
Novel Approaches in the Immunotherapy of Multiple Sclerosis: Cyclization of Myelin Epitope Peptides and Conjugation with Mannan

 ,
,  ,
,  and
and
Abstract
: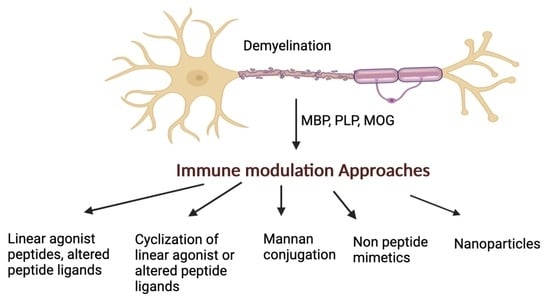
1. Introduction
- (a)
- Use linear MBP, PLP, and MOG epitopes with or without amino acid mutations to gain insights into the molecular basis of the disease. This information is aided by nuclear magnetic resonance (NMR) studies and structural information of the epitope ligand bound to the MHC [19];
- (b)
- (c)
- (d)
- Synthesis of non-peptide mimetic analogs to decrease their flexibility and increase their therapeutic index;
- (e)
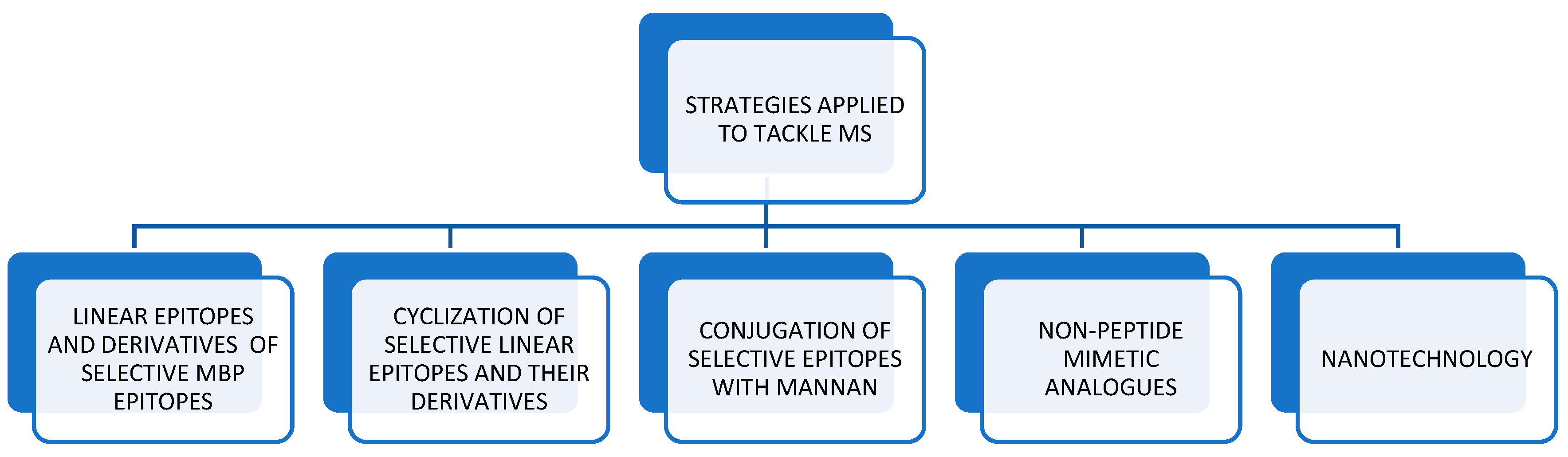
2. Applied Strategies Utilized against MS
2.1. Linear Epitopes and Derivatives of Selective MBP Epitopes
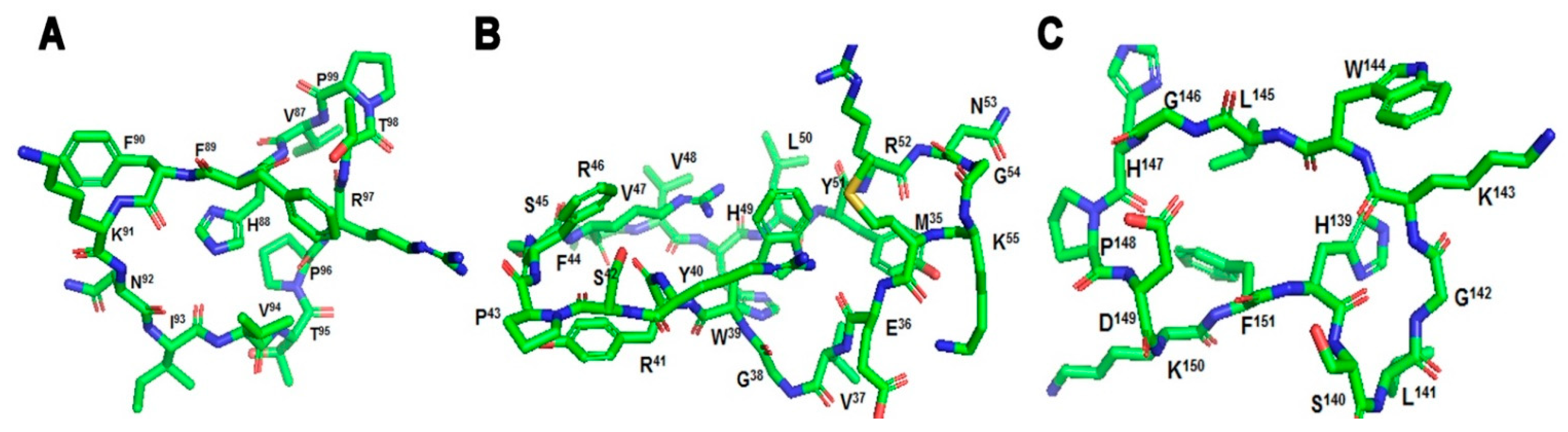
2.2. Cyclization of Selective Linear Epitopes and Their Derivatives
2.3. Conjugation of Selective Epitopes with Mannan
2.4. Citrullination of MBP Derived Peptides Role in Disease
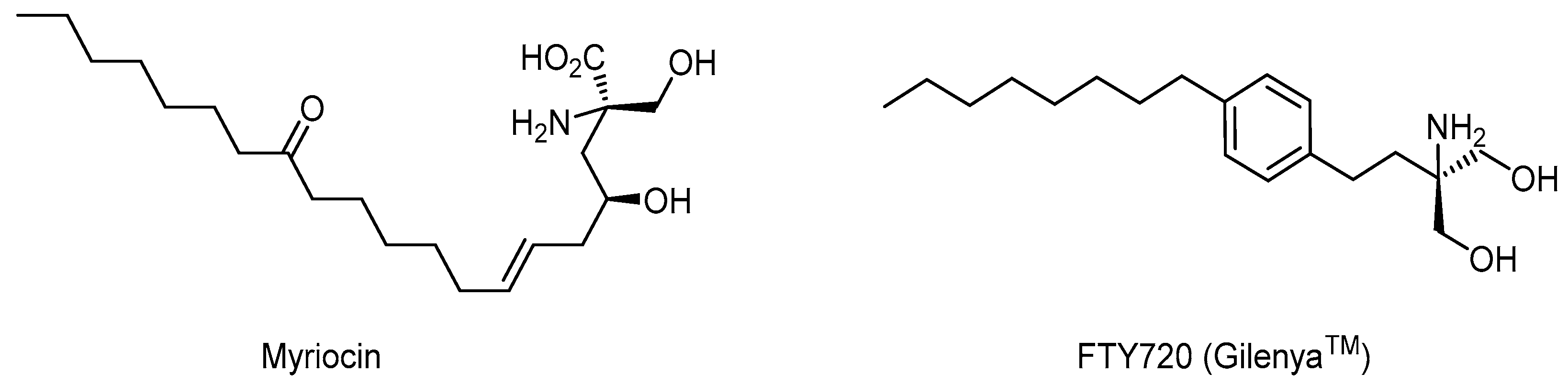
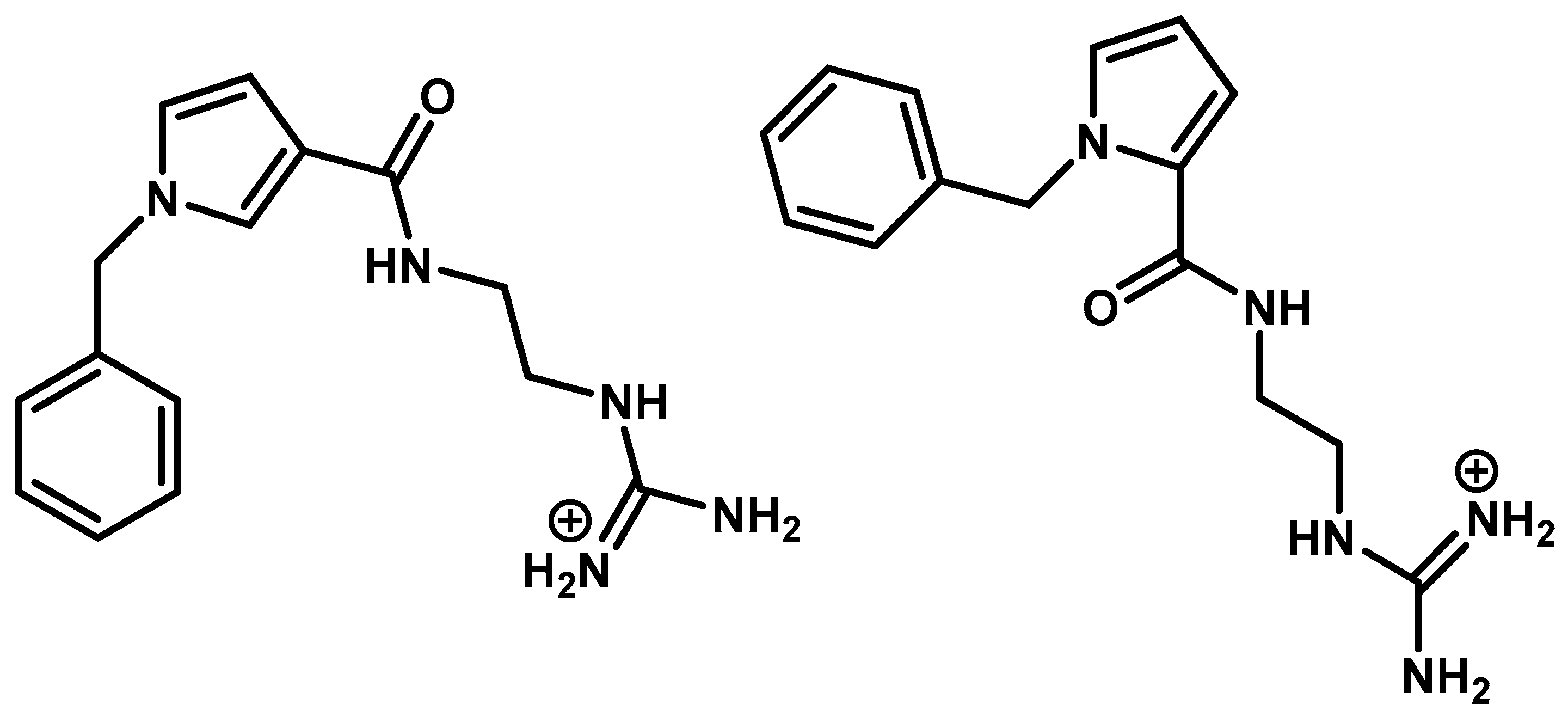
2.5. Non-Peptide Mimetic Analogs
2.6. Nanotechnology Approaches
3. Future Aspects
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Compston, A.; Coles, A. Multiple sclerosis. Lancet 2002, 359, 1221–1231. [Google Scholar] [CrossRef]
