
The Human Myelin Proteome and Sub-Metalloproteome Interaction Map: Relevance to Myelin-Related Neurological Diseases

 ,
,  and
and 
Abstract
:
1. Introduction
2. Materials and Methods
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Trapp, B.D.; Kidd, G.J. Structure of the Myelinated Axon. In Myelin Biology and Disorders; Academic Press: Cambridge, MA, USA, 2004; pp. 3–27. [Google Scholar]
- Apostolopoulos, V.; Rostami, A.; Matsoukas, J. The Long Road of Immunotherapeutics against Multiple Sclerosis. Brain Sci. 2020, 10, 288. [Google Scholar] [CrossRef] [PubMed]
- Tzakos, A.; Kursula, P.; Troganis, A.; Theodorou, V.; Tselios, T.; Svarnas, C.; Matsoukas, J.; Apostolopoulos, V.; Gerothanassis, I. Structure and Function of the Myelin Proteins: Current Status and Perspectives in Relation to Multiple Sclerosis. Curr. Med. Chem. 2005, 12, 1569–1587. [Google Scholar] [CrossRef] [PubMed]
- Romanelli, E.; Merkler, D.; Mezydlo, A.; Weil, M.T.; Weber, M.S.; Nikic, I.; Potz, S.; Meinl, E.; Matznick, F.E.; Kreutzfeldt, M.; et al. Myelinosome formation represents an early stage of oligodendrocyte damage in multiple sclerosis and its animal model. Nat. Commun. 2016, 7, 13275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, K.L.; Stewart, D.G.; Friedman, J.I.; Buchsbaum, M.; Harvey, P.D.; Hof, P.R.; Buxbaum, J.; Haroutunian, V. White Matter Changes in Schizophrenia. Arch. Gen. Psychiatry 2003, 60, 443. [Google Scholar] [CrossRef] [PubMed]
- Peters, A. The effects of normal aging on myelin and nerve fibers: A review. J. Neurocytol. 2002, 31, 581–593. [Google Scholar] [CrossRef] [PubMed]
- Poitelon, Y.; Kopec, A.M.; Belin, S. Myelin Fat Facts: An Overview of Lipids and Fatty Acid Metabolism. Cells 2020, 9, 812. [Google Scholar] [CrossRef] [Green Version]
- Werner, H.B.; Krämer-Albers, E.-M.; Strenzke, N.; Saher, G.; Tenzer, S.; Ohno-Iwashita, Y.; De Monasterio-Schrader, P.; Möbius, W.; Moser, T.; Griffiths, I.R.; et al. A critical role for the cholesterol-associated proteolipids PLP and M6B in myelination of the central nervous system. Glia 2013, 61, 567–586. [Google Scholar] [CrossRef]
- Raasakka, A.; Ruskamo, S.; Kowal, J.; Barker, R.; Baumann, A.; Martel, A.; Tuusa, J.; Myllykoski, M.; Bürck, J.; Ulrich, A.S.; et al. Membrane Association Landscape of Myelin Basic Protein Portrays Formation of the Myelin Major Dense Line. Sci. Rep. 2017, 7, 4974. [Google Scholar] [CrossRef]
