
Low-Intensity Transcranial Ultrasound Stimulation: Mechanisms of Action and Rationale for Future Applications in Movement Disorders



Abstract
:1. Introduction
2. TUS Effects in Healthy Animals and Humans
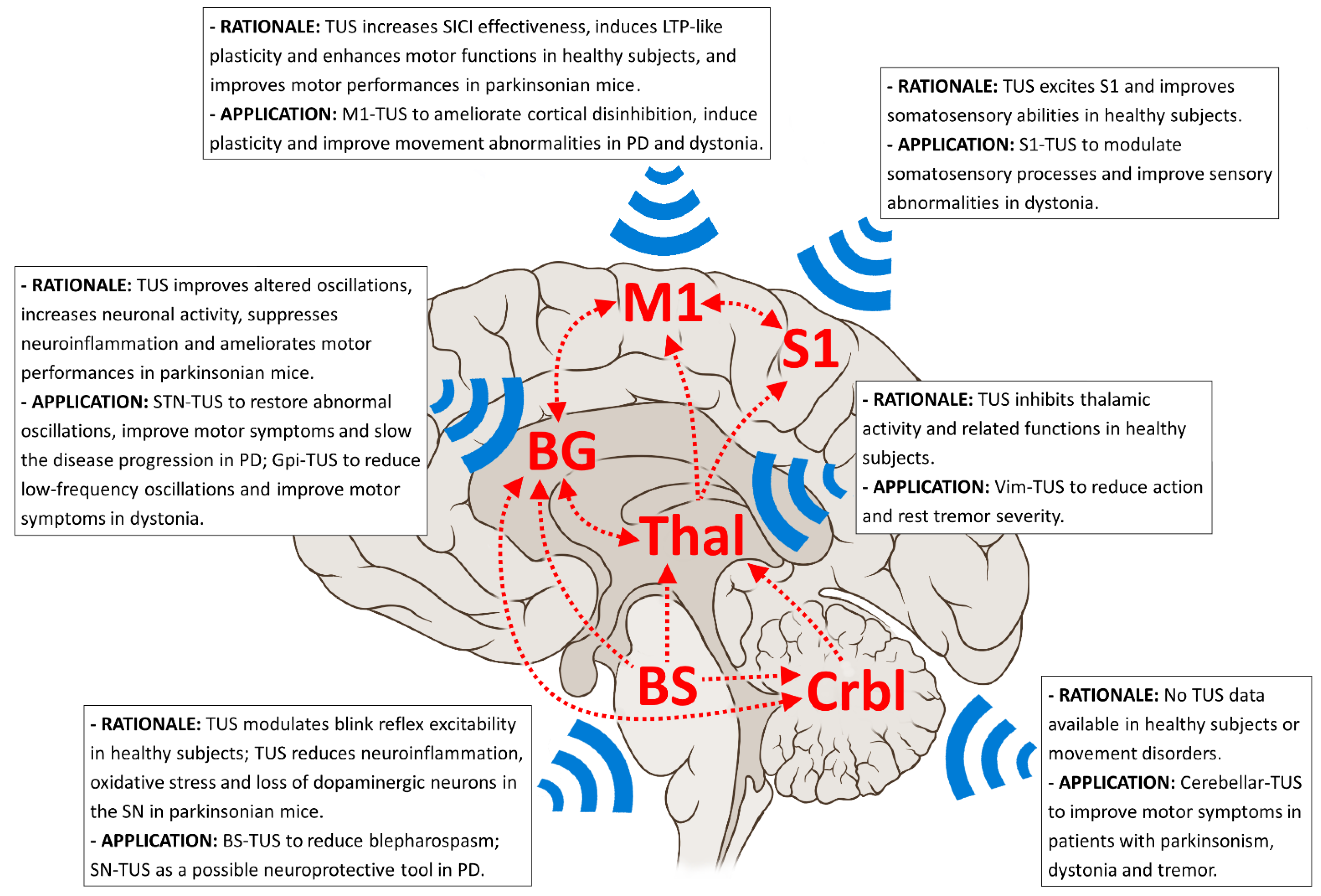
3. Possible Applications of TUS in Movement Disorders
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Blackmore, J.; Shrivastava, S.; Sallet, J.; Butler, C.R.; Cleveland, R.O. Ultrasound Neuromodulation: A Review of Results, Mechanisms and Safety. Ultrasound Med. Biol. 2019, 45, 1509–1536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darmani, G.; Bergmann, T.O.; Butts Pauly, K.; Caskey, C.F.; de Lecea, L.; Fomenko, A.; Fouragnan, E.; Legon, W.; Murphy, K.R.; Nandi, T.; et al. Non-Invasive Transcranial Ultrasound Stimulation for Neuromodulation. Clin. Neurophysiol. 2022, 135, 51–73. [Google Scholar] [CrossRef] [PubMed]
- Hameroff, S.; Trakas, M.; Duffield, C.; Annabi, E.; Gerace, M.B.; Boyle, P.; Lucas, A.; Amos, Q.; Buadu, A.; Badal, J.J. Transcranial Ultrasound (TUS) Effects on Mental States: A Pilot Study. Brain Stimul. 2013, 6, 409–415. [Google Scholar] [CrossRef] [PubMed]
- Di Biase, L.; Falato, E.; Di Lazzaro, V. Transcranial Focused Ultrasound (TFUS) and Transcranial Unfocused Ultrasound (TUS) Neuromodulation: From Theoretical Principles to Stimulation Practices. Front. Neurol. 2019, 10, 549. [Google Scholar] [CrossRef] [Green Version]
- Legon, W.; Ai, L.; Bansal, P.; Mueller, J.K. Neuromodulation with Single-Element Transcranial Focused Ultrasound in Human Thalamus. Hum. Brain Mapp. 2018, 39, 1995–2006. [Google Scholar] [CrossRef] [Green Version]
- Guerra, A.; Vicenzini, E.; Cioffi, E.; Colella, D.; Cannavacciuolo, A.; Pozzi, S.; Caccia, B.; Paparella, G.; Di Stefano, G.; Berardelli, A.; et al. Effects of Transcranial Ultrasound Stimulation on Trigeminal Blink Reflex Excitability. Brain Sci. 2021, 11, 645. [Google Scholar] [CrossRef]
- Monti, M.M.; Schnakers, C.; Korb, A.S.; Bystritsky, A.; Vespa, P.M. Non-Invasive Ultrasonic Thalamic Stimulation in Disorders of Consciousness after Severe Brain Injury: A First-in-Man Report. Brain Stimul. 2016, 9, 940–941. [Google Scholar] [CrossRef]
- Kim, T.; Park, C.; Chhatbar, P.Y.; Feld, J.; Mac Grory, B.; Nam, C.S.; Wang, P.; Chen, M.; Jiang, X.; Feng, W. Effect of Low Intensity Transcranial Ultrasound Stimulation on Neuromodulation in Animals and Humans: An Updated Systematic Review. Front. Neurosci. 2021, 15, 620863. [Google Scholar] [CrossRef]
- Krasovitski, B.; Frenkel, V.; Shoham, S.; Kimmel, E. Intramembrane Cavitation as a Unifying Mechanism for Ultrasound-Induced Bioeffects. Proc. Natl. Acad. Sci. USA 2011, 108, 3258–3263. [Google Scholar] [CrossRef] [Green Version]
- Plaksin, M.; Kimmel, E.; Shoham, S. Cell-Type-Selective Effects of Intramembrane Cavitation as a Unifying Theoretical Framework for Ultrasonic Neuromodulation. eNeuro 2016, 3, ENEURO.0136-15.2016. [Google Scholar] [CrossRef] [Green Version]
- Yoo, S.; Mittelstein, D.R.; Hurt, R.C.; Lacroix, J.; Shapiro, M.G. Focused Ultrasound Excites Cortical Neurons via Mechanosensitive Calcium Accumulation and Ion Channel Amplification. Nat. Commun. 2022, 13, 493. [Google Scholar] [CrossRef] [PubMed]
- Kubanek, J.; Shi, J.; Marsh, J.; Chen, D.; Deng, C.; Cui, J. Ultrasound Modulates Ion Channel Currents. Sci. Rep. 2016, 6, 24170. [Google Scholar] [CrossRef] [Green Version]
- Darrow, D.P.; O’Brien, P.; Richner, T.J.; Netoff, T.I.; Ebbini, E.S. Reversible Neuroinhibition by Focused Ultrasound Is Mediated by a Thermal Mechanism. Brain Stimul. 2019, 12, 1439–1447. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Pan, N.; Wang, Y.; Liu, C.; Hu, S. Transcranial Focused Ultrasound Neuromodulation: A Review of the Excitatory and Inhibitory Effects on Brain Activity in Human and Animals. Front. Hum. Neurosci. 2021, 15, 749162. [Google Scholar] [CrossRef] [PubMed]
- Pichardo, S.; Sin, V.W.; Hynynen, K. Multi-Frequency Characterization of the Speed of Sound and Attenuation Coefficient for Longitudinal Transmission of Freshly Excised Human Skulls. Phys. Med. Biol. 2011, 56, 219–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quadri, S.A.; Waqas, M.; Khan, I.; Khan, M.A.; Suriya, S.S.; Farooqui, M.; Fiani, B. High-Intensity Focused Ultrasound: Past, Present, and Future in Neurosurgery. Neurosurg. Focus 2018, 44, E16. [Google Scholar] [CrossRef] [Green Version]
