
Ultrasound-Mediated Bioeffects in Senescent Mice and Alzheimer’s Mouse Models


{kind=link}
Abstract
:1. Introduction to Therapeutic Ultrasound
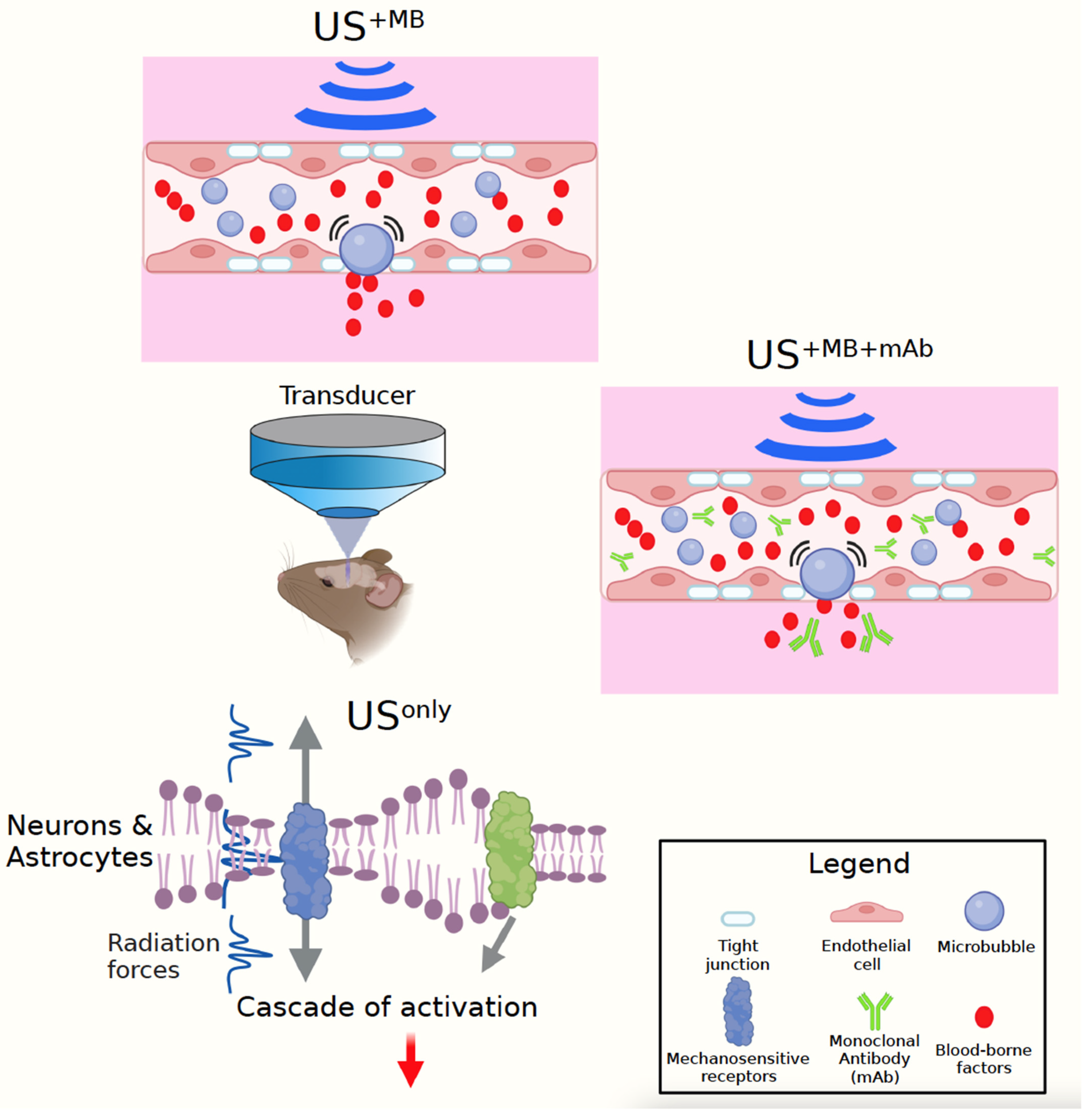
2. Applications of Low-Frequency Ultrasound to Alzheimer’s Mouse Models
3. Ultrasound-Mediated Neuromodulation to Treat Dementia—Lessons from Senescent Mice
4. Conclusions and Outlook
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Karran, E.; De Strooper, B. The amyloid hypothesis in Alzheimer disease: New insights from new therapeutics. Nat. Rev. Drug Discov. 2022, 21, 306–318. [Google Scholar] [CrossRef] [PubMed]
- Koch, G.; Bonni, S.; Pellicciari, M.C.; Casula, E.P.; Mancini, M.; Esposito, R.; Ponzo, V.; Picazio, S.; Di Lorenzo, F.; Serra, L.; et al. Transcranial magnetic stimulation of the precuneus enhances memory and neural activity in prodromal Alzheimer’s disease. NeuroImage 2018, 169, 302–311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Targum, S.D.; Fosdick, L.; Drake, K.E.; Rosenberg, P.B.; Burke, A.D.; Wolk, D.A.; Foote, K.D.; Asaad, W.F.; Sabbagh, M.; Smith, G.S.; et al. Effect of Age on Clinical Trial Outcome in Participants with Probable Alzheimer’s Disease. J. Alzheimers Dis. 2021, 82, 1243–1257. [Google Scholar] [CrossRef] [PubMed]
- Burgess, A.; Hynynen, K. Microbubble-Assisted Ultrasound for Drug Delivery in the Brain and Central Nervous System. Adv. Exp. Med. Biol. 2016, 880, 293–308. [Google Scholar] [CrossRef]
- Elias, W.J.; Lipsman, N.; Ondo, W.G.; Ghanouni, P.; Kim, Y.G.; Lee, W.; Schwartz, M.; Hynynen, K.; Lozano, A.M.; Shah, B.B.; et al. A Randomized Trial of Focused Ultrasound Thalamotomy for Essential Tremor. N. Engl. J. Med. 2016, 375, 730–739. [Google Scholar] [CrossRef]
- Sinai, A.; Nassar, M.; Sprecher, E.; Constantinescu, M.; Zaaroor, M.; Schlesinger, I. Focused Ultrasound Thalamotomy in Tremor Dominant Parkinson’s Disease: Long-Term Results. J. Parkinson’s Dis. 2022, 12, 199–206. [Google Scholar] [CrossRef]
- Cohen-Inbar, O.; Xu, Z.; Sheehan, J.P. Focused ultrasound-aided immunomodulation in glioblastoma multiforme: A therapeutic concept. J. Ther. Ultrasound 2016, 4, 2. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, M.; Kobayashi, H.; Terasaka, S.; Endo, S.; Yamaguchi, S.; Motegi, H.; Itay, R.; Suzuki, S.; Brokman, O.; Shapira, Y.; et al. Sonodynamic Therapy for Malignant Glioma Using 220-kHz Transcranial Magnetic Resonance Imaging-Guided Focused Ultrasound and 5-Aminolevulinic acid. Ultrasound Med. Biol. 2019, 45, 526–538. [Google Scholar] [CrossRef]
- Baron, C.; Aubry, J.F.; Tanter, M.; Meairs, S.; Fink, M. Simulation of intracranial acoustic fields in clinical trials of sonothrombolysis. Ultrasound Med. Biol. 2009, 35, 1148–1158. [Google Scholar] [CrossRef]
- Alexandrov, A.V.; Kohrmann, M.; Soinne, L.; Tsivgoulis, G.; Barreto, A.D.; Demchuk, A.M.; Sharma, V.K.; Mikulik, R.; Muir, K.W.; Brandt, G.; et al. Safety and efficacy of sonothrombolysis for acute ischaemic stroke: A multicentre, double-blind, phase 3, randomised controlled trial. Lancet Neurol. 2019, 18, 338–347. [Google Scholar] [CrossRef] [Green Version]