- Steinman, M.D.L. Multiple Sclerosis: A Coordinated Immunological Attack against Myelin in the Central Nervous System. Cell 1996, 85, 299–302. [Google Scholar] [CrossRef] [Green Version]
- Grytten, N.; Torkildsen, Ø.; Myhr, K.M. Time trends in the incidence and prevalence of multiple sclerosis in Norway during eight decades. Acta Neurol. Scand. 2015, 132, 29–36. [Google Scholar] [CrossRef] [Green Version]
- Polman, C.H.; Reingold, S.C.; Banwell, B.; Clanet, M.; Cohen, J.A.; Filippi, M.; Fujihara, K.; Havrdova, E.; Hutchinson, M.; Kappos, L.; et al. Diagnostic criteria for multiple sclerosis: 2010 Revisions to the McDonald criteria. Ann. Neurol. 2011, 69, 292–302. [Google Scholar] [CrossRef] [Green Version]
- Eckstein, C.; Bhatti, M.T. Currently approved and emerging oral therapies in multiple sclerosis: An update for the ophthalmologist. Surv. Ophthalmol. 2016, 61, 318–332. [Google Scholar] [CrossRef]
- Dargahi, N.; Katsara, M.; Tselios, T.; Androutsou, M.E.; de Courten, M.; Matsoukas, J.; Apostolopoulos, V. Multiple Sclerosis: Immunopathology and Treatment Update. Brain Sci. 2017, 7, 78. [Google Scholar] [CrossRef] [Green Version]
- Florou, D.; Katsara, M.; Feehan, J.; Dardiotis, E.; Apostolopoulos, V. Anti-CD20 Agents for Multiple Sclerosis: Spotlight on Ocrelizumab and Ofatumumab. Brain Sci. 2020, 10, 758. [Google Scholar] [CrossRef]
- Kammona, O.; Kiparissides, C. Recent Advances in Antigen-Specific Immunotherapies for the Treatment of Multiple Sclerosis. Brain Sci. 2020, 10, 333. [Google Scholar] [CrossRef]