- Moosa, S.; Martínez-Fernández, R.; Elias, W.J.; Del Alamo, M.; Eisenberg, H.M.; Fishman, P.S. The Role of High-Intensity Focused Ultrasound as a Symptomatic Treatment for Parkinson’s Disease. Mov. Disord. 2019, 34, 1243–1251. [Google Scholar] [CrossRef] [PubMed]
- Kubanek, J. Neuromodulation with Transcranial Focused Ultrasound. Neurosurg. Focus 2018, 44, E14. [Google Scholar] [CrossRef] [Green Version]
- Guerra, A.; Suppa, A.; Asci, F.; De Marco, G.; D’Onofrio, V.; Bologna, M.; Di Lazzaro, V.; Berardelli, A. LTD-like Plasticity of the Human Primary Motor Cortex Can Be Reversed by γ-TACS. Brain Stimul. 2019, 12, 1490–1499. [Google Scholar] [CrossRef]
- Guerra, A.; Asci, F.; D’Onofrio, V.; Sveva, V.; Bologna, M.; Fabbrini, G.; Berardelli, A.; Suppa, A. Enhancing Gamma Oscillations Restores Primary Motor Cortex Plasticity in Parkinson’s Disease. J. Neurosci. 2020, 40, 4788–4796. [Google Scholar] [CrossRef]
- Suppa, A.; Asci, F.; Guerra, A. Transcranial Magnetic Stimulation as a Tool to Induce and Explore Plasticity in Humans. Handb. Clin. Neurol. 2022, 184, 73–89. [Google Scholar] [CrossRef] [PubMed]
- Beisteiner, R.; Matt, E.; Fan, C.; Baldysiak, H.; Schönfeld, M.; Philippi Novak, T.; Amini, A.; Aslan, T.; Reinecke, R.; Lehrner, J.; et al. Transcranial Pulse Stimulation with Ultrasound in Alzheimer’s Disease-A New Navigated Focal Brain Therapy. Adv. Sci. 2020, 7, 1902583. [Google Scholar] [CrossRef] [Green Version]
- Jeong, H.; Im, J.J.; Park, J.-S.; Na, S.-H.; Lee, W.; Yoo, S.-S.; Song, I.-U.; Chung, Y.-A. A Pilot Clinical Study of Low-Intensity Transcranial Focused Ultrasound in Alzheimer’s Disease. Ultrasonography 2021, 40, 512–519. [Google Scholar] [CrossRef] [PubMed]
- Jeong, H.; Song, I.-U.; Chung, Y.-A.; Park, J.-S.; Na, S.-H.; Im, J.J.; Bikson, M.; Lee, W.; Yoo, S.-S. Short-Term Efficacy of Transcranial Focused Ultrasound to the Hippocampus in Alzheimer’s Disease: A Preliminary Study. J. Pers. Med. 2022, 12, 250. [Google Scholar] [CrossRef] [PubMed]
- Stern, J.M.; Spivak, N.M.; Becerra, S.A.; Kuhn, T.P.; Korb, A.S.; Kronemyer, D.; Khanlou, N.; Reyes, S.D.; Monti, M.M.; Schnakers, C.; et al. Safety of Focused Ultrasound Neuromodulation in Humans with Temporal Lobe Epilepsy. Brain Stimul. 2021, 14, 1022–1031. [Google Scholar] [CrossRef]
- Lee, C.-C.; Chou, C.-C.; Hsiao, F.-J.; Chen, Y.-H.; Lin, C.-F.; Chen, C.-J.; Peng, S.-J.; Liu, H.-L.; Yu, H.-Y. Pilot Study of Focused Ultrasound for Drug-Resistant Epilepsy. Epilepsia 2022, 63, 162–175. [Google Scholar] [CrossRef]
- Chen, X.; Wang, D.; Zhang, L.; Yao, H.; Zhu, H.; Zhao, N.; Peng, X.; Yang, K. Neuroprotective Effect of Low-Intensity Pulsed Ultrasound on the Mouse MPTP/MPP+ Model of Dopaminergic Neuron Injury. Ultrasound Med. Biol. 2021, 47, 2321–2330. [Google Scholar] [CrossRef]
- Wang, Z.; Yan, J.; Wang, X.; Yuan, Y.; Li, X. Transcranial Ultrasound Stimulation Directly Influences the Cortical Excitability of the Motor Cortex in Parkinsonian Mice. Mov. Disord. 2020, 35, 693–698. [Google Scholar] [CrossRef]
- Xu, T.; Lu, X.; Peng, D.; Wang, G.; Chen, C.; Liu, W.; Wu, W.; Mason, T.J. Ultrasonic Stimulation of the Brain to Enhance the Release of Dopamine—A Potential Novel Treatment for Parkinson’s Disease. Ultrason. Sonochem. 2020, 63, 104955. [Google Scholar] [CrossRef]
- Zhou, H.; Meng, L.; Xia, X.; Lin, Z.; Zhou, W.; Pang, N.; Bian, T.; Yuan, T.; Niu, L.; Zheng, H. Transcranial Ultrasound Stimulation Suppresses Neuroinflammation in a Chronic Mouse Model of Parkinson’s Disease. IEEE Trans. Biomed. Eng. 2021, 68, 3375–3387. [Google Scholar] [CrossRef]
- Zhou, H.; Niu, L.; Xia, X.; Lin, Z.; Liu, X.; Su, M.; Guo, R.; Meng, L.; Zheng, H. Wearable Ultrasound Improves Motor Function in an MPTP Mouse Model of Parkinson’s Disease. IEEE Trans. Biomed. Eng. 2019, 66, 3006–3013. [Google Scholar] [CrossRef]