- Meng, Y.; Hynynen, K.; Lipsman, N. Applications of focused ultrasound in the brain: From thermoablation to drug delivery. Nat. Rev. Neurol. 2021, 17, 7–22. [Google Scholar] [CrossRef] [PubMed]
- Naor, O.; Krupa, S.; Shoham, S. Ultrasonic neuromodulation. J. Neural Eng. 2016, 13, 031003. [Google Scholar] [CrossRef] [PubMed]
- Tyler, W.J.; Lani, S.W.; Hwang, G.M. Ultrasonic modulation of neural circuit activity. Curr. Opin. Neurobiol. 2018, 50, 222–231. [Google Scholar] [CrossRef] [PubMed]
- Munoz, F.; Aurup, C.; Konofagou, E.E.; Ferrera, V.P. Modulation of Brain Function and Behavior by Focused Ultrasound. Curr. Behav. Neurosci. Rep. 2018, 5, 153–164. [Google Scholar] [CrossRef] [PubMed]
- Blackmore, J.; Shrivastava, S.; Sallet, J.; Butler, C.R.; Cleveland, R.O. Ultrasound Neuromodulation: A Review of Results, Mechanisms and Safety. Ultrasound Med. Biol. 2019, 45, 1509–1536. [Google Scholar] [CrossRef] [Green Version]
- Rabut, C.; Yoo, S.; Hurt, R.C.; Jin, Z.; Li, H.; Guo, H.; Ling, B.; Shapiro, M.G. Ultrasound Technologies for Imaging and Modulating Neural Activity. Neuron 2020, 108, 93–110. [Google Scholar] [CrossRef]
- Götz, J.; Richter-Stretton, G.; Cruz, E. Therapeutic Ultrasound as a Treatment Modality for Physiological and Pathological Ageing Including Alzheimer’s Disease. Pharmaceutics 2021, 13, 1002. [Google Scholar] [CrossRef]
- Venturino, A.; Schulz, R.; De Jesus-Cortes, H.; Maes, M.E.; Nagy, B.; Reilly-Andujar, F.; Colombo, G.; Cubero, R.J.A.; Schoot Uiterkamp, F.E.; Bear, M.F.; et al. Microglia enable mature perineuronal nets disassembly upon anesthetic ketamine exposure or 60-Hz light entrainment in the healthy brain. Cell Rep. 2021, 36, 109313. [Google Scholar] [CrossRef]
- Blackmore, D.G.; Turpin, F.; Palliyaguru, T.; Evans, H.T.; Chicoteau, A.; Lee, W.; Pelekanos, M.; Nguyen, N.; Song, J.; Sullivan, R.K.P.; et al. Low-intensity ultrasound restores long-term potentiation and memory in senescent mice through pleiotropic mechanisms including NMDAR signaling. Mol. Psychiatry 2021, 26, 6975–6991. [Google Scholar] [CrossRef]
- Krasovitski, B.; Frenkel, V.; Shoham, S.; Kimmel, E. Intramembrane cavitation as a unifying mechanism for ultrasound-induced bioeffects. Proc. Natl. Acad. Sci. USA 2011, 108, 3258–3263. [Google Scholar] [CrossRef] [Green Version]
- Gateau, J.; Aubry, J.F.; Chauvet, D.; Boch, A.L.; Fink, M.; Tanter, M. In vivo bubble nucleation probability in sheep brain tissue. Phys. Med. Biol. 2011, 56, 7001–7015. [Google Scholar] [CrossRef]
- Oh, S.J.; Lee, J.M.; Kim, H.B.; Lee, J.; Han, S.; Bae, J.Y.; Hong, G.S.; Koh, W.; Kwon, J.; Hwang, E.S.; et al. Ultrasonic Neuromodulation via Astrocytic TRPA1. Curr. Biol. 2020, 30, 948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoo, S.; Mittelstein, D.R.; Hurt, R.C.; Lacroix, J.; Shapiro, M.G. Focused ultrasound excites cortical neurons via mechanosensitive calcium accumulation and ion channel amplification. Nat. Commun. 2022, 13, 493. [Google Scholar] [CrossRef] [PubMed]