- Metaxakis, A.; Petratou, D.; Tavernarakis, N. Molecular Interventions towards Multiple Sclerosis Treatment. Brain Sci. 2020, 10, 299. [Google Scholar] [CrossRef]
- Dai, H.; Ciric, B.; Zhang, G.-X.; Rostami, A. Interleukin-10 plays a crucial role in suppression of experimental autoimmune encephalomyelitis by Bowman–Birk inhibitor. J. Neuroimmunol. 2012, 245, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rostami, A.; Gregorian, S.K. Peptide 53–78 of myelin P2 protein is a T cell epitope for the induction of experimental autoimmune neuritis. Cell. Immunol. 1991, 132, 433–441. [Google Scholar] [CrossRef]
- Rostami, A.; Gregorian, S.K.; Brown, M.J.; Pleasure, D.E. Induction of severe experimental autoimmune neuritis with a synthetic peptide corresponding to the 53–78 amino acid sequence of the myelin P2 protein. J. Neuroimmunol. 1990, 30, 145–151. [Google Scholar] [CrossRef]
- Martin, R.; McFarland, H.F.; McFarlin, D.E. Immunological Aspects of Demyelinating Diseases. Annu. Rev. Immunol. 1992, 10, 153–187. [Google Scholar] [CrossRef] [PubMed]
- Hafler, D.A.; Weiner, H.L. Immunologic Mechanisms and Therapy in Multiple Sclerosis. Immunol. Rev. 1995, 144, 75–107. [Google Scholar] [CrossRef]
