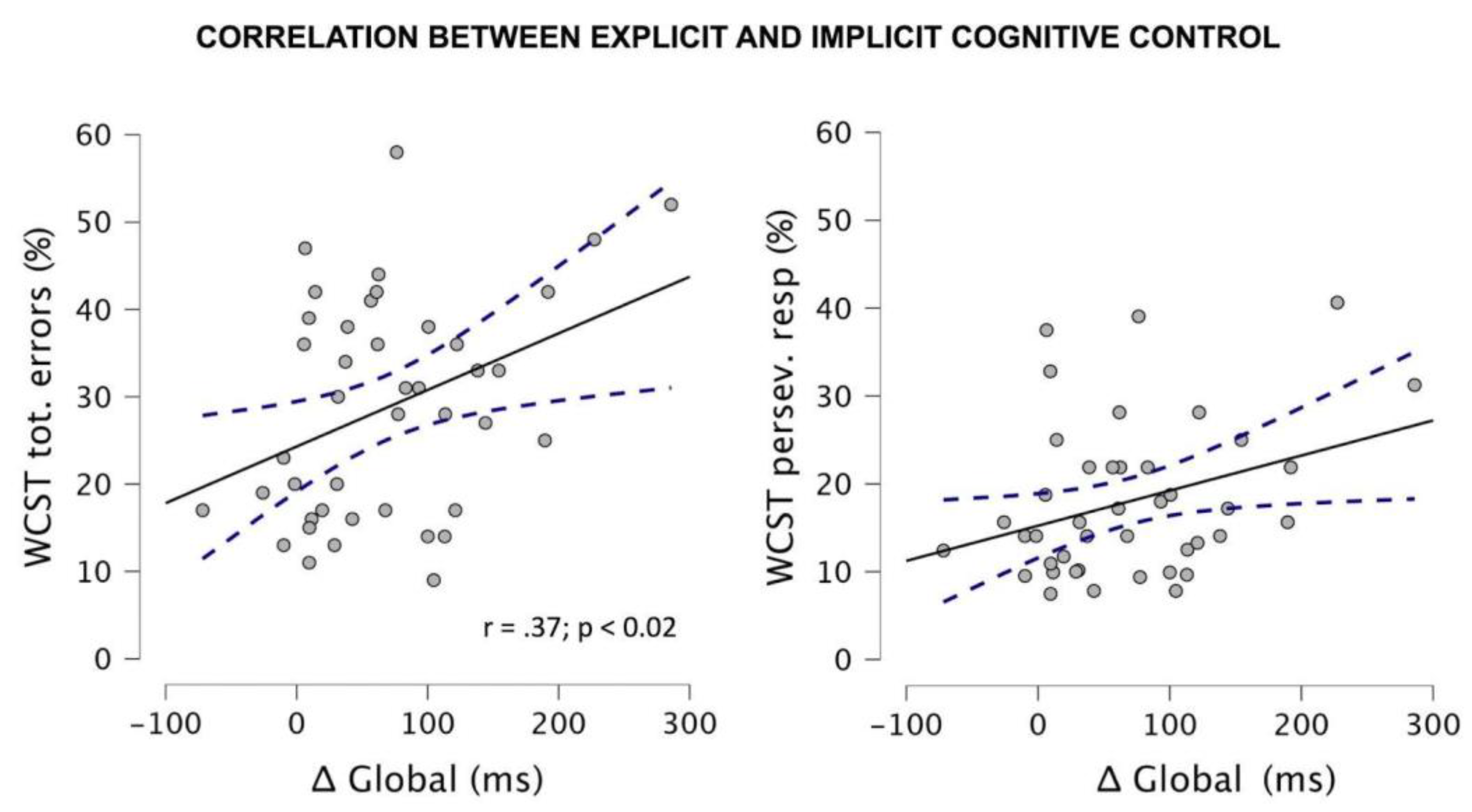
3.1.3. Correlational Analyses
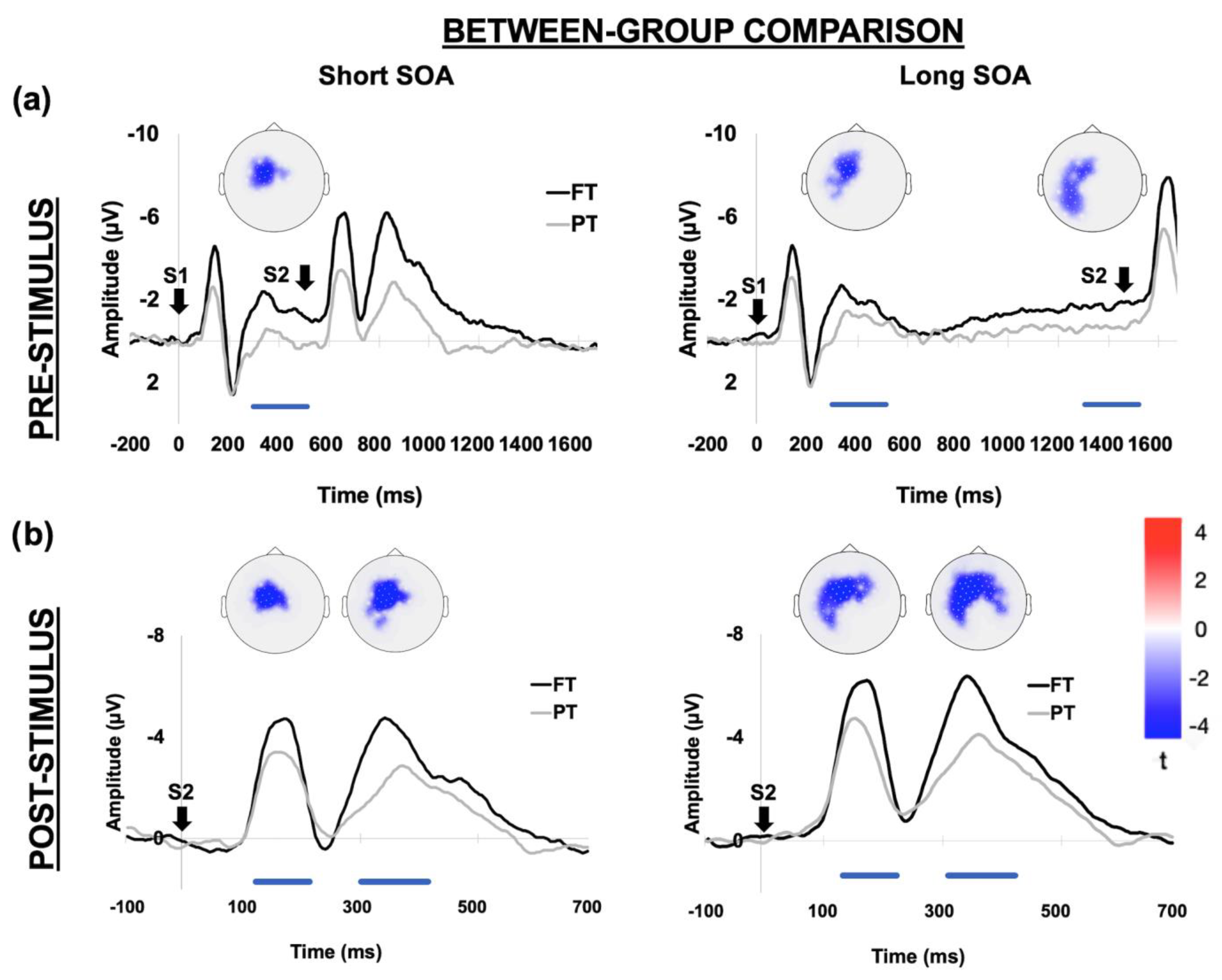
3.2. EEG Results
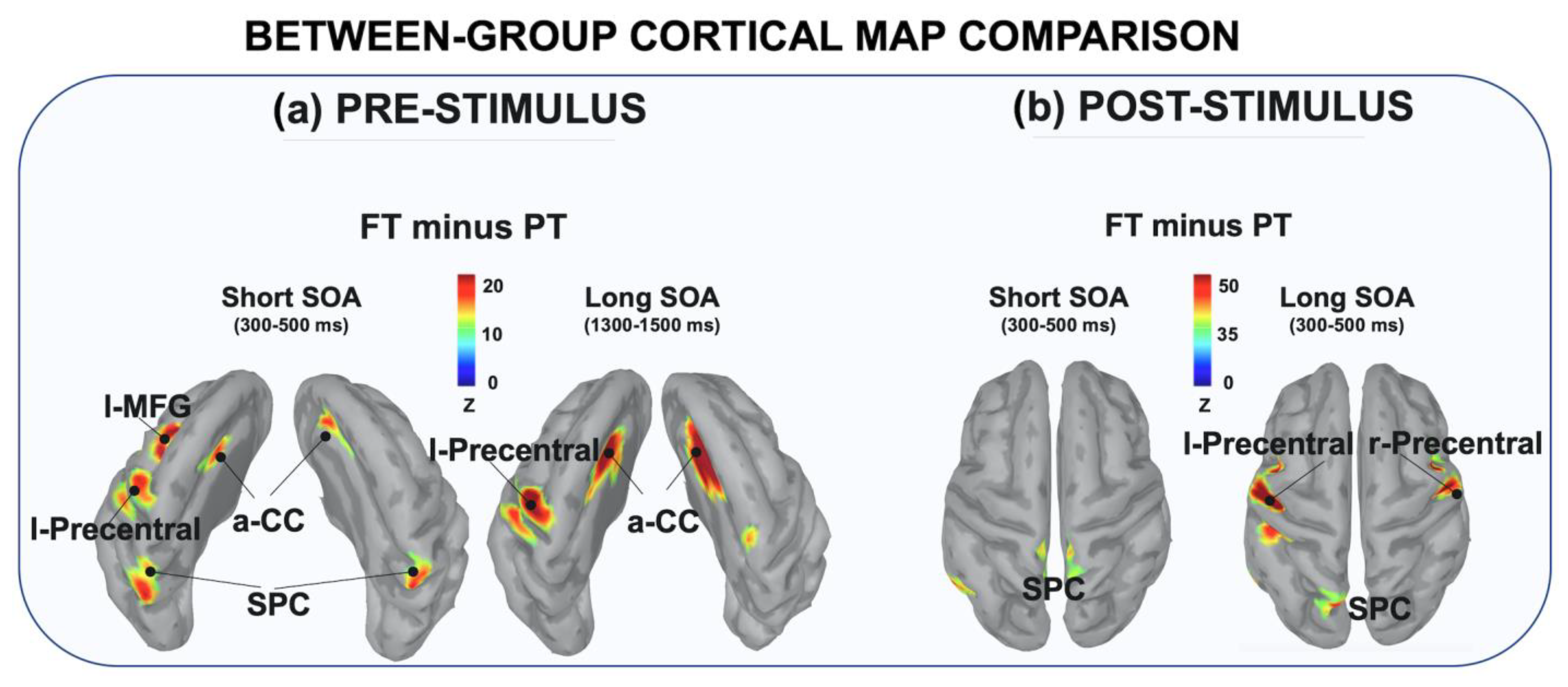
3.2.1. Between-Group Comparison
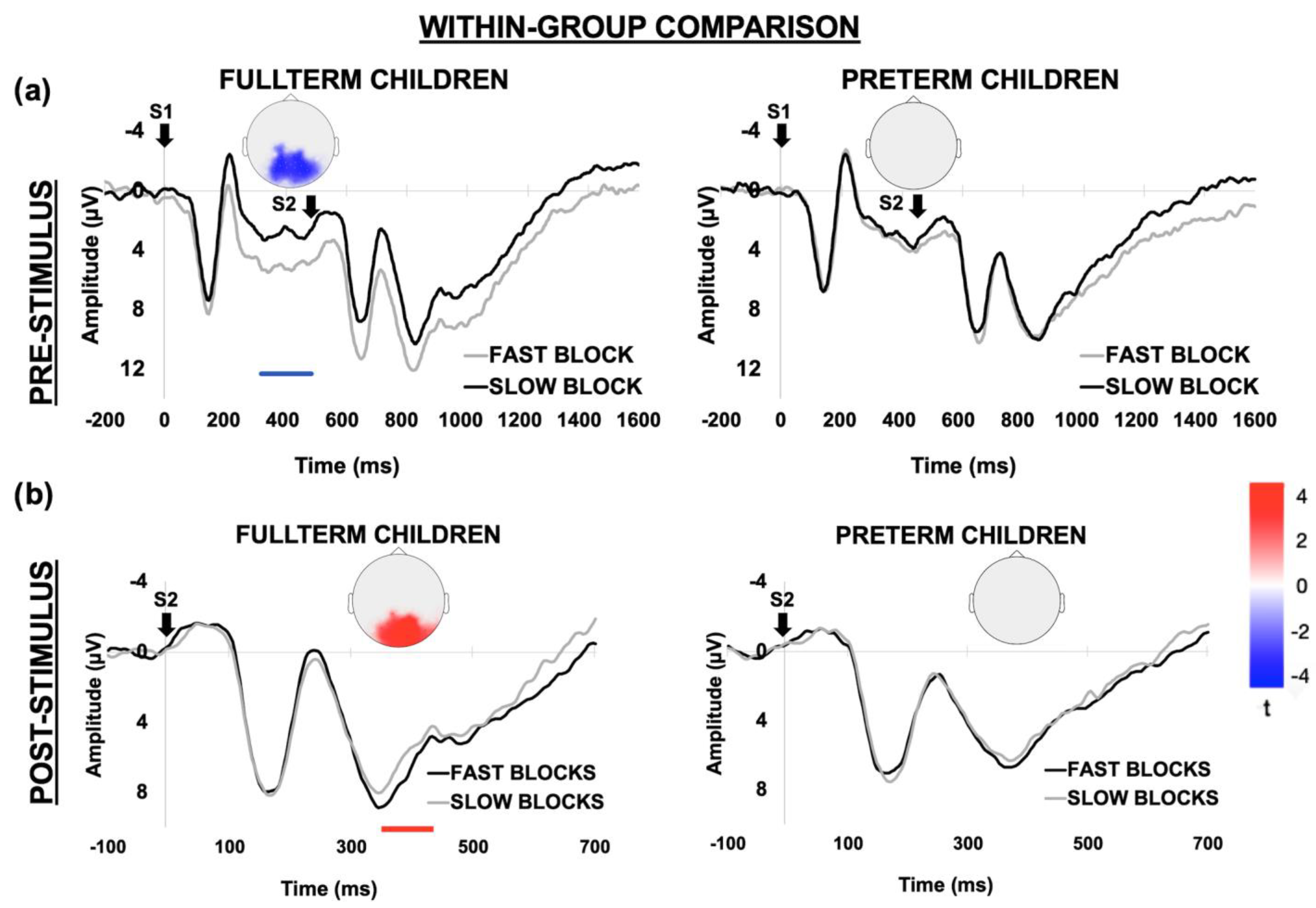
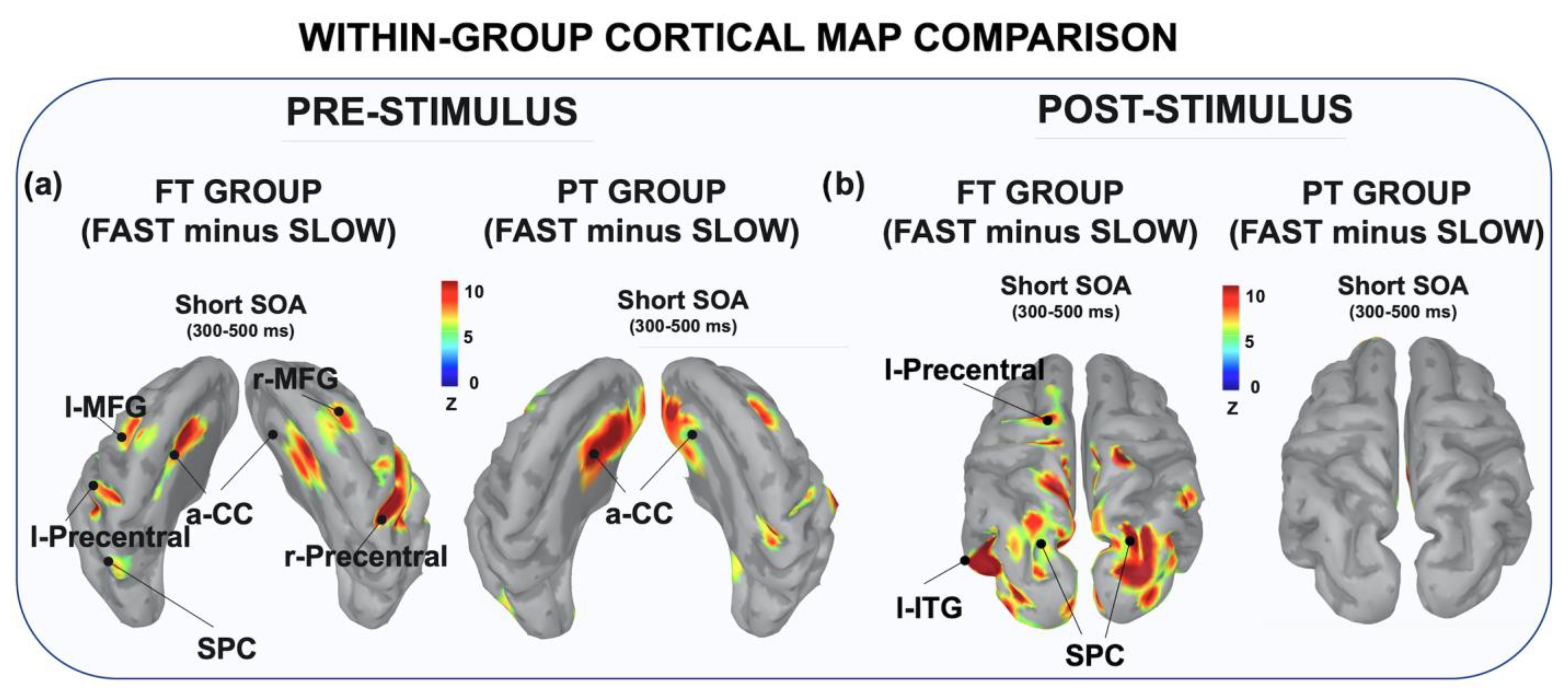
3.2.2. Within-Group Comparison
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aarnoudse-Moens, C.S.H.; Weisglas-Kuperus, N.; van Goudoever, J.B.; Oosterlaan, J. Meta-Analysis of Neurobehavioral Outcomes in Very Preterm and/or Very Low Birth Weight Children. Pediatrics 2009, 124, 717–728. [Google Scholar] [CrossRef]
- Tosello, B.; Méziane, S.; Resseguier, N.; Marret, S.; Cambonie, G.; Zahed, M.; Brévaut-Malaty, V.; Beltran Anzola, A.; Gire, C. The Neurobehavioral Phenotype of School-Aged, Very Prematurely Born Children with No Serious Neurological Sequelae: A Quality of Life Predictor. Children 2021, 8, 943. [Google Scholar] [CrossRef] [PubMed]
- Gire, C.; The GPQoL Study Group; Tosello, B.; Marret, S.; Cambonie, G.; Souksi-Medioni, I.; Müller, J.-B.; Garcia, P.; Berbis, J.; Auquier, P.; et al. Specific cognitive correlates of the quality of life of extremely preterm school-aged children without major neurodevelopmental disability. Pediatr. Res. 2020, 88, 642–652. [Google Scholar] [CrossRef] [PubMed]
- Mento, G.; Bisiacchi, P.S. Neurocognitive development in preterm infants: Insights from different approaches. Neurosci. Biobehav. Rev. 2012, 36, 536–555. [Google Scholar] [CrossRef]
- Mento, G.; Nosarti, C. The case of late preterm birth: Sliding forwards the critical window for cognitive outcome risk. Transl. Pediatr. 2015, 4, 214–218. [Google Scholar] [CrossRef] [PubMed]
- Nosarti, C. Structural and functional brain correlates of behavioral outcomes during adolescence. Early Hum. Dev. 2013, 89, 221–227. [Google Scholar] [CrossRef]
- White, A.; McKee, M.; De Sousa, B.; De Visser, R.; Hogston, R.; Madsen, S.A.; Makara, P.; Richardson, N.; Zatoński, W.; Raine, G. An examination of the association between premature mortality and life expectancy among men in Europe. Eur. J. Public Health 2014, 24, 673–679. [Google Scholar] [CrossRef]
- van Houdt, C.A.; Oosterlaan, J.; van Wassenaer-Leemhuis, A.G.; van Kaam, A.H.; Aarnoudse-Moens, C.S.H. Executive function deficits in children born preterm or at low birthweight: A meta-analysis. Dev. Med. Child Neurol. 2019, 61, 1015–1024. [Google Scholar] [CrossRef]