- Nisbet, R.M.; van der Jeugd, A.; Leinenga, G.; Evans, H.T.; Janowicz, P.W.; Götz, J. Combined effects of scanning ultrasound and a tau-specific single chain antibody in a tau transgenic mouse model. Brain 2017, 140, 1220–1230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, R.M.; Hwang, Y.C.; Liu, I.J.; Lee, C.C.; Tsai, H.Z.; Li, H.J.; Wu, H.C. Development of therapeutic antibodies for the treatment of diseases. J. Biomed. Sci. 2020, 27, 1. [Google Scholar] [CrossRef] [PubMed]
- Golde, T.E. Open questions for Alzheimer’s disease immunotherapy. Alzheimers Res. Ther. 2014, 6, 3. [Google Scholar] [CrossRef]
- Goedert, M.; Spillantini, M.G. A century of Alzheimer’s disease. Science 2006, 314, 777–781. [Google Scholar] [CrossRef] [Green Version]
- Hardy, J. Alzheimer’s disease: The amyloid cascade hypothesis: An update and reappraisal. J. Alzheimers Dis. 2006, 9, 151–153. [Google Scholar] [CrossRef] [Green Version]
- Ittner, L.M.; Ke, Y.D.; Delerue, F.; Bi, M.; Gladbach, A.; van Eersel, J.; Wolfing, H.; Chieng, B.C.; Christie, M.J.; Napier, I.A.; et al. Dendritic function of tau mediates amyloid-beta toxicity in Alzheimer’s disease mouse models. Cell 2010, 142, 387–397. [Google Scholar] [CrossRef] [Green Version]
- Ittner, L.M.; Götz, J. Amyloid-beta and tau—A toxic pas de deux in Alzheimer’s disease. Nat. Rev. Neurosci. 2011, 12, 65–72. [Google Scholar] [CrossRef]
- Götz, J.; Bodea, L.G.; Goedert, M. Rodent models for Alzheimer disease. Nat. Rev. Neurosci. 2018, 19, 583–598. [Google Scholar] [CrossRef] [PubMed]
- Jordao, J.F.; Thevenot, E.; Markham-Coultes, K.; Scarcelli, T.; Weng, Y.Q.; Xhima, K.; O’Reilly, M.; Huang, Y.; McLaurin, J.; Hynynen, K.; et al. Amyloid-beta plaque reduction, endogenous antibody delivery and glial activation by brain-targeted, transcranial focused ultrasound. Exp. Neurol. 2013, 248, 16–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leinenga, G.; Götz, J. Scanning ultrasound removes amyloid-beta and restores memory in an Alzheimer’s disease mouse model. Sci. Transl. Med. 2015, 7, 278ra233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leinenga, G.; Bodea, L.G.; Schröder, J.; Sun, G.; Zhou, Y.; Song, J.; Grubman, A.; Polo, J.M.; Götz, J. Transcriptional signature in microglia isolated from an Alzheimer’s disease mouse model treated with scanning ultrasound. Bioeng. Transl. Med. 2021. [Google Scholar] [CrossRef]
- Mathew, A.S.; Gorick, C.M.; Price, R.J. Multiple regression analysis of a comprehensive transcriptomic data assembly elucidates mechanically- and biochemically-driven responses to focused ultrasound blood-brain barrier disruption. Theranostics 2021, 11, 9847–9858. [Google Scholar] [CrossRef]
- Poon, C.T.; Shah, K.; Lin, C.; Tse, R.; Kim, K.K.; Mooney, S.; Aubert, I.; Stefanovic, B.; Hynynen, K. Time course of focused ultrasound effects on beta-amyloid plaque pathology in the TgCRND8 mouse model of Alzheimer’s disease. Sci. Rep. 2018, 8, 14061. [Google Scholar] [CrossRef]
- Poon, C.; Pellow, C.; Hynynen, K. Neutrophil recruitment and leukocyte response following focused ultrasound and microbubble mediated blood-brain barrier treatments. Theranostics 2021, 11, 1655–1671. [Google Scholar] [CrossRef]