- Kappos, L.; Comi, G.; Panitch, H.; Oger, J.; Antel, J.; Conlon, P.; Steinman, L.; Comi, G.; Kappos, L.; Oger, J.; et al. Induction of a non-encephalitogenic type 2 T helper-cell autoimmune response in multiple sclerosis after administration of an altered peptide ligand in a placebo-controlled, randomized phase II trial. Nat. Med. 2000, 6, 1176–1182. [Google Scholar] [CrossRef]
- Bielekova, B.; Goodwin, B.; Richert, N.; Cortese, I.; Kondo, T.; Afshar, G.; Gran, B.; Eaton, J.; Antel, J.; Frank, J.A.; et al. Encephalitogenic potential of the myelin basic protein peptide (amino acids 83–99) in multiple sclerosis: Results of a phase II clinical trial with an altered peptide ligand. Nat. Med. 2000, 6, 1167–1175. [Google Scholar] [CrossRef]
- Anagnostouli, M.; Artemiadis, A.; Gontika, M.; Skarlis, C.; Markoglou, N.; Katsavos, S.; Kilindireas, K.; Doxiadis, I.; Stefanis, L. HLA-DPB1*03 as Risk Allele and HLA-DPB1*04 as Protective Allele for Both Early- and Adult-Onset Multiple Sclerosis in a Hellenic Cohort. Brain Sci. 2020, 10, 374. [Google Scholar] [CrossRef]
- Vergelli, M.; Hemmer, B.; Utz, U.; Vogt, A.; Kalbus, M.; Tranquill, L.; Conlon, P.; Ling, N.; Steinman, L.; McFarland, H.F.; et al. Differential activation of human autoreactive T cell clones by altered peptide ligands derived from myelin basic protein peptide (87–99). Eur. J. Immunol. 1996, 26, 2624–2634. [Google Scholar] [CrossRef]
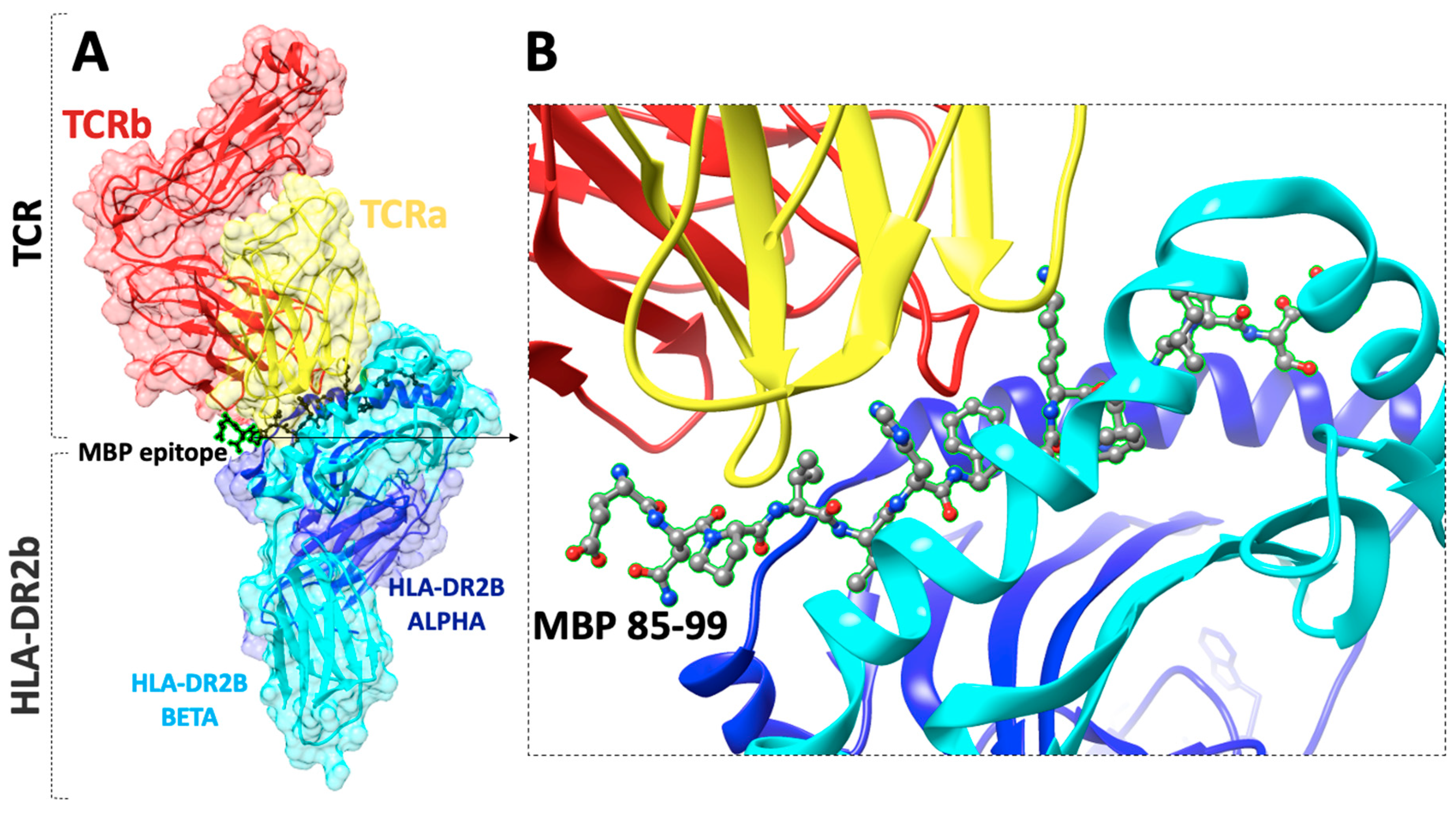