- Nawaz, S.; Schweitzer, J.; Jahn, O.; Werner, H.B. Molecular evolution of myelin basic protein, an abundant structural myelin component. Glia 2013, 61, 1364–1377. [Google Scholar] [CrossRef]
- Quarles, H.R. Myelin-associated glycoprotein (MAG): Past, present and beyond. J. Neurochem. 2007, 100, 1431–1448. [Google Scholar] [CrossRef]
- Pham-Dinh, D.; Della Gaspera, B.; Kerlero de Rosbo, N.; Dautigny, A. Structure of the human myelin/oligodendrocyte glyco-protein gene and multiple alternative spliced isoforms. Genomics 1995, 29, 345–352. [Google Scholar] [CrossRef] [PubMed]
- Devaux, J.; Gow, A. Tight junctions potentiate the insulative properties of small CNS myelinated axons. J. Cell Biol. 2008, 183, 909–921. [Google Scholar] [CrossRef] [PubMed]
- Jahn, O.; Siems, S.B.; Kusch, K.; Hesse, D.; Jung, R.B.; Liepold, T.; Uecker, M.; Sun, T.; Werner, H.B. The CNS Myelin Proteome: Deep Profile and Persistence After Post-mortem Delay. Front. Cell. Neurosci. 2020, 14, 239. [Google Scholar] [CrossRef] [PubMed]
- Siems, S.B.; Jahn, O.; Eichel, M.A.; Kannaiyan, N.; Wu, L.M.N.; Sherman, D.L.; Kusch, K.; Hesse, D.; Jung, R.B.; Fledrich, R.; et al. Proteome profile of peripheral myelin in healthy mice and in a neuropathy model. eLife 2020, 9, e51406. [Google Scholar] [CrossRef] [PubMed]
- Ishii, A.; Dutta, R.; Wark, G.M.; Hwang, S.I.; Han, D.K.; Trapp, B.D.; Pfeiffer, S.E.; Bansal, R. Human myelin proteome and comparative analysis with mouse myelin. Proc. Natl. Acad. Sci. USA 2009, 106, 14605–14610. [Google Scholar] [CrossRef] [Green Version]
- Orchard, S.; Ammari, M.; Aranda, B.; Breuza, L.; Briganti, L.; Broackes-Carter, F.; Campbell, N.H.; Chavali, G.; Chen, C.; del-Toro, N.; et al. The MIntAct project—IntAct as a common curation platform for 11 molecular interaction databases. Nucleic Acids Res. 2014, 42, D358–D363. [Google Scholar] [CrossRef] [Green Version]
- Chasapis, C.T. Interactions between metal binding viral proteins and human targets as revealed by network-based bioinformatics. J. Inorg. Biochem. 2018, 186, 157–161. [Google Scholar] [CrossRef]
- Chasapis, C.T.; Andreini, C.; Georgiopolou, A.K.; Stefanidou, M.E.; Vlamis-Gardikas, A. Identification of the zinc, copper and cadmium metalloproteome of the protozoon Tetrahymena thermophila by systematic bioinformatics. Arch. Microbiol. 2017, 199, 1141–1149. [Google Scholar] [CrossRef]
- Peana, M.; Chasapis, C.T.; Simula, G.; Medici, S.; Zoroddu, M.A. A Model for Manganese interaction with Deinococcus radiodurans proteome network involved in ROS response and defense. J. Trace Elem. Med. Biol. 2018, 50, 465–473. [Google Scholar] [CrossRef]