- Bologna, M.; Valls-Solé, J.; Kamble, N.; Pal, P.K.; Conte, A.; Guerra, A.; Belvisi, D.; Berardelli, A. Dystonia, Chorea, and Other Dyskinesia. Clin. Neurophysiol. 2022; in press. [Google Scholar]
- Bologna, M.; Paparella, G.; Fasano, A.; Hallett, M.; Berardelli, A. Evolving Concepts on Bradykinesia. Brain 2020, 143, 727–750. [Google Scholar] [CrossRef] [PubMed]
- Wichmann, T. Changing Views of the Pathophysiology of Parkinsonism. Mov. Disord. 2019, 34, 1130–1143. [Google Scholar] [CrossRef]
- Bologna, M.; Berardelli, A. Cerebellum: An Explanation for Dystonia? Cerebellum Ataxias 2017, 4, 6. [Google Scholar] [CrossRef] [PubMed]
- Jinnah, H.A.; Neychev, V.; Hess, E.J. The Anatomical Basis for Dystonia: The Motor Network Model. Tremor Other Hyperkinet Mov. 2017, 7, 506. [Google Scholar] [CrossRef]
- Dirkx, M.F.; Bologna, M. The Pathophysiology of Parkinson’s Disease Tremor. J. Neurol. Sci. 2022, 435, 120196. [Google Scholar] [CrossRef]
- Haubenberger, D.; Hallett, M. Essential Tremor. N. Engl. J. Med. 2018, 378, 1802–1810. [Google Scholar] [CrossRef]
- Tufail, Y.; Matyushov, A.; Baldwin, N.; Tauchmann, M.L.; Georges, J.; Yoshihiro, A.; Tillery, S.I.H.; Tyler, W.J. Transcranial Pulsed Ultrasound Stimulates Intact Brain Circuits. Neuron 2010, 66, 681–694. [Google Scholar] [CrossRef] [Green Version]
- Lee, W.; Croce, P.; Margolin, R.W.; Cammalleri, A.; Yoon, K.; Yoo, S.-S. Transcranial Focused Ultrasound Stimulation of Motor Cortical Areas in Freely-Moving Awake Rats. BMC Neurosci. 2018, 19, 57. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.; Kim, S.; Sim, N.S.; Pasquinelli, C.; Thielscher, A.; Lee, J.H.; Lee, H.J. Miniature Ultrasound Ring Array Transducers for Transcranial Ultrasound Neuromodulation of Freely-Moving Small Animals. Brain Stimul. 2019, 12, 251–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, Z.; Li, D.; Feng, Y.; Xu, T.; Wu, S.; Li, Y.; Bouakaz, A.; Wan, M.; Zhang, S. Enhanced Neuronal Activity in Mouse Motor Cortex with Microbubbles’ Oscillations by Transcranial Focused Ultrasound Stimulation. Ultrason. Sonochem. 2019, 59, 104745. [Google Scholar] [CrossRef] [PubMed]
- Cui, Z.; Li, D.; Xu, S.; Xu, T.; Wu, S.; Bouakaz, A.; Wan, M.; Zhang, S. Effect of Scattered Pressures from Oscillating Microbubbles on Neuronal Activity in Mouse Brain under Transcranial Focused Ultrasound Stimulation. Ultrason. Sonochem. 2020, 63, 104935. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhou, X.; Cui, D.; Liu, R.; Tan, R.; Wang, X.; Liu, Z.; Yin, T. Comparative Study of Transcranial Magneto-Acoustic Stimulation and Transcranial Ultrasound Stimulation of Motor Cortex. Front. Behav. Neurosci. 2019, 13, 241. [Google Scholar] [CrossRef]
- King, R.L.; Brown, J.R.; Newsome, W.T.; Pauly, K.B. Effective Parameters for Ultrasound-Induced in Vivo Neurostimulation. Ultrasound Med. Biol. 2013, 39, 312–331. [Google Scholar] [CrossRef]
- Wang, X.; Yan, J.; Wang, Z.; Li, X.; Yuan, Y. Neuromodulation Effects of Ultrasound Stimulation Under Different Parameters on Mouse Motor Cortex. IEEE Trans. Biomed. Eng. 2020, 67, 291–297. [Google Scholar] [CrossRef]
- Wang, Y.; Xie, P.; Zhou, S.; Wang, X.; Yuan, Y. Low-Intensity Pulsed Ultrasound Modulates Multi-Frequency Band Phase Synchronization between LFPs and EMG in Mice. J. Neural Eng. 2019, 16, 026036. [Google Scholar] [CrossRef]
- Yuan, Y.; Wang, Z.; Liu, M.; Shoham, S. Cortical Hemodynamic Responses Induced by Low-Intensity Transcranial Ultrasound Stimulation of Mouse Cortex. Neuroimage 2020, 211, 116597. [Google Scholar] [CrossRef]
- Gibson, B.C.; Sanguinetti, J.L.; Badran, B.W.; Yu, A.B.; Klein, E.P.; Abbott, C.C.; Hansberger, J.T.; Clark, V.P. Increased Excitability Induced in the Primary Motor Cortex by Transcranial Ultrasound Stimulation. Front. Neurol. 2018, 9, 1007. [Google Scholar] [CrossRef] [Green Version]
- Fomenko, A.; Chen, K.-H.S.; Nankoo, J.-F.; Saravanamuttu, J.; Wang, Y.; El-Baba, M.; Xia, X.; Seerala, S.S.; Hynynen, K.; Lozano, A.M.; et al. Systematic Examination of Low-Intensity Ultrasound Parameters on Human Motor Cortex Excitability and Behavior. eLife 2020, 9, e54497. [Google Scholar] [CrossRef]