- Cutini, S.; Duma, G.M.; Mento, G. How time shapes cognitive control: A high-density EEG study of task-switching. Biol. Psychol. 2021, 160, 108030. [Google Scholar] [CrossRef]
- Diamond, A. Executive Functions. Annu. Rev. Psychol. 2013, 64, 135–168. [Google Scholar] [CrossRef]
- Friedman, N.P.; Miyake, A. Unity and diversity of executive functions: Individual differences as a window on cognitive structure. Cortex 2017, 86, 186–204. [Google Scholar] [CrossRef] [PubMed]
- Miyake, A.; Friedman, N.P.; Emerson, M.J.; Witzki, A.H.; Howerter, A.; Wager, T.D. The Unity and Diversity of Executive Functions and Their Contributions to Complex “Frontal Lobe” Tasks: A Latent Variable Analysis. Cogn. Psychol. 2000, 41, 49–100. [Google Scholar] [CrossRef] [PubMed]
- Miller, E.K.; Cohen, J.D. An Integrative Theory of Prefrontal Cortex Function. Annu. Rev. Neurosci. 2001, 24, 167–202. [Google Scholar] [CrossRef] [PubMed]
- Norman, D.A.; Shallice, T. Attention to action. In Consciousness and Self-Regulation; Springer: Berlin/Heidelberg, Germany, 1986; pp. 1–18. [Google Scholar]
- Braem, S.; Verguts, T.; Notebaert, W. Conflict adaptation by means of associative learning. J. Exp. Psychol. Hum. Percept. Perform. 2011, 37, 1662–1666. [Google Scholar] [CrossRef]
- Braem, S.; Egner, T. Getting a Grip on Cognitive Flexibility. Curr. Dir. Psychol. Sci. 2018, 27, 470–476. [Google Scholar] [CrossRef]
- Saffran, J.R.; Aslin, R.N.; Newport, E.L. Statistical Learning by 8-Month-Old Infants. Science 1996, 274, 1926–1928. [Google Scholar] [CrossRef] [PubMed]
- Saffran, J.R.; Kirkham, N.Z. Infant Statistical Learning. Annu. Rev. Psychol. 2018, 69, 181–203. [Google Scholar] [CrossRef]
- Braver, T.S. The variable nature of cognitive control: A dual mechanisms framework. Trends Cogn. Sci. 2012, 16, 106–113. [Google Scholar] [CrossRef]
- Alink, A.; Schwiedrzik, C.M.; Kohler, A.; Singer, W.; Muckli, L. Stimulus Predictability Reduces Responses in Primary Visual Cortex. J. Neurosci. 2010, 30, 2960–2966. [Google Scholar] [CrossRef]
- Stefanics, G.; Heinzle, J.; Horváth, A.A.; Stephan, K.E. Visual Mismatch and Predictive Coding: A Computational Single-Trial ERP Study. J. Neurosci. 2018, 38, 4020–4030. [Google Scholar] [CrossRef]
- Baldeweg, T. Repetition effects to sounds: Evidence for predictive coding in the auditory system. Trends Cogn. Sci. 2006, 10, 93–94. [Google Scholar] [CrossRef]
- Duma, G.; Granziol, U.; Mento, G. Should I stay or should I go? How local-global implicit temporal expectancy shapes proactive motor control: An hdEEG study. NeuroImage 2020, 220, 117071. [Google Scholar] [CrossRef]