- Chasapis, C.T. Shared gene-network signatures between the human heavy metal proteome and neurological disorders and cancer types. Metallomics 2018, 10, 1678–1686. [Google Scholar] [CrossRef]
- Andreini, C.; Bertini, I.; Cavallaro, G.; Decaria, L.; Rosato, A. A Simple Protocol for the Comparative Analysis of the Structure and Occurrence of Biochemical Pathways Across Superkingdoms. J. Chem. Inf. Modeling 2011, 51, 730–738. [Google Scholar] [CrossRef] [PubMed]
- Chasapis, C.T.; Georgiopoulou, A.K.; Perlepes, S.P.; Bjørklund, G.; Peana, M. A SARS-CoV-2 –human metalloproteome interaction map. J. Inorg. Biochem. 2021, 219, 111423. [Google Scholar] [CrossRef] [PubMed]
- Stefanidou, M.C.; Loutsidou, A.T.; Chasapis, C.A.; Spiliopoulou, C. Immunotoxicity of Cocaine and Crack. Curr. Drug Abus. Rev. 2011, 4, 95–97. [Google Scholar] [CrossRef] [PubMed]
- Chasapis, C.T. Building Bridges Between Structural and Network-Based Systems Biology. Mol. Biotechnol. 2019, 61, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Putignano, V.; Rosato, A.; Banci, L.; Andreini, C. MetalPDB in 2018: A database of metal sites in biological macromolecular structures. Nucleic Acids Res. 2018, 46, D459–D464. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, T.J.; Eddy, S.R. nhmmer: DNA homology search with profile HMMs. Bioinformatics 2013, 29, 2487–2489. [Google Scholar] [CrossRef] [Green Version]
- Li, T.; Wernersson, R.; Hansen, R.B.; Horn, H.; Mercer, J.; Slodkowicz, G.; Workman, C.T.; Rigina, O.; Rapacki, K.; Stærfeldt, H.H.; et al. A scored human protein–protein interaction network to catalyze genomic interpretation. Nat. Methods 2016, 14, 61–64. [Google Scholar] [CrossRef] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A Software Environment for Integrated Models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Jahn, O.; Tenzer, S.; Werner, H.B. Myelin Proteomics: Molecular Anatomy of an Insulating Sheath. Mol. Neurobiol. 2009, 40, 55–72. [Google Scholar] [CrossRef] [Green Version]
- Barabasi, A.L.; Albert, R. Emergence of scaling in random networks. Science 1999, 286, 509–512. [Google Scholar] [CrossRef] [Green Version]
- Foote, M.; Zhou, Y. 14-3-3 proteins in neurological disorders. Int. J. Biochem. Mol. Biol. 2012, 3, 152–164. [Google Scholar] [PubMed]