- Zhang, Y.; Ren, L.; Liu, K.; Tong, S.; Yuan, T.-F.; Sun, J. Transcranial Ultrasound Stimulation of the Human Motor Cortex. iScience 2021, 24, 103429. [Google Scholar] [CrossRef] [PubMed]
- Zeng, K.; Darmani, G.; Fomenko, A.; Xia, X.; Tran, S.; Nankoo, J.-F.; Shamli Oghli, Y.; Wang, Y.; Lozano, A.M.; Chen, R. Induction of Human Motor Cortex Plasticity by Theta Burst Transcranial Ultrasound Stimulation. Ann. Neurol. 2022, 91, 238–252. [Google Scholar] [CrossRef] [PubMed]
- Legon, W.; Bansal, P.; Tyshynsky, R.; Ai, L.; Mueller, J.K. Transcranial Focused Ultrasound Neuromodulation of the Human Primary Motor Cortex. Sci. Rep. 2018, 8, 10007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ai, L.; Bansal, P.; Mueller, J.K.; Legon, W. Effects of Transcranial Focused Ultrasound on Human Primary Motor Cortex Using 7T FMRI: A Pilot Study. BMC Neurosci. 2018, 19, 56. [Google Scholar] [CrossRef]
- Kujirai, T.; Caramia, M.D.; Rothwell, J.C.; Day, B.L.; Thompson, P.D.; Ferbert, A.; Wroe, S.; Asselman, P.; Marsden, C.D. Corticocortical Inhibition in Human Motor Cortex. J. Physiol. 1993, 471, 501–519. [Google Scholar] [CrossRef]
- Ziemann, U.; Reis, J.; Schwenkreis, P.; Rosanova, M.; Strafella, A.; Badawy, R.; Müller-Dahlhaus, F. TMS and Drugs Revisited 2014. Clin. Neurophysiol. 2015, 126, 1847–1868. [Google Scholar] [CrossRef]
- Huang, Y.-Z.; Edwards, M.J.; Rounis, E.; Bhatia, K.P.; Rothwell, J.C. Theta Burst Stimulation of the Human Motor Cortex. Neuron 2005, 45, 201–206. [Google Scholar] [CrossRef] [Green Version]
- Verhagen, L.; Gallea, C.; Folloni, D.; Constans, C.; Jensen, D.E.; Ahnine, H.; Roumazeilles, L.; Santin, M.; Ahmed, B.; Lehericy, S.; et al. Offline Impact of Transcranial Focused Ultrasound on Cortical Activation in Primates. eLife 2019, 8, e40541. [Google Scholar] [CrossRef]
- Moore, M.E.; Loft, J.M.; Clegern, W.C.; Wisor, J.P. Manipulating Neuronal Activity in the Mouse Brain with Ultrasound: A Comparison with Optogenetic Activation of the Cerebral Cortex. Neurosci. Lett. 2015, 604, 183–187. [Google Scholar] [CrossRef]
- Fisher, J.A.N.; Gumenchuk, I. Low-Intensity Focused Ultrasound Alters the Latency and Spatial Patterns of Sensory-Evoked Cortical Responses in Vivo. J. Neural Eng. 2018, 15, 035004. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Qiu, W.; Zhang, Z.; Jiang, Q.; Su, M.; Cai, R.; Li, Y.; Cai, F.; Deng, Z.; Xu, D.; et al. Noninvasive Ultrasonic Neuromodulation in Freely Moving Mice. IEEE Trans. Biomed. Eng. 2019, 66, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.; Anguluan, E.; Youn, S.; Kim, J.; Hwang, J.Y.; Kim, J.G. Non-Invasive Measurement of Hemodynamic Change during 8 MHz Transcranial Focused Ultrasound Stimulation Using near-Infrared Spectroscopy. BMC Neurosci. 2019, 20, 12. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.; Kim, H.; Jung, Y.; Song, I.-U.; Chung, Y.A.; Yoo, S.-S. Image-Guided Transcranial Focused Ultrasound Stimulates Human Primary Somatosensory Cortex. Sci. Rep. 2015, 5, 8743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, W.; Chung, Y.A.; Jung, Y.; Song, I.-U.; Yoo, S.-S. Simultaneous Acoustic Stimulation of Human Primary and Secondary Somatosensory Cortices Using Transcranial Focused Ultrasound. BMC Neurosci. 2016, 17, 68. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Yu, K.; Niu, X.; He, B. Transcranial Focused Ultrasound Enhances Sensory Discrimination Capability through Somatosensory Cortical Excitation. Ultrasound Med. Biol. 2021, 47, 1356–1366. [Google Scholar] [CrossRef]
- Legon, W.; Sato, T.F.; Opitz, A.; Mueller, J.; Barbour, A.; Williams, A.; Tyler, W.J. Transcranial Focused Ultrasound Modulates the Activity of Primary Somatosensory Cortex in Humans. Nat. Neurosci. 2014, 17, 322–329. [Google Scholar] [CrossRef]
- Yoo, S.-S.; Yoon, K.; Croce, P.; Cammalleri, A.; Margolin, R.W.; Lee, W. Focused Ultrasound Brain Stimulation to Anesthetized Rats Induces Long-Term Changes in Somatosensory Evoked Potentials. Int. J. Imaging Syst. Technol. 2018, 28, 106–112. [Google Scholar] [CrossRef]
- Mueller, J.; Legon, W.; Opitz, A.; Sato, T.F.; Tyler, W.J. Transcranial Focused Ultrasound Modulates Intrinsic and Evoked EEG Dynamics. Brain Stimul. 2014, 7, 900–908. [Google Scholar] [CrossRef] [Green Version]