- Mento, G.; Tarantino, V.; Vallesi, A.; Bisiacchi, P.S. Spatiotemporal Neurodynamics Underlying Internally and Externally Driven Temporal Prediction: A High Spatial Resolution ERP Study. J. Cogn. Neurosci. 2015, 27, 425–439. [Google Scholar] [CrossRef]
- Mento, G.; Vallesi, A. Spatiotemporally dissociable neural signatures for generating and updating expectation over time in children: A High Density-ERP study. Dev. Cogn. Neurosci. 2016, 19, 98–106. [Google Scholar] [CrossRef]
- Barron, H.C.; Auksztulewicz, R.; Friston, K. Prediction and memory: A predictive coding account. Prog. Neurobiol. 2020, 192, 101821. [Google Scholar] [CrossRef]
- Barrett, L.F. The theory of constructed emotion: An active inference account of interoception and categorization. Soc. Cogn. Affect. Neurosci. 2017, 12, 1–23. [Google Scholar] [CrossRef]
- Cristaldi, F.D.P.; Mento, G.; Sarlo, M.; Buodo, G. Dealing with uncertainty: A high-density EEG investigation on how intolerance of uncertainty affects emotional predictions. PLoS ONE 2021, 16, e0254045. [Google Scholar] [CrossRef]
- Del Popolo Cristaldi, F.; Buodo, G.; Duma, G.M.; Sarlo, M.; Mento, G. Unbalanced functional connectivity at rest affects the ERP correlates of affective prediction in high intolerance of uncertainty individuals: A high density EEG investigation. Int. J. Psychophysiol. 2022, 178, 22–33. [Google Scholar] [CrossRef]
- Seth, A.K.; Friston, K.J. Active interoceptive inference and the emotional brain. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20160007. [Google Scholar] [CrossRef]
- Correa, Á. Enhancing Behavioural Performance by Visual Temporal Orienting. Attention and Time; Oxford University Press: Oxford, UK, 2010; pp. 357–370. [Google Scholar]
- Pezzulo, G. An Active Inference view of cognitive control. Front. Psychol. 2012, 3, 478. [Google Scholar] [CrossRef]
- Dixon, M.L.; Zelazo, P.D.; De Rosa, E. Evidence for intact memory-guided attention in school-aged children. Dev. Sci. 2010, 13, 161–169. [Google Scholar] [CrossRef]
- Nussenbaum, K.; Scerif, G.; Nobre, A.C. Differential Effects of Salient Visual Events on Memory—Guided Attention in Adults and Children. Child Dev. 2019, 90, 1369–1388. [Google Scholar] [CrossRef]
- Gonthier, C.; Blaye, A. Preschoolers can be instructed to use proactive control. Cogn. Dev. 2022, 62, 101175. [Google Scholar] [CrossRef]
- Chevalier, N.; Martis, S.B.; Curran, T.; Munakata, Y. Metacognitive Processes in Executive Control Development: The Case of Reactive and Proactive Control. J. Cogn. Neurosci. 2015, 27, 1125–1136. [Google Scholar] [CrossRef]