- Hsu, P.H.; Lin, Y.T.; Chung, Y.H.; Lin, K.J.; Yang, L.Y.; Yen, T.C.; Liu, H.L. Focused Ultrasound-Induced Blood-Brain Barrier Opening Enhances GSK-3 Inhibitor Delivery for Amyloid-Beta Plaque Reduction. Sci. Rep. 2018, 8, 12882. [Google Scholar] [CrossRef]
- Xhima, K.; Markham-Coultes, K.; Kofoed, R.H.; Saragovi, H.U.; Hynynen, K.; Aubert, I. Ultrasound delivery of a TrkA agonist confers neuroprotection to Alzheimer-associated pathologies. Brain 2021, awab460. [Google Scholar] [CrossRef]
- Deng, Z.; Wang, J.; Xiao, Y.; Li, F.; Niu, L.; Liu, X.; Meng, L.; Zheng, H. Ultrasound-mediated augmented exosome release from astrocytes alleviates amyloid-beta-induced neurotoxicity. Theranostics 2021, 11, 4351–4362. [Google Scholar] [CrossRef]
- Chan, T.G.; Ruehl, C.L.; Morse, S.V.; Simon, M.; Rakers, V.; Watts, H.; Aprile, F.A.; Choi, J.J.; Vilar, R. Modulation of amyloid-beta aggregation by metal complexes with a dual binding mode and their delivery across the blood-brain barrier using focused ultrasound. Chem. Sci. 2021, 12, 9485–9493. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Xu, X.; Chen, B.; Liao, Y.; Guan, X.; He, Y.; Cui, H.; Rong, Y.; Liu, Z.; Xu, Y. Ultrasound-targeted microbubbles destruction assists dual delivery of beta-amyloid antibody and neural stem cells to restore neural function in transgenic mice of Alzheimer’s disease. Med. Phys. 2022, 49, 1357–1367. [Google Scholar] [CrossRef] [PubMed]
- Leinenga, G.; Koh, W.K.; Götz, J. Scanning ultrasound in the absence of blood-brain barrier opening is not sufficient to clear beta-amyloid plaques in the APP23 mouse model of Alzheimer’s disease. Brain Res. Bull. 2019, 153, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Eguchi, K.; Shindo, T.; Ito, K.; Ogata, T.; Kurosawa, R.; Kagaya, Y.; Monma, Y.; Ichijo, S.; Kasukabe, S.; Miyata, S.; et al. Whole-brain low-intensity pulsed ultrasound therapy markedly improves cognitive dysfunctions in mouse models of dementia—Crucial roles of endothelial nitric oxide synthase. Brain Stimul. 2018, 11, 959–973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karakatsani, M.E.; Kugelman, T.; Ji, R.; Murillo, M.; Wang, S.; Niimi, Y.; Small, S.A.; Duff, K.E.; Konofagou, E.E. Unilateral Focused Ultrasound-Induced Blood-Brain Barrier Opening Reduces Phosphorylated Tau from The rTg4510 Mouse Model. Theranostics 2019, 9, 5396–5411. [Google Scholar] [CrossRef]
- Pandit, R.; Leinenga, G.; Götz, J. Repeated ultrasound treatment of tau transgenic mice clears neuronal tau by autophagy and improves behavioral functions. Theranostics 2019, 9, 3754–3767. [Google Scholar] [CrossRef]
- Caballero, B.; Wang, Y.; Diaz, A.; Tasset, I.; Juste, Y.R.; Stiller, B.; Mandelkow, E.M.; Mandelkow, E.; Cuervo, A.M. Interplay of pathogenic forms of human tau with different autophagic pathways. Aging Cell 2018, 17, e12692. [Google Scholar] [CrossRef] [Green Version]
- Shen, Y.; Hua, L.; Yeh, C.K.; Shen, L.; Ying, M.; Zhang, Z.; Liu, G.; Li, S.; Chen, S.; Chen, X.; et al. Ultrasound with microbubbles improves memory, ameliorates pathology and modulates hippocampal proteomic changes in a triple transgenic mouse model of Alzheimer’s disease. Theranostics 2020, 10, 11794–11819. [Google Scholar] [CrossRef]