- Angrand, P.-O.; Segura, I.; Völkel, P.; Ghidelli, S.; Terry, R.; Brajenovic, M.; Vintersten, K.; Klein, R.; Superti-Furga, G.; Drewes, G.; et al. Transgenic Mouse Proteomics Identifies New 14-3-3-associated Proteins Involved in Cytoskeletal Rearrangements and Cell Signaling. Mol. Cell. Proteom. 2006, 5, 2211–2227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buchner, J.; Li, J. Structure, Function and Regulation of the Hsp90 Machinery. Biomed. J. 2013, 36, 106. [Google Scholar] [CrossRef] [PubMed]
- Streicher, J.M. The role of heat shock protein 90 in regulating pain, opioid signaling, and opioid antinociception. Vitam. Horm. 2019, 111, 91–103. [Google Scholar] [PubMed]
- Cao, W.; Harnett, K.M.; Behar, J.; Biancani, P. Group I Secreted PLA2 in the Maintenance of Human Lower Esophageal Sphincter Tone. Gastroenterology 2000, 119, 1243–1252. [Google Scholar] [CrossRef]
- Ferraz, C.R.; Arrahman, A.; Xie, C.; Casewell, N.R.; Lewis, R.J.; Kool, J.; Cardoso, F.C. Multifunctional toxins in snake venoms and therapeutic implications: From pain to hemorrhage and necrosis. Front. Ecol. Evol. 2019, 7, 218. [Google Scholar] [CrossRef] [Green Version]
- Misslin, C.; Velasco-Estevez, M.; Albert, M.; O’Sullivan, S.A.; Dev, K.K. Phospholipase A2 is involved in galactosylsphingosine-induced astrocyte toxicity, neuronal damage and demyelination. PLoS ONE 2017, 12, e0187217. [Google Scholar] [CrossRef] [Green Version]
- Trapp, B.D.; Kidd, G.J.; Pfeiffer, S.E.; Anitei, M. Cell Biology of Myelin Assembly. In Myelin Biology and Disorders; Academic Press: Cambridge, MA, USA, 2004; pp. 29–55. [Google Scholar]
- Huber, L.A.; Madison, D.L.; Simons, K.; Pfeiffer, S.E. Myelin membrane biogenesis by oligodendrocytes Developmental regulation of low molecular weight GTP-binding proteins. FEBS Lett. 1994, 347, 273–278. [Google Scholar]
- Anitei, M.; Cowan, A.E.; Pfeiffer, S.E.; Bansal, R. Role for Rab3a in oligodendrocyte morphological differentiation. J. Neurosci. Res. 2009, 87, 342–352. [Google Scholar] [CrossRef]
- Madison, D.L.; Krueger, W.H.; Cheng, D.; Trapp, B.D.; Pfeiffer, S.E. SNARE Complex Proteins, Including the Cognate Pair VAMP-2and Syntaxin-4, Are Expressed in Cultured Oligodendrocytes. J. Neurochem. 2008, 72, 988–998. [Google Scholar] [CrossRef]
- Ashley, R.H. Challenging accepted ion channel biology: p64 and the CLIC family of putative intracellular anion channel proteins (Review). Mol. Membr. Biol. 2009, 20, 1–11. [Google Scholar] [CrossRef]