- Chevalier, N.; Blaye, A. Metacognitive Monitoring of Executive Control Engagement during Childhood. Child Dev. 2016, 87, 1264–1276. [Google Scholar] [CrossRef]
- Niebaum, J.C.; Chevalier, N.; Guild, R.M.; Munakata, Y. Developing adaptive control: Age-related differences in task choices and awareness of proactive and reactive control demands. Cogn. Affect. Behav. Neurosci. 2021, 21, 561–572. [Google Scholar] [CrossRef]
- Gonthier, C.; Blaye, A. Preschoolers are capable of fine-grained implicit cognitive control: Evidence from development of the context-specific proportion congruency effect. J. Exp. Child Psychol. 2021, 210, 105211. [Google Scholar] [CrossRef]
- Gonthier, C.; Ambrosi, S.; Blaye, A. Learning-based before intentional cognitive control: Developmental evidence for a dissociation between implicit and explicit control. J. Exp. Psychol. Learn. Mem. Cogn. 2021, 47, 1660–1685. [Google Scholar] [CrossRef]
- Mento, G.; Granziol, U. The developing predictive brain: How implicit temporal expectancy induced by local and global prediction shapes action preparation across development. Dev. Sci. 2020, 23, e12954. [Google Scholar] [CrossRef]
- Mento, G.; Scerif, G.; Granziol, U.; Franzoi, M.; Lanfranchi, S. The Effect of Probabilistic Context on Implicit Temporal Expectations in Down Syndrome. Front. Psychol. 2020, 11, 369. [Google Scholar] [CrossRef]
- Duma, G.M.; Danieli, A.; Morao, V.; Da Rold, M.; Baggio, M.; Toffoli, L.; Zanatta, A.; Vettorel, A.; Bonanni, P.; Mento, G. Implicit cognitive flexibility in self-limited focal epilepsy of childhood: An HD-EEG study. Epilepsy Behav. 2021, 116, 107747. [Google Scholar] [CrossRef]
- Sarant, J.Z.; Harris, D.C.; Bennet, L.A. Academic Outcomes for School-Aged Children with Severe–Profound Hearing Loss and Early Unilateral and Bilateral Cochlear Implants. J. Speech Lang. Hear. Res. 2015, 58, 1017–1032. [Google Scholar] [CrossRef]
- Fastelli, A.; Mento, G.; Marshall, C.R.; Arfé, B. Implicit learning of non-verbal regularities by deaf children with cochlear implants: An investigation with a dynamic temporal prediction task. PLoS ONE 2021, 16, e0251050. [Google Scholar] [CrossRef]
- Berg, E.A. A Simple Objective Technique for Measuring Flexibility in Thinking. J. Gen. Psychol. 1948, 39, 15–22. [Google Scholar] [CrossRef]