- Dallapiazza, R.F.; Timbie, K.F.; Holmberg, S.; Gatesman, J.; Lopes, M.B.; Price, R.J.; Miller, G.W.; Elias, W.J. Noninvasive Neuromodulation and Thalamic Mapping with Low-Intensity Focused Ultrasound. J. Neurosurg. 2018, 128, 875–884. [Google Scholar] [CrossRef]
- Yoon, K.; Lee, W.; Lee, J.E.; Xu, L.; Croce, P.; Foley, L.; Yoo, S.-S. Effects of Sonication Parameters on Transcranial Focused Ultrasound Brain Stimulation in an Ovine Model. PLoS ONE 2019, 14, e0224311. [Google Scholar] [CrossRef] [Green Version]
- Badran, B.W.; Caulfield, K.A.; Stomberg-Firestein, S.; Summers, P.M.; Dowdle, L.T.; Savoca, M.; Li, X.; Austelle, C.W.; Short, E.B.; Borckardt, J.J.; et al. Sonication of the Anterior Thalamus with MRI-Guided Transcranial Focused Ultrasound (TFUS) Alters Pain Thresholds in Healthy Adults: A Double-Blind, Sham-Controlled Study. Brain Stimul. 2020, 13, 1805–1812. [Google Scholar] [CrossRef] [PubMed]
- Folloni, D.; Verhagen, L.; Mars, R.B.; Fouragnan, E.; Constans, C.; Aubry, J.-F.; Rushworth, M.F.S.; Sallet, J. Manipulation of Subcortical and Deep Cortical Activity in the Primate Brain Using Transcranial Focused Ultrasound Stimulation. Neuron 2019, 101, 1109–1116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, H.; Yuan, Y.; Wang, X.; Li, X. Closed-Loop Transcranial Ultrasound Stimulation for Real-Time Non-Invasive Neuromodulation in Vivo. Front. Neurosci. 2020, 14, 445. [Google Scholar] [CrossRef] [PubMed]
- Mohammadjavadi, M.; Ye, P.P.; Xia, A.; Brown, J.; Popelka, G.; Pauly, K.B. Elimination of Peripheral Auditory Pathway Activation Does Not Affect Motor Responses from Ultrasound Neuromodulation. Brain Stimul. 2019, 12, 901–910. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Niu, L.; Meng, W.; Lin, Z.; Zou, J.; Bian, T.; Huang, X.; Zhou, H.; Meng, L.; Xie, P.; et al. Ultrasound Stimulation of Periaqueductal Gray Induces Defensive Behaviors. IEEE Trans. Ultrason. Ferroelectr. Freq. Control 2021, 68, 38–45. [Google Scholar] [CrossRef]
- Bologna, M.; Guerra, A.; Paparella, G.; Giordo, L.; Alunni Fegatelli, D.; Vestri, A.R.; Rothwell, J.C.; Berardelli, A. Neurophysiological Correlates of Bradykinesia in Parkinson’s Disease. Brain 2018, 141, 2432–2444. [Google Scholar] [CrossRef]
- Guerra, A.; Colella, D.; Giangrosso, M.; Cannavacciuolo, A.; Paparella, G.; Fabbrini, G.; Suppa, A.; Berardelli, A.; Bologna, M. Driving Motor Cortex Oscillations Modulates Bradykinesia in Parkinson’s Disease. Brain 2022, 145, 224–236. [Google Scholar] [CrossRef]
- Ammann, C.; Dileone, M.; Pagge, C.; Catanzaro, V.; Mata-Marín, D.; Hernández-Fernández, F.; Monje, M.H.G.; Sánchez-Ferro, Á.; Fernández-Rodríguez, B.; Gasca-Salas, C.; et al. Cortical Disinhibition in Parkinson’s Disease. Brain 2020, 143, 3408–3421. [Google Scholar] [CrossRef]
- Berardelli, A.; Abbruzzese, G.; Chen, R.; Orth, M.; Ridding, M.C.; Stinear, C.; Suppa, A.; Trompetto, C.; Thompson, P.D. Consensus Paper on Short-Interval Intracortical Inhibition and Other Transcranial Magnetic Stimulation Intracortical Paradigms in Movement Disorders. Brain Stimul. 2008, 1, 183–191. [Google Scholar] [CrossRef]
- Ridding, M.C.; Inzelberg, R.; Rothwell, J.C. Changes in Excitability of Motor Cortical Circuitry in Patients with Parkinson’s Disease. Ann. Neurol. 1995, 37, 181–188. [Google Scholar] [CrossRef]
- Beck, S.; Richardson, S.P.; Shamim, E.A.; Dang, N.; Schubert, M.; Hallett, M. Short Intracortical and Surround Inhibition Are Selectively Reduced during Movement Initiation in Focal Hand Dystonia. J. Neurosci. 2008, 28, 10363–10369. [Google Scholar] [CrossRef] [Green Version]
- Kanovský, P.; Bares, M.; Streitová, H.; Klajblová, H.; Daniel, P.; Rektor, I. Abnormalities of Cortical Excitability and Cortical Inhibition in Cervical Dystonia Evidence from Somatosensory Evoked Potentials and Paired Transcranial Magnetic Stimulation Recordings. J. Neurol. 2003, 250, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Ridding, M.C.; Sheean, G.; Rothwell, J.C.; Inzelberg, R.; Kujirai, T. Changes in the Balance between Motor Cortical Excitation and Inhibition in Focal, Task Specific Dystonia. J. Neurol. Neurosurg. Psychiatry 1995, 59, 493–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stinear, C.M.; Byblow, W.D. Impaired Modulation of Intracortical Inhibition in Focal Hand Dystonia. Cereb Cortex 2004, 14, 555–561. [Google Scholar] [CrossRef] [Green Version]