- Janowicz, P.W.; Leinenga, G.; Götz, J.; Nisbet, R.M. Ultrasound-mediated blood-brain barrier opening enhances delivery of therapeutically relevant formats of a tau-specific antibody. Sci. Rep. 2019, 9, 9255. [Google Scholar] [CrossRef] [Green Version]
- Jordao, J.F.; Ayala-Grosso, C.A.; Markham, K.; Huang, Y.; Chopra, R.; McLaurin, J.; Hynynen, K.; Aubert, I. Antibodies targeted to the brain with image-guided focused ultrasound reduces amyloid-beta plaque load in the TgCRND8 mouse model of Alzheimer’s disease. PLoS ONE 2010, 5, e10549. [Google Scholar] [CrossRef] [Green Version]
- Leinenga, G.; Koh, W.K.; Götz, J. A comparative study of the effects of Aducanumab and scanning ultrasound on amyloid plaques and behavior in the APP23 mouse model of Alzheimer disease. Alzheimers Res. Ther. 2021, 13, 76. [Google Scholar] [CrossRef]
- Sun, T.; Shi, Q.; Zhang, Y.; Power, C.; Hoesch, C.; Antonelli, S.; Schroeder, M.K.; Caldarone, B.J.; Taudte, N.; Schenk, M.; et al. Focused ultrasound with anti-pGlu3 Abeta enhances efficacy in Alzheimer’s disease-like mice via recruitment of peripheral immune cells. J. Control. Release 2021, 336, 443–456. [Google Scholar] [CrossRef]
- Lipsman, N.; Meng, Y.; Bethune, A.J.; Huang, Y.; Lam, B.; Masellis, M.; Herrmann, N.; Heyn, C.; Aubert, I.; Boutet, A.; et al. Blood-brain barrier opening in Alzheimer’s disease using MR-guided focused ultrasound. Nat. Commun. 2018, 9, 2336. [Google Scholar] [CrossRef] [Green Version]
- Park, S.H.; Baik, K.; Jeon, S.; Chang, W.S.; Ye, B.S.; Chang, J.W. Extensive frontal focused ultrasound mediated blood-brain barrier opening for the treatment of Alzheimer’s disease: A proof-of-concept study. Transl. Neurodegener. 2021, 10, 44. [Google Scholar] [CrossRef]
- Epelbaum, S.; Burgos, N.; Canney, M.; Matthews, D.; Houot, M.; Santin, M.D.; Desseaux, C.; Bouchoux, G.; Stroer, S.; Martin, C.; et al. Pilot study of repeated blood-brain barrier disruption in patients with mild Alzheimer’s disease with an implantable ultrasound device. Alzheimers Res. Ther. 2022, 14, 40. [Google Scholar] [CrossRef] [PubMed]
- Pouliopoulos, A.N.; Wu, S.Y.; Burgess, M.T.; Karakatsani, M.E.; Kamimura, H.A.S.; Konofagou, E.E. A Clinical System for Non-invasive Blood-Brain Barrier Opening Using a Neuronavigation-Guided Single-Element Focused Ultrasound Transducer. Ultrasound Med. Biol. 2020, 46, 73–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turturro, A.; Duffy, P.; Hass, B.; Kodell, R.; Hart, R. Survival characteristics and age-adjusted disease incidences in C57BL/6 mice fed a commonly used cereal-based diet modulated by dietary restriction. J. Gerontol. A Biol. Sci. Med. Sci. 2002, 57, B379–B389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez-Otin, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The hallmarks of aging. Cell 2013, 153, 1194–1217. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, K.; Agarwal, P.; Haggerty, G. Alzheimer’s disease—Not an exaggeration of healthy aging. Indian J. Psychol. Med. 2011, 33, 106–114. [Google Scholar] [CrossRef] [Green Version]