- Los, S.A.; Kruijne, W.; Meeter, M. Hazard versus history: Temporal preparation is driven by past experience. J. Exp. Psychol. Hum. Percept. Perform. 2017, 43, 78–88. [Google Scholar] [CrossRef]
- Raven, J.C.; Court, J.H.; Raven, J. Coloured Progress. Matrices; Oxford Psychologists Press: Oxford, UK, 1990; pp. 10–11. [Google Scholar]
- Belacchi, C.; Scalisi, T.G.; Cannoni, E.; Cornoldi, C. CPM—Coloured Progressive Matrices. Standarizzazione italiana. Manuale; Giunti O.S.: Firenze, Italy, 2008. [Google Scholar]
- Karlin, L. Reaction time as a function of foreperiod duration and variability. J. Exp. Psychol. 1959, 58, 185–191. [Google Scholar] [CrossRef]
- Los, S.A. Foreperiod and sequential effects: Theory and data. Atten. Time 2010, 289, 289–302. [Google Scholar]
- Luce, R.D. Response Times: Their Role in Inferring Elementary Mental Organization; Oxford University Press: Oxford, UK, 1986. [Google Scholar]
- Niemi, P.; Näätänen, R. Foreperiod and simple reaction time. Psychol. Bull. 1981, 89, 133–162. [Google Scholar] [CrossRef]
- Nobre, A.; Correa, A.; Coull, J. The hazards of time. Curr. Opin. Neurobiol. 2007, 17, 465–470. [Google Scholar] [CrossRef]
- Woodrow, H. The measurement of attention. Psychol. Monogr. Gen. Appl. 1914, 17, i-158. [Google Scholar] [CrossRef]
- Mento, G. The role of the P3 and CNV components in voluntary and automatic temporal orienting: A high spatial-resolution ERP study. Neuropsychologia 2017, 107, 31–40. [Google Scholar] [CrossRef]
- Mento, G.; Tarantino, V. Developmental Trajectories of Internally and Externally Driven Temporal Prediction. PLoS ONE 2015, 10, e0135098. [Google Scholar] [CrossRef]
- Vallesi, A. Neuro-Anatomical Substrates of Foreperiod Effects. Attention and Time; Oxford University Press: Oxford, UK, 2010; pp. 303–316. [Google Scholar]
- Johnson, K.A.; Burrowes, E.; Coull, J.T. Children Can Implicitly, but Not Voluntarily, Direct Attention in Time. PLoS ONE 2015, 10, e0123625. [Google Scholar] [CrossRef]
- Vallesi, A.; Shallice, T. Developmental dissociations of preparation over time: Deconstructing the variable foreperiod phenomena. J. Exp. Psychol. Hum. Percept. Perform. 2007, 33, 1377–1388. [Google Scholar] [CrossRef]
- Trillenberg, P.; Verleger, R.; Wascher, E.; Wauschkuhn, B.; Wessel, K. CNV and temporal uncertainty with ‘ageing’ and ‘non-ageing’ S1–S2 intervals. Clin. Neurophysiol. 2000, 111, 1216–1226. [Google Scholar] [CrossRef]
- Delorme, A.; Makeig, S. EEGLAB: An Open Source Toolbox for Analysis of Single-Trial EEG Dynamics Including Independent Component Analysis. J. Neurosci. Methods 2004, 134, 9–21. [Google Scholar] [CrossRef]