- Berger, P.; Sirkowski, E.E.; Scherer, S.S.; Suter, U. Expression analysis of the N-Myc downstream-regulated gene 1 indicates that myelinating Schwann cells are the primary disease target in hereditary motor and sensory neuropathy-Lom. Neurobiol. Dis. 2004, 17, 290–299. [Google Scholar] [CrossRef] [PubMed]
- Blanco-Arias, P.; Einholm, A.P.; Mamsa, H.; Concheiro, C.; Gutierrez-de-Teran, H.; Romero, J.; Toustrup-Jensen, M.S.; Carracedo, A.; Jen, J.C.; Vilsen, B.; et al. A C-terminal mutation of ATP1A3 underscores the crucial role of sodium affinity in the pathophysiology of rapid-onset dystonia-parkinsonism. Hum. Mol. Genet. 2009, 18, 2370–2377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.; Chung, A.Y.; Na, J.E.; Lee, S.J.; Jeong, S.H.; Kim, E.; Sun, W.; Rhyu, I.J.; Park, H.C. Myelin degeneration induced by mutant superoxide dismutase 1 accumulation promotes amyotrophic lateral sclerosis. Glia 2019, 67, 1910–1921. [Google Scholar]
- Mani, K.; Cheng, F.; Havsmark, B.; Jönsson, M.; Belting, M.; Fransson, L.-Å. Prion, Amyloid β-derived Cu(II) Ions, or Free Zn(II) Ions Support S-Nitroso-dependent Autocleavage of Glypican-1 Heparan Sulfate. J. Biol. Chem. 2003, 278, 38956–38965. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Lee, J.E.; Heynen-Genel, S.; Suyama, E.; Ono, K.; Lee, K.; Ideker, T.; Aza-Blanc, P.; Gleeson, J.G. Functional genomic screen for modulators of ciliogenesis and cilium length. Nature 2010, 464, 1048–1051. [Google Scholar] [CrossRef] [Green Version]
- Gusic, M.; Schottmann, G.; Feichtinger, R.G.; Du, C.; Scholz, C.; Wagner, M.; Mayr, J.A.; Lee, C.-Y.; Yépez, V.A.; Lorenz, N.; et al. Bi-Allelic UQCRFS1 Variants Are Associated with Mitochondrial Complex III Deficiency, Cardiomyopathy, and Alopecia Totalis. Am. J. Hum. Genet. 2020, 106, 102–111. [Google Scholar] [CrossRef]
- Elkholi, R.; Abraham-Enachescu, I.; Trotta, A.P.; Rubio-Patiño, C.; Mohammed, J.N.; Luna-Vargas, M.P.A.; Gelles, J.D.; Kaminetsky, J.R.; Serasinghe, M.N.; Zou, C.; et al. MDM2 Integrates Cellular Respiration and Apoptotic Signaling through NDUFS1 and the Mitochondrial Network. Mol. Cell 2019, 74, 452–465.e7. [Google Scholar] [CrossRef] [Green Version]
- Rose, A.J.; Kiens, B.; Richter, E.A. Ca2+-calmodulin-dependent protein kinase expression and signalling in skeletal muscle during exercise. J. Physiol. 2006, 574, 889–903. [Google Scholar] [CrossRef]
- De Ligt, J.; Willemsen, M.H.; van Bon, B.W.M.; Kleefstra, T.; Yntema, H.G.; Kroes, T.; Vulto-van Silfhout, A.T.; Koolen, D.A.; de Vries, P.; Gilissen, C.; et al. Diagnostic Exome Sequencing in Persons with Severe Intellectual Disability. N. Engl. J. Med. 2012, 367, 1921–1929. [Google Scholar] [CrossRef] [Green Version]