- Pievani, M.; Agosta, F.; Pagani, E.; Canu, E.; Sala, S.; Absinta, M.; Geroldi, C.; Ganzola, R.; Frisoni, G.B.; Filippi, M. Assessment of white matter tract damage in mild cognitive impairment and Alzheimer’s disease. Hum. Brain Mapp. 2010, 31, 1862–1875. [Google Scholar] [CrossRef]
- Fjell, A.M.; Amlien, I.K.; Westlye, L.T.; Stenset, V.; Fladby, T.; Skinningsrud, A.; Eilsertsen, D.E.; Bjornerud, A.; Walhovd, K.B. CSF biomarker pathology correlates with a medial temporo-parietal network affected by very mild to moderate Alzheimer’s disease but not a fronto-striatal network affected by healthy aging. NeuroImage 2010, 49, 1820–1830. [Google Scholar] [CrossRef] [PubMed]
- Irwin, K.; Sexton, C.; Daniel, T.; Lawlor, B.; Naci, L. Healthy Aging and Dementia: Two Roads Diverging in Midlife? Front. Aging Neurosci. 2018, 10, 275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willis, E.F.; Bartlett, P.F.; Vukovic, J. Protocol for Short- and Longer-term Spatial Learning and Memory in Mice. Front. Behav. Neurosci. 2017, 11, 197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benice, T.S.; Rizk, A.; Kohama, S.; Pfankuch, T.; Raber, J. Sex-differences in age-related cognitive decline in C57BL/6J mice associated with increased brain microtubule-associated protein 2 and synaptophysin immunoreactivity. Neuroscience 2006, 137, 413–423. [Google Scholar] [CrossRef] [PubMed]
- van Praag, H.; Shubert, T.; Zhao, C.; Gage, F.H. Exercise enhances learning and hippocampal neurogenesis in aged mice. J. Neurosci. 2005, 25, 8680–8685. [Google Scholar] [CrossRef] [PubMed]
- Balbi, M.; Ghosh, M.; Longden, T.A.; Jativa Vega, M.; Gesierich, B.; Hellal, F.; Lourbopoulos, A.; Nelson, M.T.; Plesnila, N. Dysfunction of mouse cerebral arteries during early aging. J. Cereb. Blood Flow Metab. 2015, 35, 1445–1453. [Google Scholar] [CrossRef] [PubMed]
- Hatch, R.J.; Leinenga, G.; Götz, J. Scanning ultrasound (SUS) causes no changes to neuronal excitability and prevents age-related reductions in hippocampal CA1 dendritic structure in wild-type mice. PLoS ONE 2016, 11, e0164278. [Google Scholar] [CrossRef] [Green Version]
- Blackmore, D.G.; Turpin, F.; Mohamed, A.Z.; Zong, F.; Pandit, R.; Pelekanos, M.; Nasrallah, F.; Sah, P.; Bartlett, P.F.; Götz, J. Multimodal analysis of aged wild-type mice exposed to repeated scanning ultrasound treatments demonstrates long-term safety. Theranostics 2018, 8, 6233–6247. [Google Scholar] [CrossRef]
- Scarcelli, T.; Jordao, J.F.; O’Reilly, M.A.; Ellens, N.; Hynynen, K.; Aubert, I. Stimulation of hippocampal neurogenesis by transcranial focused ultrasound and microbubbles in adult mice. Brain Stimul. 2014, 7, 304–307. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Lin, Z.; Wang, K.; Liu, X.; Zhou, W.; Meng, L.; Huang, J.; Yuan, K.; Niu, L.; Zheng, H. Transcranial Low-Intensity Pulsed Ultrasound Modulates Structural and Functional Synaptic Plasticity in Rat Hippocampus. IEEE Trans. Ultrason. Ferroelectr. Freq. Control 2019, 66, 930–938. [Google Scholar] [CrossRef]