- Stone, J.V. Independent component analysis: An introduction. Trends Cogn. Sci. 2002, 6, 59–64. [Google Scholar] [CrossRef]
- Bell, A.J.; Sejnowski, T.J. An Information-Maximization Approach to Blind Separation and Blind Deconvolution. Neural Comput. 1995, 7, 1129–1159. [Google Scholar] [CrossRef]
- Perrin, F.; Pernier, J.; Bertrand, O.; Echallier, J.F. Spherical splines for scalp potential and current density mapping. Electroencephalogr. Clin. Neurophysiol. 1989, 72, 184–187. [Google Scholar] [CrossRef]
- Tadel, F.; Baillet, S.; Mosher, J.C.; Pantazis, D.; Leahy, R.M. Brainstorm: A User-Friendly Application for MEG/EEG Analysis. Comput. Intell. Neurosci. 2011, 2011, 879716. [Google Scholar] [CrossRef]
- Kotowski, K.; Stapor, K.; Leski, J. Improved robust weighted averaging for event-related potentials in EEG. Biocybern. Biomed. Eng. 2019, 39, 1036–1046. [Google Scholar] [CrossRef]
- Groppe, D.M.; Urbach, T.P.; Kutas, M. Mass univariate analysis of event-related brain potentials/fields I: A critical tutorial review. Psychophysiology 2011, 48, 1711–1725. [Google Scholar] [CrossRef]
- Maris, E.; Oostenveld, R. Nonparametric statistical testing of EEG-and MEG-dataJ. Neurosci. Methods 2007, 164, 177–190. [Google Scholar] [CrossRef]
- Walter, W.G.; Cooper, R.; Aldridge, V.J.; McCallum, W.C.; Winter, A.L. Contingent Negative Variation: An Electric Sign of Sensori-Motor Association and Expectancy in the Human Brain. Nature 1964, 203, 380–384. [Google Scholar] [CrossRef]
- Mento, G. The passive CNV: Carving out the contribution of task-related processes to expectancy. Front. Hum. Neurosci. 2013, 7. [Google Scholar] [CrossRef]
- Evans, A.C.; Janke, A.L.; Collins, D.L.; Baillet, S. Brain templates and atlases. NeuroImage 2012, 62, 911–922. [Google Scholar] [CrossRef]
- Gramfort, A.; Papadopoulo, T.; Olivi, E.; Clerc, M. Forward Field Computation with OpenMEEG. Comput. Intell. Neurosci. 2011, 2011, 923703. [Google Scholar] [CrossRef]
- Kybic, J.; Clerc, M.; Abboud, T.; Faugeras, O.; Keriven, R.; Papadopoulo, T. A common formalism for the Integral formulations of the forward EEG problem. IEEE Trans. Med. Imaging 2005, 24, 12–28. [Google Scholar] [CrossRef]
- Desikan, R.S.; Ségonne, F.; Fischl, B.; Quinn, B.T.; Dickerson, B.C.; Blacker, D.; Buckner, R.L.; Dale, A.M.; Maguire, R.P.; Hyman, B.T.; et al. An automated labeling system for subdividing the human cerebral cortex on MRI scans into gyral based regions of interest. NeuroImage 2006, 31, 968–980. [Google Scholar] [CrossRef]