- Mantzourani, E.; Mavromoustakos, T.; Platts, J.; Matsoukas, J.; Tselios, T. Structural Requirements for Binding of Myelin Basic Protein (MBP) Peptides to MHC II: Effects on Immune Regulation. Curr. Med. Chem. 2005, 12, 1521–1535. [Google Scholar] [CrossRef]
- Katsara, M.; Deraos, G.; Tselios, T.; Matsoukas, M.-T.; Friligou, I.; Matsoukas, J.; Apostolopoulos, V. Design and Synthesis of a Cyclic Double Mutant Peptide (cyclo(87−99)[A91,A96]MBP87−99) Induces Altered Responses in Mice after Conjugation to Mannan: Implications in the Immunotherapy of Multiple Sclerosis. J. Med. Chem. 2008, 52, 214–218. [Google Scholar] [CrossRef]
- Katsara, M.; Yuriev, E.; Ramsland, P.A.; Deraos, G.; Tselios, T.; Matsoukas, J.; Apostolopoulos, V. A double mutation of MBP83–99 peptide induces IL-4 responses and antagonizes IFN-γ responses. J. Neuroimmunol. 2008, 200, 77–89. [Google Scholar] [CrossRef] [PubMed]
- Tselios, T.; Probert, L.; Daliani, I.; Matsoukas, E.; Troganis, A.; Gerothanassis, I.P.; Mavromoustakos, T.; Moore, G.J.; Matsoukas, J.M. Design and Synthesis of a Potent Cyclic Analogue of the Myelin Basic Protein Epitope MBP72-85: Importance of the Ala81 Carboxyl Group and of a Cyclic Conformation for Induction of Experimental Allergic Encephalomyelitis. J. Med. Chem. 1999, 42, 1170–1177. [Google Scholar] [CrossRef] [PubMed]