- Scinicariello, F.; Murray, H.E.; Moffett, D.B.; Abadin, H.G.; Sexton, M.J.; Fowler, B.A. Lead and δ-Aminolevulinic Acid Dehydratase Polymorphism: Where Does It Lead? A Meta-Analysis. Environ. Health Perspect. 2007, 115, 35–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rama Rao, K.V.; Norenberg, M.D. Glutamine in the Pathogenesis of Hepatic Encephalopathy: The Trojan Horse Hypothesis Revisited. Neurochem. Res. 2013, 39, 593–598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weatherall, D.J.; Williams, T.N.; Allen, S.J.; O’Donnell, A. The Population Genetics and Dynamics of the Thalassemias. Hematol. Oncol. Clin. N. Am. 2010, 24, 1021–1031. [Google Scholar] [CrossRef] [PubMed]
- Freedman, M.S.; Bar-Or, A.; Oger, J.; Traboulsee, A.; Patry, D.; Young, C.; Olsson, T.; Li, D.; Hartung, H.P.; Krantz, M.; et al. MAESTRO-01, Investigators. A phase III study evaluating the efficacy and safety of MBP8298 in secondary progressive MS. Neurology 2011, 77, 1551–1560. [Google Scholar] [CrossRef] [PubMed]
- Mantzourani, E.; Mavromoustakos, T.; Platts, J.; Matsoukas, J.; Tselios, T. Structural Requirements for Binding of Myelin Basic Protein (MBP) Peptides to MHC II: Effects on Immune Regulation. Curr. Med. Chem. 2005, 12, 1521–1535. [Google Scholar] [CrossRef]
- Katsara, M.; Matsoukas, J.; Deraos, G.; Apostolopoulos, V. Towards immunotherapeutic drugs and vaccines against multiple sclerosis. Acta Biochim. Biophys. Sin. 2008, 40, 636–642. [Google Scholar] [CrossRef] [Green Version]
- Friligou, I.; Papadimitriou, E.; Gatos, D.; Matsoukas, J.; Tselios, T. Microwave-assisted solid-phase peptide synthesis of the 60–110 domain of human pleiotrophin on 2-chlorotrityl resin. Amino Acids 2010, 40, 1431–1440. [Google Scholar] [CrossRef]
- Dagkonaki, A.; Avloniti, M.; Evangelidou, M.; Papazian, I.; Kanistras, I.; Tseveleki, V.; Lampros, F.; Tselios, T.; Jensen, L.T.; Möbius, W.; et al. Mannan-MOG35-55 Reverses Experimental Autoimmune Encephalomyelitis, inducing a Peripheral Type 2 Myeloid Response, Reducing CNS Inflammation, and Preserving Axons in Spinal Cord Lesions. Front. Immunol. 2020, 11, 575451. [Google Scholar] [CrossRef]
- Matsoukas, J.; Deraos, G.; Kelaidonis, K.; Hossain, M.K.; Feehan, J.; Tzakos, A.G.; Matsoukas, E.; Topoglidis, E.; Apostolopoulos, V. Myelin Peptide–Mannan Conjugate Multiple Sclerosis Vaccines: Conjugation Efficacy and Stability of Vaccine Ingredient. Vaccines 2021, 9, 1456. [Google Scholar] [CrossRef]