- James, S.-N.; Rommel, A.-S.; Cheung, C.; McLoughlin, G.; Brandeis, D.; Banaschewski, T.; Asherson, P.; Kuntsi, J. Association of preterm birth with ADHD-like cognitive impairments and additional subtle impairments in attention and arousal malleability. Psychol. Med. 2018, 48, 1484–1493. [Google Scholar] [CrossRef]
- Rommel, A.-S.; James, S.-N.; McLoughlin, G.; Brandeis, D.; Banaschewski, T.; Asherson, P.; Kuntsi, J. Association of Preterm Birth With Attention-Deficit/Hyperactivity Disorder–Like and Wider-Ranging Neurophysiological Impairments of Attention and Inhibition. J. Am. Acad. Child Adolesc. Psychiatry 2017, 56, 40–50. [Google Scholar] [CrossRef]
- Mulder, H.; Pitchford, N.J.; Marlow, N. Processing Speed Mediates Executive Function Difficulties in Very Preterm Children in Middle Childhood. J. Int. Neuropsychol. Soc. 2011, 17, 445–454. [Google Scholar] [CrossRef]
- Pizzo, R.; Urben, S.; VAN DER Linden, M.; Borradori-Tolsa, C.; Freschi, M.; Forcada-Guex, M.; Hüppi, P.; Barisnikov, K. Attentional networks efficiency in preterm children. J. Int. Neuropsychol. Soc. 2010, 16, 130–137. [Google Scholar] [CrossRef]
- Réveillon, M.; Hüppi, P.S.; Barisnikov, K. Inhibition difficulties in preterm children: Developmental delay or persistent deficit? Child Neuropsychol. 2018, 24, 734–762. [Google Scholar] [CrossRef]
- Réveillon, M.; Tolsa, C.B.; Monnier, M.; Hüppi, P.S.; Barisnikov, K. Response inhibition difficulties in preterm children aged 9–12 years: Relations with emotion and behavior. Child Neuropsychol. 2016, 22, 420–442. [Google Scholar] [CrossRef]
- Murray, A.L.; Scratch, S.E.; Thompson, D.K.; Inder, T.E.; Doyle, L.W.; Anderson, J.F.I.; Anderson, P.J. Neonatal brain pathology predicts adverse attention and processing speed outcomes in very preterm and/or very low birth weight children. Neuropsychology 2014, 28, 552–562. [Google Scholar] [CrossRef]
- Mento, G.; Tarantino, V.; Sarlo, M.; Bisiacchi, P.S. Automatic Temporal Expectancy: A High-Density Event-Related Potential Study. PLoS ONE 2013, 8, e62896. [Google Scholar] [CrossRef]
- Rogers, C.E.; Lean, R.E.; Wheelock, M.D.; Smyser, C.D. Aberrant structural and functional connectivity and neurodevelopmental impairment in preterm children. J. Neurodev. Disord. 2018, 10, 38. [Google Scholar] [CrossRef]
- Thomason, M.E.; Scheinost, D.; Manning, J.H.; Grove, L.E.; Hect, J.; Marshall, N.; Hernandez-Andrade, E.; Berman, S.; Pappas, A.; Yeo, L.; et al. Weak functional connectivity in the human fetal brain prior to preterm birth. Sci. Rep. 2017, 7, 39286. [Google Scholar] [CrossRef]