- Tyler, W.J.; Tufail, Y.; Finsterwald, M.; Tauchmann, M.L.; Olson, E.J.; Majestic, C. Remote excitation of neuronal circuits using low-intensity, low-frequency ultrasound. PLoS ONE 2008, 3, e3511. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Z.; Wang, C.; Wei, B.; Gan, W.; Zhou, Q.; Cui, M. High resolution ultrasonic neural modulation observed via in vivo two-photon calcium imaging. Brain Stimul. 2022, 15, 190–196. [Google Scholar] [CrossRef] [PubMed]
- Beisteiner, R.; Matt, E.; Fan, C.; Baldysiak, H.; Schonfeld, M.; Philippi Novak, T.; Amini, A.; Aslan, T.; Reinecke, R.; Lehrner, J.; et al. Transcranial Pulse Stimulation with Ultrasound in Alzheimer’s Disease-A New Navigated Focal Brain Therapy. Adv. Sci. 2020, 7, 1902583. [Google Scholar] [CrossRef] [Green Version]
- Popescu, T.; Pernet, C.; Beisteiner, R. Transcranial ultrasound pulse stimulation reduces cortical atrophy in Alzheimer’s patients: A follow-up study. Alzheimers Dement. 2021, 7, e12121. [Google Scholar] [CrossRef] [PubMed]
- Mueller, J.; Legon, W.; Opitz, A.; Sato, T.F.; Tyler, W.J. Transcranial focused ultrasound modulates intrinsic and evoked EEG dynamics. Brain Stimul. 2014, 7, 900–908. [Google Scholar] [CrossRef] [Green Version]
- Legon, W.; Sato, T.F.; Opitz, A.; Mueller, J.; Barbour, A.; Williams, A.; Tyler, W.J. Transcranial focused ultrasound modulates the activity of primary somatosensory cortex in humans. Nat. Neurosci. 2014, 17, 322–329. [Google Scholar] [CrossRef]
- Lee, W.; Kim, H.; Jung, Y.; Song, I.U.; Chung, Y.A.; Yoo, S.S. Image-guided transcranial focused ultrasound stimulates human primary somatosensory cortex. Sci. Rep. 2015, 5, 8743. [Google Scholar] [CrossRef] [Green Version]
- Lee, W.; Chung, Y.A.; Jung, Y.; Song, I.U.; Yoo, S.S. Simultaneous acoustic stimulation of human primary and secondary somatosensory cortices using transcranial focused ultrasound. BMC Neurosci. 2016, 17, 68. [Google Scholar] [CrossRef] [Green Version]
- Monti, M.M.; Schnakers, C.; Korb, A.S.; Bystritsky, A.; Vespa, P.M. Non-Invasive Ultrasonic Thalamic Stimulation in Disorders of Consciousness after Severe Brain Injury: A First-in-Man Report. Brain Stimul. 2016, 9, 940–941. [Google Scholar] [CrossRef]
- Lee, W.; Kim, H.C.; Jung, Y.; Chung, Y.A.; Song, I.U.; Lee, J.H.; Yoo, S.S. Transcranial focused ultrasound stimulation of human primary visual cortex. Sci. Rep. 2016, 6, 34026. [Google Scholar] [CrossRef] [Green Version]
- Lee, W.; Kim, S.; Kim, B.; Lee, C.; Chung, Y.A.; Kim, L.; Yoo, S.S. Non-invasive transmission of sensorimotor information in humans using an EEG/focused ultrasound brain-to-brain interface. PLoS ONE 2017, 12, e0178476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ai, L.; Bansal, P.; Mueller, J.K.; Legon, W. Effects of transcranial focused ultrasound on human primary motor cortex using 7T fMRI: A pilot study. BMC Neurosci. 2018, 19, 56. [Google Scholar] [CrossRef] [PubMed]
- Legon, W.; Bansal, P.; Tyshynsky, R.; Ai, L.; Mueller, J.K. Transcranial focused ultrasound neuromodulation of the human primary motor cortex. Sci. Rep. 2018, 8, 10007. [Google Scholar] [CrossRef] [Green Version]