- Robinson, W.H.; Steinman, L. Epstein-Barr virus and multiple sclerosis. Science 2022, 375, 264–265. [Google Scholar] [CrossRef]
- Bjornevik, K.; Cortese, M.; Healy, B.C.; Kuhle, J.; Mina, M.J.; Leng, Y.; Elledge, S.J.; Niebuhr, D.W.; Scher, A.I.; Munger, K.L.; et al. Longitudinal analysis reveals high prevalence of Epstein-Barr virus associated with multiple sclerosis. Science 2022, 375, 296–301. [Google Scholar] [CrossRef] [PubMed]
- Bar-Or, A.; Pender, M.P.; Khanna, R.; Steinman, L.; Hartung, H.-P.; Maniar, T.; Croze, E.; Aftab, B.T.; Giovannoni, G.; Joshi, M.A. Epstein–Barr Virus in Multiple Sclerosis: Theory and Emerging Immunotherapies. Trends Mol. Med. 2020, 26, 296–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
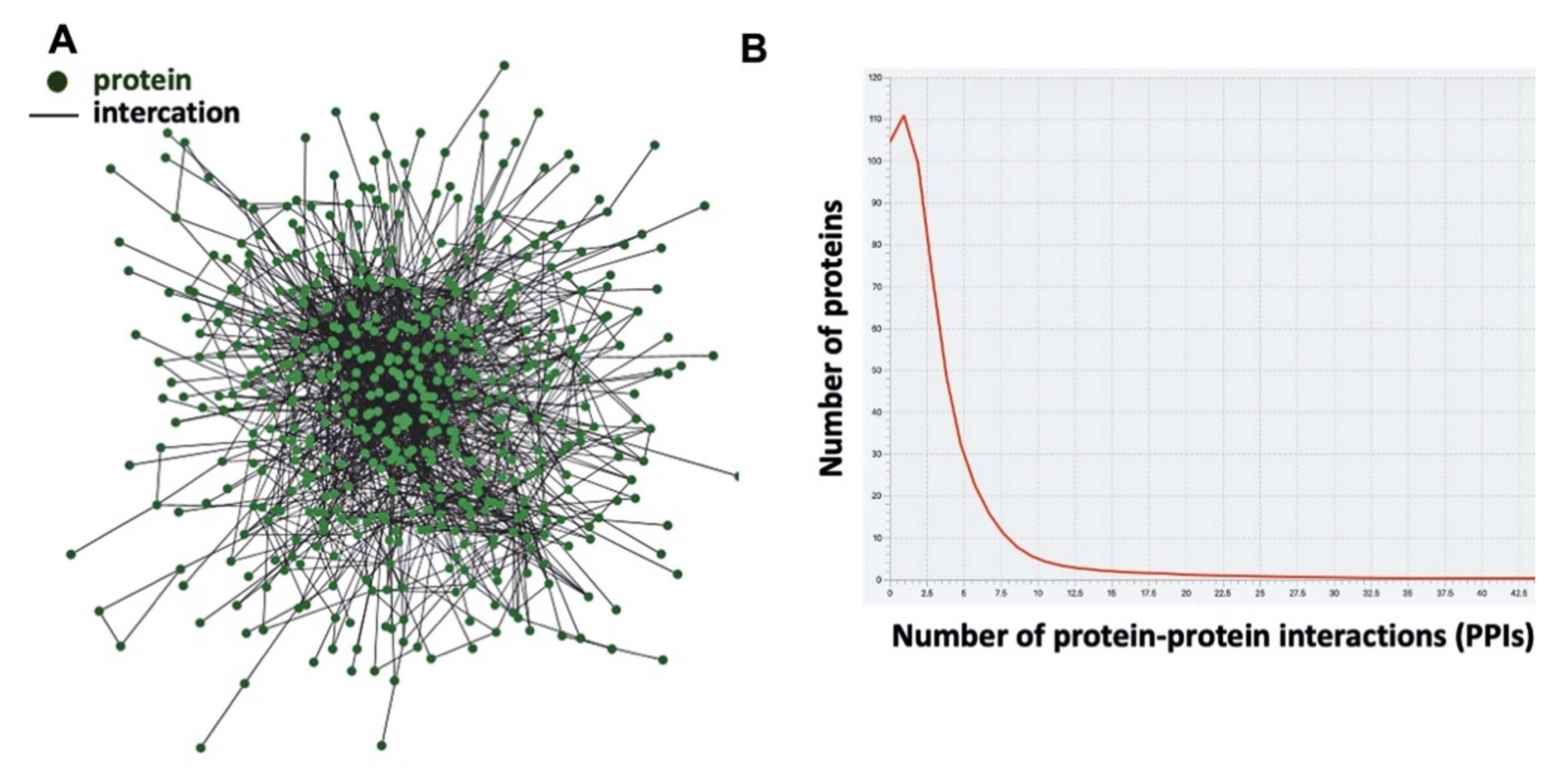
| Summary Statistics | |
|---|---|
| Number of proteins | 567 |
| Number of interactions | 2017 |
| Avarage number of interactions | 6.2 |
| Network diameter | 11 |
| Number of homodimers (self-loops) | 268 |
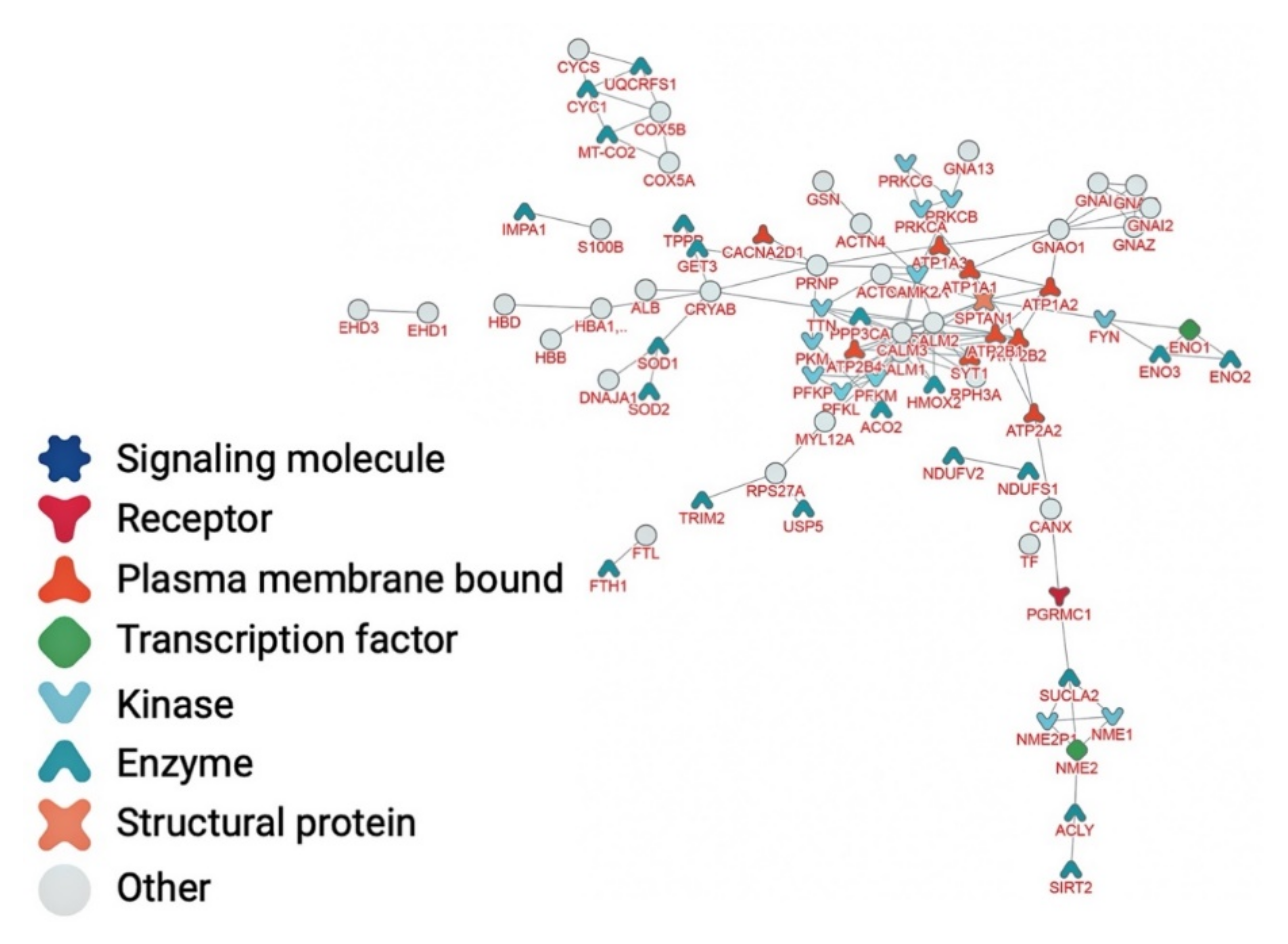
| Entry | Gene Name | Protein Name | Degree |
|---|---|---|---|
| P63104 | YWHAZ | 14-3-3 protein zeta/delta | 46 |
| P27348 | YWHAQ | 14-3-3 protein theta | 43 |
| P07900 | HSP90AA1 | Heat shock protein HSP 90-alpha | 41 |
| P0CG48 | UBC | Polyubiquitin-C | 35 |
| P02545 | LMNA | Prelamin-A/C | 32 |
| P04792 | HSPB1 | Heat shock protein beta-1 (HspB1) | 29 |
| P0DP24 | CALM2 | Calmodulin-2 | 29 |
| P17252 | PRKCA | Protein kinase C alpha type (PKC-A) | 28 |
| P0DP23 | CALM1 | Calmodulin-1 | 26 |
| P20648 | ATP4A | Potassium-transporting ATPase alpha chain 1 | 24 |
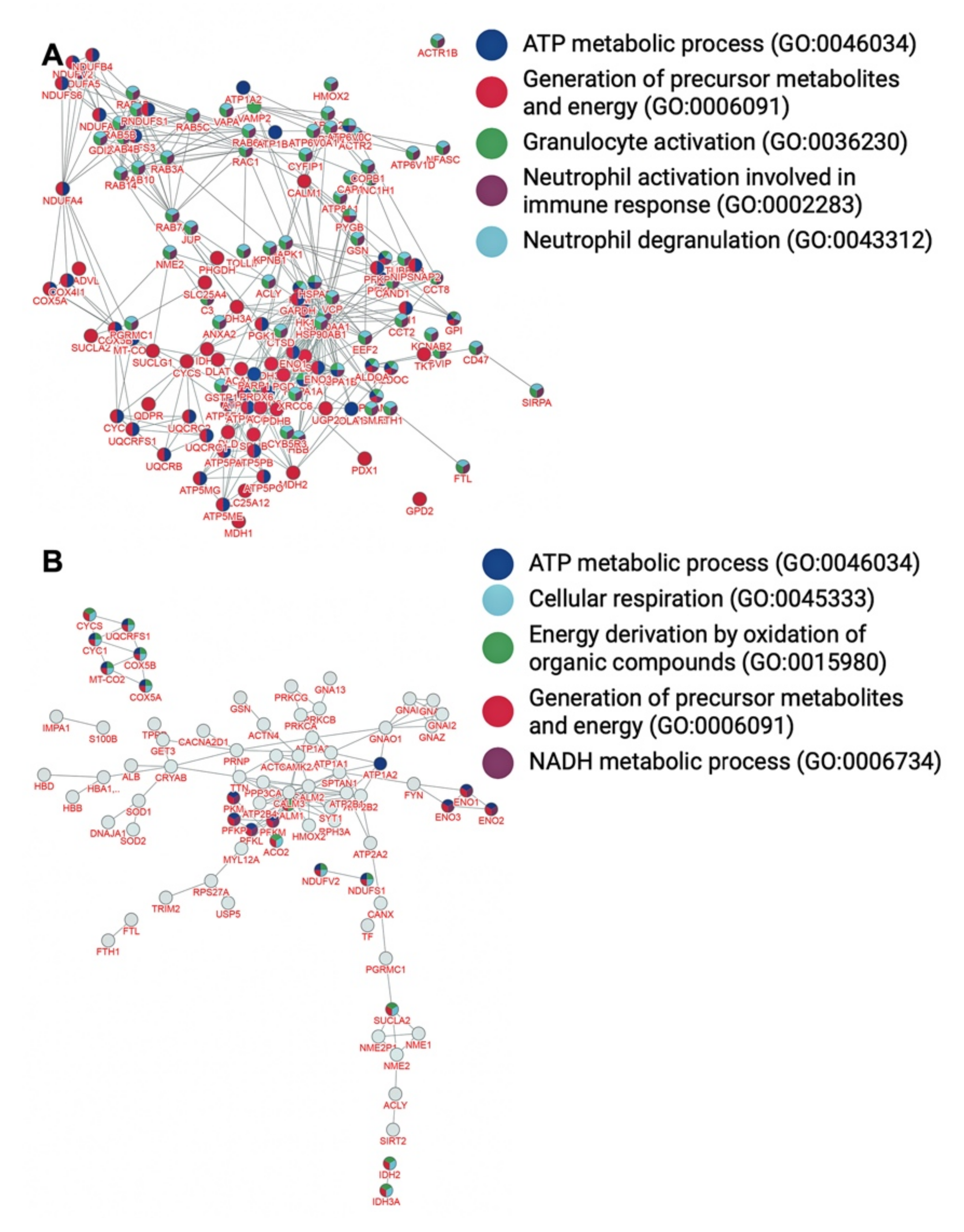
| Biological Processes | Overlap | p-Value |
|---|---|---|
| Generation of precursor metabolites and energy (GO:0006091) | 73/567 | 4.2 × 10−92 |
| Granulocyte activation (GO:0036230) | 80/567 | 3.9 × 10−89 |
| Neutrophil degranulation (GO:0043312) | 79/567 | 3.6 × 10−88 |
| Neutrophil activation involved in immune response (GO:0002283) | 79/567 | 4.3 × 10−88 |
| TP metabolic process (GO:0046034) | 49/567 | 2.9 × 10−67 |
| Pathways | Overlap | p-Value |
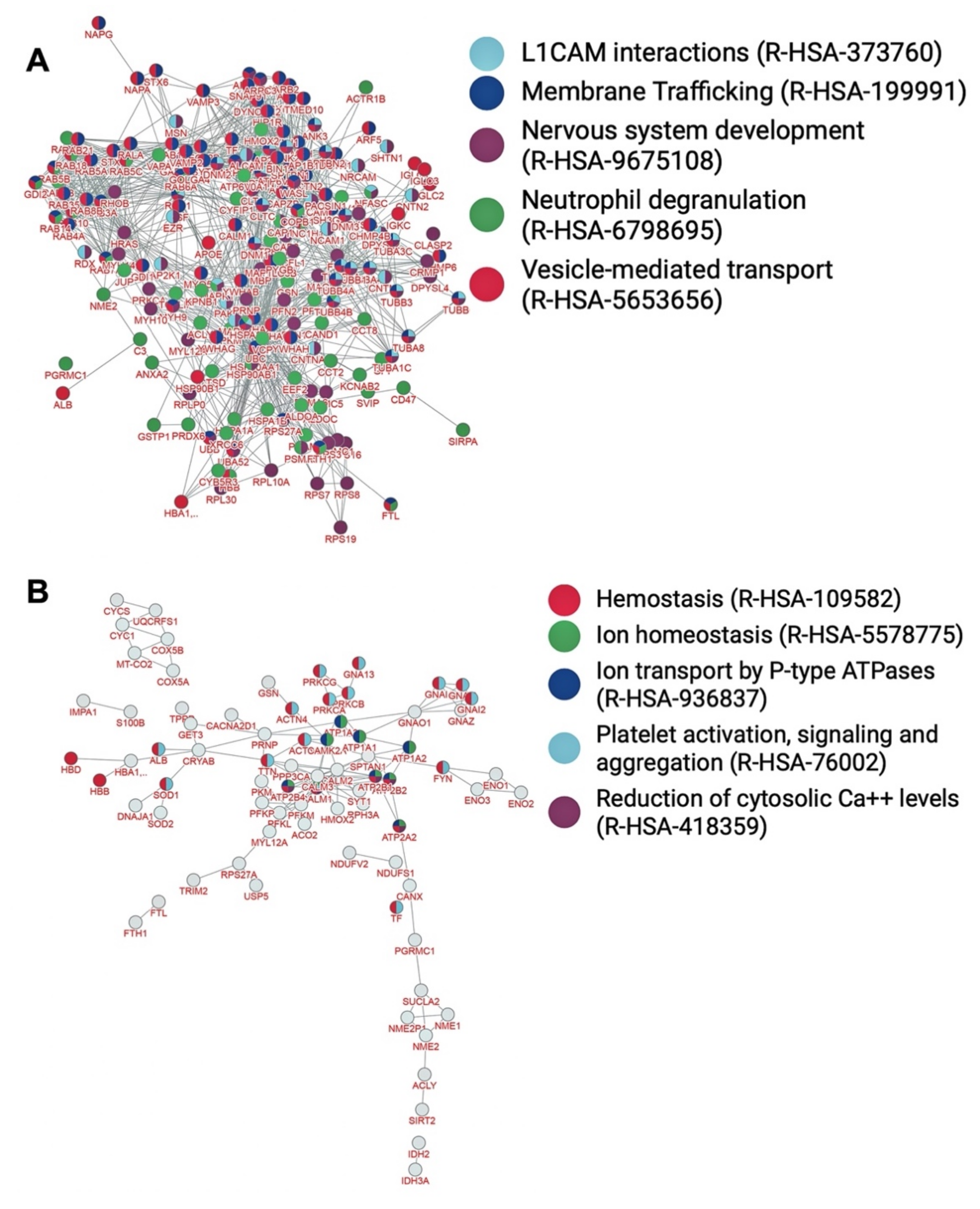
| Vesicle-mediated transport (R-HSA-5653656) | 105/567 | 2.6 × 10−95 |
| Membrane trafficking (R-HSA-199991) | 95/567 | 4.8 × 10−87 |
| Nervous system development (R-HSA-9675108) | 85/567 | 6.6 × 10−76 |
| Neutrophil degranulation (R-HSA-6798695) | 79/567 | 4.8 × 10−74 |