- Hallett, M. Neurophysiology of Dystonia: The Role of Inhibition. Neurobiol. Dis. 2011, 42, 177–184. [Google Scholar] [CrossRef] [Green Version]
- Udupa, K.; Chen, R. Motor Cortical Plasticity in Parkinson’s Disease. Front. Neurol. 2013, 4, 128. [Google Scholar] [CrossRef] [Green Version]
- Quartarone, A.; Hallett, M. Emerging Concepts in the Physiological Basis of Dystonia. Mov. Disord. 2013, 28, 958–967. [Google Scholar] [CrossRef]
- Kojovic, M.; Caronni, A.; Bologna, M.; Rothwell, J.C.; Bhatia, K.P.; Edwards, M.J. Botulinum Toxin Injections Reduce Associative Plasticity in Patients with Primary Dystonia. Mov. Disord. 2011, 26, 1282–1289. [Google Scholar] [CrossRef]
- Quartarone, A.; Morgante, F.; Sant’angelo, A.; Rizzo, V.; Bagnato, S.; Terranova, C.; Siebner, H.R.; Berardelli, A.; Girlanda, P. Abnormal Plasticity of Sensorimotor Circuits Extends beyond the Affected Body Part in Focal Dystonia. J. Neurol. Neurosurg. Psychiatry 2008, 79, 985–990. [Google Scholar] [CrossRef]
- Latorre, A.; Rocchi, L.; Berardelli, A.; Bhatia, K.P.; Rothwell, J.C. The Use of Transcranial Magnetic Stimulation as a Treatment for Movement Disorders: A Critical Review. Mov. Disord. 2019, 34, 769–782. [Google Scholar] [CrossRef]
- Wu, T.; Long, X.; Wang, L.; Hallett, M.; Zang, Y.; Li, K.; Chan, P. Functional Connectivity of Cortical Motor Areas in the Resting State in Parkinson’s Disease. Hum. Brain Mapp. 2011, 32, 1443–1457. [Google Scholar] [CrossRef]
- Pollok, B.; Kamp, D.; Butz, M.; Wojtecki, L.; Timmermann, L.; Südmeyer, M.; Krause, V.; Schnitzler, A. Increased SMA-M1 Coherence in Parkinson’s Disease—Pathophysiology or Compensation? Exp. Neurol. 2013, 247, 178–181. [Google Scholar] [CrossRef]
- Tessitore, A.; Cirillo, M.; De Micco, R. Functional Connectivity Signatures of Parkinson’s Disease. J. Parkinsons Dis. 2019, 9, 637–652. [Google Scholar] [CrossRef] [Green Version]
- Stamelou, M.; Edwards, M.J.; Hallett, M.; Bhatia, K.P. The Non-Motor Syndrome of Primary Dystonia: Clinical and Pathophysiological Implications. Brain 2012, 135, 1668–1681. [Google Scholar] [CrossRef]
- Tisch, S.; Limousin, P. Neurophysiological Insights in Dystonia and Its Response to Deep Brain Stimulation Treatment. Exp. Brain Res. 2020, 238, 1645–1657. [Google Scholar] [CrossRef]
- Bara-Jimenez, W.; Shelton, P.; Hallett, M. Spatial Discrimination Is Abnormal in Focal Hand Dystonia. Neurology 2000, 55, 1869–1873. [Google Scholar] [CrossRef]
- Fiorio, M.; Tinazzi, M.; Bertolasi, L.; Aglioti, S.M. Temporal Processing of Visuotactile and Tactile Stimuli in Writer’s Cramp. Ann. Neurol. 2003, 53, 630–635. [Google Scholar] [CrossRef]
- Antelmi, E.; Erro, R.; Rocchi, L.; Liguori, R.; Tinazzi, M.; Di Stasio, F.; Berardelli, A.; Rothwell, J.C.; Bhatia, K.P. Neurophysiological Correlates of Abnormal Somatosensory Temporal Discrimination in Dystonia. Mov. Disord. 2017, 32, 141–148. [Google Scholar] [CrossRef] [Green Version]
- Kühn, A.A.; Tsui, A.; Aziz, T.; Ray, N.; Brücke, C.; Kupsch, A.; Schneider, G.-H.; Brown, P. Pathological Synchronisation in the Subthalamic Nucleus of Patients with Parkinson’s Disease Relates to Both Bradykinesia and Rigidity. Exp. Neurol. 2009, 215, 380–387. [Google Scholar] [CrossRef]
- Lofredi, R.; Tan, H.; Neumann, W.-J.; Yeh, C.-H.; Schneider, G.-H.; Kühn, A.A.; Brown, P. Beta Bursts during Continuous Movements Accompany the Velocity Decrement in Parkinson’s Disease Patients. Neurobiol. Dis. 2019, 127, 462–471. [Google Scholar] [CrossRef]
- Lofredi, R.; Neumann, W.-J.; Bock, A.; Horn, A.; Huebl, J.; Siegert, S.; Schneider, G.-H.; Krauss, J.K.; Kühn, A.A. Dopamine-Dependent Scaling of Subthalamic Gamma Bursts with Movement Velocity in Patients with Parkinson’s Disease. eLife 2018, 7, e31895. [Google Scholar] [CrossRef]
- Oswal, A.; Brown, P.; Litvak, V. Synchronized Neural Oscillations and the Pathophysiology of Parkinson’s Disease. Curr. Opin. Neurol. 2013, 26, 662–670. [Google Scholar] [CrossRef] [Green Version]