- Mento, G.; Scerif, G.; Granziol, U.; Franzoi, M.; Lanfranchi, S. Dissociating top-down and bottom-up temporal attention in Down syndrome: A neurocostructive perspective. Cogn. Dev. 2019, 49, 81–93. [Google Scholar] [CrossRef]
- Cristaldi, F.D.P.; Toffoli, L.; Duma, G.M.; Mento, G. Little fast, little slow, should I stay or should I go? Flexibility in adapting motor control to local-global temporal prediction across typical development. NeuroImage 2022, 220, 117071. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PT | FT | |
|---|---|---|
| N Tot (% Males) | 21 (52%) | 21 (42%) |
| Age in Years M (SD) | 8.42 (1.3) | 8.19 (1.4) |
| Gestational Age in Days M (SD, Range) | 213 (20.9; range 168–252) | >270 |
| Birth Weight in Grams M (SD) | 1448 (623) | >2500 |
| BLOCK | FAST | UNIFORM | SLOW | ||||||
|---|---|---|---|---|---|---|---|---|---|
| SOA | Short | Medium | Long | Short | Medium | Long | Short | Medium | Long |
| PT | 94.3 (6.1) | 93.1 (7.1) | 93.8 (9.4) | 92.3 (5.6) | 95.7 (4.6) | 93.8 (5.3) | 92.2 (7.4) | 93.7 (7.1) | 92.9 (6.4) |
| FT | 97.7 (2.2) | 96.8 (3.7) | 91.8 (5.7) | 97.6 (4) | 96.9 (4.1) | 96.7 (4.4) | 98.6 (4.9) | 96.8 (4) | 97 (2.9) |
| BLOCK | FAST | UNIFORM | SLOW | ||||||
|---|---|---|---|---|---|---|---|---|---|
| SOA | Short | Medium | Long | Short | Medium | Long | Short | Medium | Long |
| PT | 477 (84) | 418 (88) | 402 (90) | 508 (88) | 441 (99) | 402 (70) | 521 (86) | 444 (92) | 420 (79) |
| FT | 507 (76) | 444 (80) | 438 (86) | 570 (68) | 492 (86) | 457 (68) | 602 (92) | 510 (86) | 475 (86) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mento, G.; Toffoli, L.; Della Longa, L.; Farroni, T.; Del Popolo Cristaldi, F.; Duma, G.M. Adaptive Cognitive Control in Prematurely Born Children: An HD-EEG Investigation. Brain Sci. 2022, 12, 1074. https://doi.org/10.3390/brainsci12081074
Mento G, Toffoli L, Della Longa L, Farroni T, Del Popolo Cristaldi F, Duma GM. Adaptive Cognitive Control in Prematurely Born Children: An HD-EEG Investigation. Brain Sciences. 2022; 12(8):1074. https://doi.org/10.3390/brainsci12081074
Chicago/Turabian StyleMento, Giovanni, Lisa Toffoli, Letizia Della Longa, Teresa Farroni, Fiorella Del Popolo Cristaldi, and Gian Marco Duma. 2022. "Adaptive Cognitive Control in Prematurely Born Children: An HD-EEG Investigation" Brain Sciences 12, no. 8: 1074. https://doi.org/10.3390/brainsci12081074