- Legon, W.; Ai, L.; Bansal, P.; Mueller, J.K. Neuromodulation with single-element transcranial focused ultrasound in human thalamus. Hum. Brain Mapp. 2018, 39, 1995–2006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brinker, S.T.; Preiswerk, F.; White, P.J.; Mariano, T.Y.; McDannold, N.J.; Bubrick, E.J. Focused Ultrasound Platform for Investigating Therapeutic Neuromodulation Across the Human Hippocampus. Ultrasound Med. Biol. 2020, 46, 1270–1274. [Google Scholar] [CrossRef]
- Fomenko, A.; Chen, K.S.; Nankoo, J.F.; Saravanamuttu, J.; Wang, Y.; El-Baba, M.; Xia, X.; Seerala, S.S.; Hynynen, K.; Lozano, A.M.; et al. Systematic examination of low-intensity ultrasound parameters on human motor cortex excitability and behavior. eLife 2020, 9, e54497. [Google Scholar] [CrossRef]
- Badran, B.W.; Caulfield, K.A.; Stomberg-Firestein, S.; Summers, P.M.; Dowdle, L.T.; Savoca, M.; Li, X.; Austelle, C.W.; Short, E.B.; Borckardt, J.J.; et al. Sonication of the anterior thalamus with MRI-Guided transcranial focused ultrasound (tFUS) alters pain thresholds in healthy adults: A double-blind, sham-controlled study. Brain Stimul. 2020, 13, 1805–1812. [Google Scholar] [CrossRef]
- Sanguinetti, J.L.; Hameroff, S.; Smith, E.E.; Sato, T.; Daft, C.M.W.; Tyler, W.J.; Allen, J.J.B. Transcranial Focused Ultrasound to the Right Prefrontal Cortex Improves Mood and Alters Functional Connectivity in Humans. Front. Hum. Neurosci. 2020, 14, 52. [Google Scholar] [CrossRef] [Green Version]
- Braun, V.; Blackmore, J.; Cleveland, R.O.; Butler, C.R. Transcranial ultrasound stimulation in humans is associated with an auditory confound that can be effectively masked. Brain Stimul. 2020, 13, 1527–1534. [Google Scholar] [CrossRef]
- Stern, J.M.; Spivak, N.M.; Becerra, S.A.; Kuhn, T.P.; Korb, A.S.; Kronemyer, D.; Khanlou, N.; Reyes, S.D.; Monti, M.M.; Schnakers, C.; et al. Safety of focused ultrasound neuromodulation in humans with temporal lobe epilepsy. Brain Stimul. 2021, 14, 1022–1031. [Google Scholar] [CrossRef]
- Kamimura, H.A.S.; Conti, A.; Toschi, N.; Konofagou, E.E. Ultrasound neuromodulation: Mechanisms and the potential of multimodal stimulation for neuronal function assessment. Front. Phys. 2020, 8, 150. [Google Scholar] [CrossRef] [PubMed]
- Dauba, A.; Delalande, A.; Kamimura, H.A.S.; Conti, A.; Larrat, B.; Tsapis, N.; Novell, A. Recent Advances on Ultrasound Contrast Agents for Blood-Brain Barrier Opening with Focused Ultrasound. Pharmaceutics 2020, 12, 1125. [Google Scholar] [CrossRef] [PubMed]
- Padilla, F.; Ter Haar, G. Recommendations for Reporting Therapeutic Ultrasound Treatment Parameters. Ultrasound Med. Biol. 2022, 48, 1299–1308. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Balbi, M.; Blackmore, D.G.; Padmanabhan, P.; Götz, J. Ultrasound-Mediated Bioeffects in Senescent Mice and Alzheimer’s Mouse Models. Brain Sci. 2022, 12, 775. https://doi.org/10.3390/brainsci12060775
Balbi M, Blackmore DG, Padmanabhan P, Götz J. Ultrasound-Mediated Bioeffects in Senescent Mice and Alzheimer’s Mouse Models. Brain Sciences. 2022; 12(6):775. https://doi.org/10.3390/brainsci12060775
Chicago/Turabian StyleBalbi, Matilde, Daniel G. Blackmore, Pranesh Padmanabhan, and Jürgen Götz. 2022. "Ultrasound-Mediated Bioeffects in Senescent Mice and Alzheimer’s Mouse Models" Brain Sciences 12, no. 6: 775. https://doi.org/10.3390/brainsci12060775