- Tan, H.; Pogosyan, A.; Anzak, A.; Foltynie, T.; Limousin, P.; Zrinzo, L.; Ashkan, K.; Bogdanovic, M.; Green, A.L.; Aziz, T.; et al. Frequency Specific Activity in Subthalamic Nucleus Correlates with Hand Bradykinesia in Parkinson’s Disease. Exp. Neurol. 2013, 240, 122–129. [Google Scholar] [CrossRef] [Green Version]
- Silberstein, P.; Kühn, A.A.; Kupsch, A.; Trottenberg, T.; Krauss, J.K.; Wöhrle, J.C.; Mazzone, P.; Insola, A.; Di Lazzaro, V.; Oliviero, A.; et al. Patterning of Globus Pallidus Local Field Potentials Differs between Parkinson’s Disease and Dystonia. Brain 2003, 126, 2597–2608. [Google Scholar] [CrossRef]
- Chen, C.C.; Kühn, A.A.; Hoffmann, K.-T.; Kupsch, A.; Schneider, G.-H.; Trottenberg, T.; Krauss, J.K.; Wöhrle, J.C.; Bardinet, E.; Yelnik, J.; et al. Oscillatory Pallidal Local Field Potential Activity Correlates with Involuntary EMG in Dystonia. Neurology 2006, 66, 418–420. [Google Scholar] [CrossRef]
- Neumann, W.-J.; Horn, A.; Ewert, S.; Huebl, J.; Brücke, C.; Slentz, C.; Schneider, G.-H.; Kühn, A.A. A Localized Pallidal Physiomarker in Cervical Dystonia. Ann. Neurol. 2017, 82, 912–924. [Google Scholar] [CrossRef]
- Moro, E.; LeReun, C.; Krauss, J.K.; Albanese, A.; Lin, J.-P.; Walleser Autiero, S.; Brionne, T.C.; Vidailhet, M. Efficacy of Pallidal Stimulation in Isolated Dystonia: A Systematic Review and Meta-Analysis. Eur. J. Neurol. 2017, 24, 552–560. [Google Scholar] [CrossRef]
- Pietracupa, S.; Bologna, M.; Tommasin, S.; Berardelli, A.; Pantano, P. The Contribution of Neuroimaging to the Understanding of Essential Tremor Pathophysiology: A Systematic Review. Cerebellum 2021. [Google Scholar] [CrossRef]
- Helmich, R.C.; Hallett, M.; Deuschl, G.; Toni, I.; Bloem, B.R. Cerebral Causes and Consequences of Parkinsonian Resting Tremor: A Tale of Two Circuits? Brain 2012, 135, 3206–3226. [Google Scholar] [CrossRef] [Green Version]
- Madelein van der Stouwe, A.M.; Nieuwhof, F.; Helmich, R.C. Tremor Pathophysiology: Lessons from Neuroimaging. Curr. Opin. Neurol. 2020, 33, 474–481. [Google Scholar] [CrossRef]
- Cury, R.G.; Fraix, V.; Castrioto, A.; Pérez Fernández, M.A.; Krack, P.; Chabardes, S.; Seigneuret, E.; Alho, E.J.L.; Benabid, A.-L.; Moro, E. Thalamic Deep Brain Stimulation for Tremor in Parkinson Disease, Essential Tremor, and Dystonia. Neurology 2017, 89, 1416–1423. [Google Scholar] [CrossRef] [PubMed]
- Dallapiazza, R.F.; Lee, D.J.; De Vloo, P.; Fomenko, A.; Hamani, C.; Hodaie, M.; Kalia, S.K.; Fasano, A.; Lozano, A.M. Outcomes from Stereotactic Surgery for Essential Tremor. J. Neurol. Neurosurg. Psychiatry 2019, 90, 474–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rohani, M.; Fasano, A. Focused Ultrasound for Essential Tremor: Review of the Evidence and Discussion of Current Hurdles. Tremor Other Hyperkinet. Mov. 2017, 7, 462. [Google Scholar] [CrossRef]
- Dhima, K.; Biars, J.; Kondylis, E.; Nagel, S.; Yu, X.X.; Floden, D.P. Neuropsychological Outcomes after Thalamic Deep Brain Stimulation for Essential Tremor. Parkinsonism Relat. Disord. 2021, 92, 88–93. [Google Scholar] [CrossRef]
- Bologna, M.; Berardelli, A. The Cerebellum and Dystonia. Handb. Clin. Neurol. 2018, 155, 259–272. [Google Scholar] [CrossRef]
- Pan, M.-K.; Kuo, S.-H. Essential Tremor: Clinical Perspectives and Pathophysiology. J. Neurol. Sci. 2022, 435, 120198. [Google Scholar] [CrossRef]
- Mavroudis, I.; Kazis, D.; Petridis, F.; Chatzikonstantinou, S.; Karantali, E.; Njau, S.N.; Costa, V.; Ciobica, A.; Trus, C.; Balmus, I.M.; et al. Morphological and Morphometric Changes in the Purkinje Cells of Patients with Essential Tremor. Exp. Ther. Med. 2022, 23, 167. [Google Scholar] [CrossRef]
- Kojovic, M.; Pareés, I.; Kassavetis, P.; Palomar, F.J.; Mir, P.; Teo, J.T.; Cordivari, C.; Rothwell, J.C.; Bhatia, K.P.; Edwards, M.J. Secondary and Primary Dystonia: Pathophysiological Differences. Brain 2013, 136, 2038–2049. [Google Scholar] [CrossRef] [Green Version]