| L1CAM interactions (R-HSA-373760) | 42/567 | 9.3 × 10−54 |
| Diseases | Overlap | p-Value |
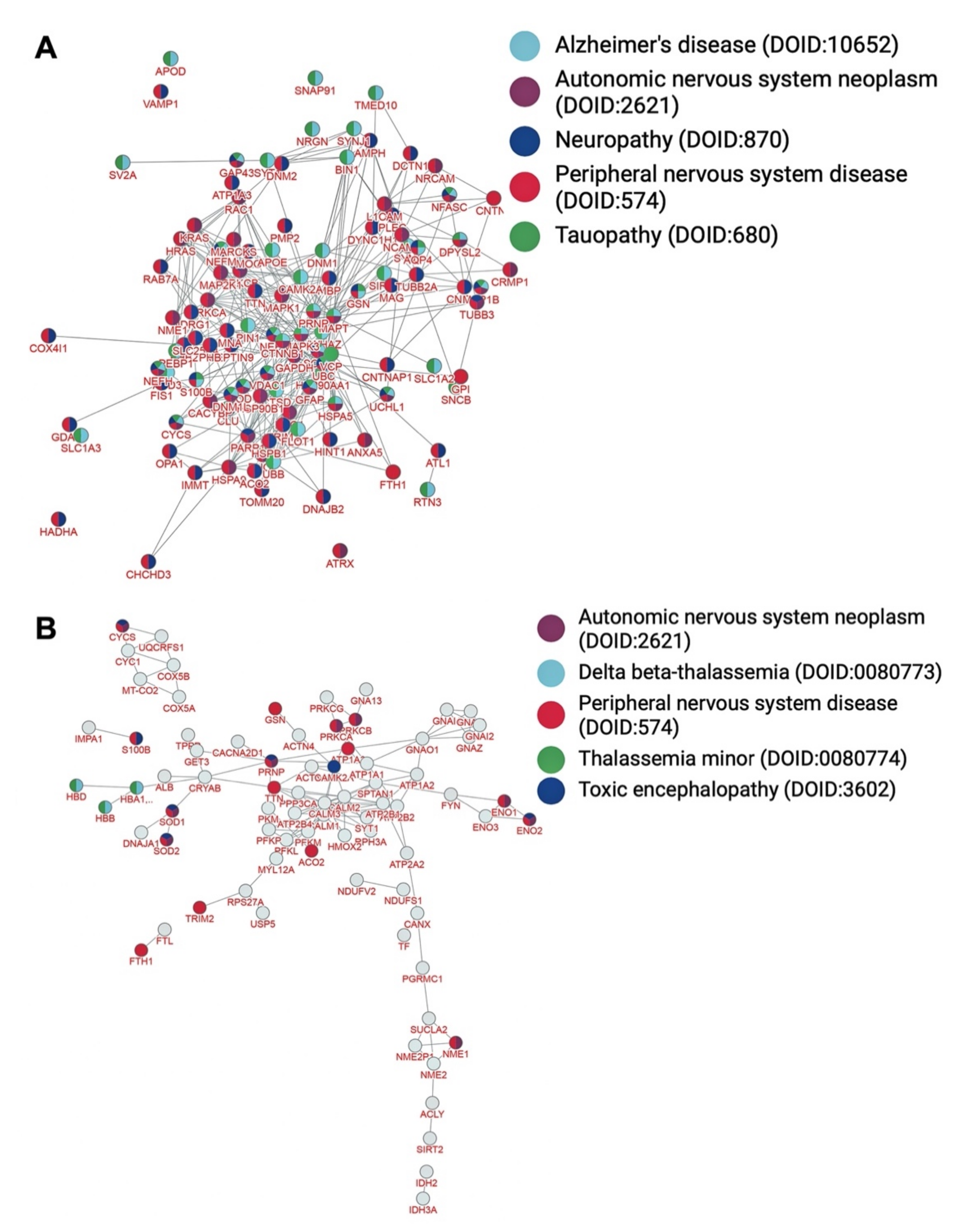
| Peripheral nervous system disease (DOID:574) | 91/567 | <9.9 × 10−99 |
| Neuropathy (DOID:870) | 59/567 | 3.9 × 10−69 |
| Tauopathy (DOID:680) | 54/567 | 2.9 × 10−62 |
| Alzheimer’s disease (DOID:10652) | 53/567 | 9.4 × 10−61 |
| Autonomic nervous system neoplasm (DOID:2621) | 47/567 | 1.7 × 10−54 |
| Biological Processes | Overlap | p-Value |
|---|---|---|
| Generation of precursor metabolites and energy (GO:0006091) | 25/119 | 4.2 × 10−21 |
| ATP metabolic process (GO:0046034) | 17/119 | 7.7 × 10−17 |
| Energy derivation by oxidation of organic compounds (GO:0015980) | 17/119 | 2.2 × 10−16 |
| Cellular respiration (GO:0045333) | 15/119 | 6.1 × 10−16 |
| NADH metabolic process (GO:0006734) | 8/119 | 3.5 × 10−12 |
| Pathways | Overlap | p-Value |
| Hemostasis (R-HSA-109582) | 30/119 | 2.4 × 10−18 |
| Ion transport by P-type ATPases (R-HSA-936837) | 12/119 | 3.9 × 10−16 |
| Ion homeostasis (R-HSA-5578775) | 11/119 | 1.6 × 10−14 |
| Platelet activation, signaling, and aggregation (R-HSA-76002) | 18/119 | 1.8 × 10−14 |
| Reduction of cytosolic Ca++ levels (R-HSA-418359) | 7/119 | 1.7 × 10−13 |
| Diseases | Overlap | p-Value |
| Peripheral nervous system disease (DOID:574) | 20/119 | 8.9 × 10−9 |
| Toxic encephalopathy (DOID:3602) | 10/119 | 1.1 × 10−7 |
| Delta beta-thalassemia (DOID:0080773) | 3/119 | 2.1 × 10−7 |
| Thalassemia minor (DOID:0080774) | 3/119 | 2.1 × 10−7 |
| Autonomic nervous system neoplasm (DOID:2621) | 12/119 | 3.2 × 10−7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chasapis, C.T.; Kelaidonis, K.; Ridgway, H.; Apostolopoulos, V.; Matsoukas, J.M. The Human Myelin Proteome and Sub-Metalloproteome Interaction Map: Relevance to Myelin-Related Neurological Diseases. Brain Sci. 2022, 12, 434. https://doi.org/10.3390/brainsci12040434
Chasapis CT, Kelaidonis K, Ridgway H, Apostolopoulos V, Matsoukas JM. The Human Myelin Proteome and Sub-Metalloproteome Interaction Map: Relevance to Myelin-Related Neurological Diseases. Brain Sciences. 2022; 12(4):434. https://doi.org/10.3390/brainsci12040434
Chicago/Turabian StyleChasapis, Christos T., Konstantinos Kelaidonis, Harry Ridgway, Vasso Apostolopoulos, and John M. Matsoukas. 2022. "The Human Myelin Proteome and Sub-Metalloproteome Interaction Map: Relevance to Myelin-Related Neurological Diseases" Brain Sciences 12, no. 4: 434. https://doi.org/10.3390/brainsci12040434