- Popa, T.; Milani, P.; Richard, A.; Hubsch, C.; Brochard, V.; Tranchant, C.; Sadnicka, A.; Rothwell, J.; Vidailhet, M.; Meunier, S.; et al. The Neurophysiological Features of Myoclonus-Dystonia and Differentiation from Other Dystonias. JAMA Neurol. 2014, 71, 612–619. [Google Scholar] [CrossRef] [Green Version]
- Sadnicka, A.; Teo, J.T.; Kojovic, M.; Pareés, I.; Saifee, T.A.; Kassavetis, P.; Schwingenschuh, P.; Katschnig-Winter, P.; Stamelou, M.; Mencacci, N.E.; et al. All in the Blink of an Eye: New Insight into Cerebellar and Brainstem Function in DYT1 and DYT6 Dystonia. Eur. J. Neurol. 2015, 22, 762–767. [Google Scholar] [CrossRef]
- Brighina, F.; Romano, M.; Giglia, G.; Saia, V.; Puma, A.; Giglia, F.; Fierro, B. Effects of Cerebellar TMS on Motor Cortex of Patients with Focal Dystonia: A Preliminary Report. Exp. Brain Res. 2009, 192, 651–656. [Google Scholar] [CrossRef] [PubMed]
- Pauly, M.G.; Steinmeier, A.; Bolte, C.; Hamami, F.; Tzvi, E.; Münchau, A.; Bäumer, T.; Weissbach, A. Cerebellar RTMS and PAS Effectively Induce Cerebellar Plasticity. Sci. Rep. 2021, 11, 3070. [Google Scholar] [CrossRef] [PubMed]
- Spampinato, D.; Avci, E.; Rothwell, J.; Rocchi, L. Frequency-Dependent Modulation of Cerebellar Excitability during the Application of Non-Invasive Alternating Current Stimulation. Brain Stimul. 2021, 14, 277–283. [Google Scholar] [CrossRef] [PubMed]
- Maiorana, N.; Guidetti, M.; Dini, M.; Priori, A.; Ferrucci, R. Cerebellar TDCS as Therapy for Cerebellar Ataxias. Cerebellum 2022. [Google Scholar] [CrossRef]


{kind=link}
| Reference | Study Subjects | Target area | TUS Parameters | Stimulation Protocol | Key Findings | Significance |
|---|---|---|---|---|---|---|
| Zhou et al., 2019 [31] | Parkinsonian rats | M1 | 800 kHz, 100 Hz PRF, 10% DC, 6 s SD, 10 s ISI, 760 mW/cm2 ISPPA | 40 min/day for 7 days | - Improved locomotor activity, movement balance and bradykinesia - Increased c-Fos + cells in M1 and total SOD and GPx activity in the striatum | M1-TUS ameliorates motor symptoms and exerts antioxidative effects in PD |
| Wang et al., 2020 [28] | Parkinsonian mice | STN | 500 kHz, 1 kHz PRF, 5% DC, 50 ms SD, 1 s ISI, 5.1 W/cm2 ISPPA | 5 min total stimulation time | - Decreased beta power - Decreased beta-gamma and beta-ripple frequency PAC | STN-TUS improves the typical pattern of altered oscillatory activity in PD |
| Xu et al., 2020 [29] | Parkinsonian mice | Whole brain | 1 MHz, continuous mode DC, 0.3 W/cm2 ISPPA (unfocused TUS) | 5 min/day for 10 days | - Improved locomotion - Increased dopamine levels in the striatum | TUS ameliorates motor symptoms and may induce dopaminergic neurons regeneration |
| Zhou et al., 2021 [30] | Parkinsonian mice | STN | 3.8 MHz, 1 kHz PRF, 50% DC, 1 s SD, 4 s ISI, 430 mW/cm2 ISPTA | 2 sessions per week for 5 weeks | - Improved movement coordination, balance, and bradykinesia - Increased c-Fos expression in the STN - Downregulation of proinflammatory signaling and reduced activation of microglia and astrocytes in the SN and the striatum | STN-TUS improves motor functions and suppresses the neuroinflammation response in basal ganglia in PD |
| Chen et al., 2021 [27] | Parkinsonian mice | Whole brain | 1 MHz, 1 kHz PRF, 20% DC, ≈120 mW/cm2 ISPTA (unfocused TUS) | 10 min/day for 5 days | - Improved movement and balance - Decreased loss of TH + neurons in the SN - Improved intracellular oxidative stress and mitochondrial dysfunction | TUS improves motor dysfunctions and may have neuroprotective effects in PD |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guerra, A.; Bologna, M. Low-Intensity Transcranial Ultrasound Stimulation: Mechanisms of Action and Rationale for Future Applications in Movement Disorders. Brain Sci. 2022, 12, 611. https://doi.org/10.3390/brainsci12050611
Guerra A, Bologna M. Low-Intensity Transcranial Ultrasound Stimulation: Mechanisms of Action and Rationale for Future Applications in Movement Disorders. Brain Sciences. 2022; 12(5):611. https://doi.org/10.3390/brainsci12050611
Chicago/Turabian StyleGuerra, Andrea, and Matteo Bologna. 2022. "Low-Intensity Transcranial Ultrasound Stimulation: Mechanisms of Action and Rationale for Future Applications in Movement Disorders" Brain Sciences 12, no. 5: 611. https://doi.org/10.3390/brainsci12050611