- Matsoukas, J.; Apostolopoulos, V.; Kalbacher, H.; Papini, A.-M.; Tselios, T.; Chatzantoni, K.; Biagioli, T.; Lolli, F.; Deraos, S.; Papathanassopoulos, P.; et al. Design And Synthesis of a Novel Potent Myelin Basic Protein Epitope 87−99 Cyclic Analogue: Enhanced Stability and Biological Properties of Mimics Render Them a Potentially New Class of Immunomodulators. J. Med. Chem. 2005, 48, 1470–1480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deraos, G.; Rodi, M.; Kalbacher, H.; Chatzantoni, K.; Karagiannis, F.; Synodinos, L.; Plotas, P.; Papalois, A.; Dimisianos, N.; Papathanasopoulos, P.; et al. Properties of myelin altered peptide ligand cyclo(87-99)(Ala91,Ala96)MBP87-99 render it a promising drug lead for immunotherapy of multiple sclerosis. Eur. J. Med. Chem. 2015, 101, 13–23. [Google Scholar] [CrossRef]
- Deraos, G.; Chatzantoni, K.; Matsoukas, M.-T.; Tselios, T.; Deraos, S.; Katsara, M.; Papathanasopoulos, P.; Vynios, D.; Apostolopoulos, V.; Mouzaki, A.; et al. Citrullination of Linear and Cyclic Altered Peptide Ligands from Myelin Basic Protein (MBP87−99) Epitope Elicits a Th1 Polarized Response by T Cells Isolated from Multiple Sclerosis Patients: Implications in Triggering Disease. J. Med. Chem. 2008, 51, 7834–7842. [Google Scholar] [CrossRef]
- Deraos, G.; Kritsi, E.; Matsoukas, M.-T.; Christopoulou, K.; Kalbacher, H.; Zoumpoulakis, P.; Apostolopoulos, V.; Matsoukas, J. Design of Linear and Cyclic Mutant Analogues of Dirucotide Peptide (MBP82–98) against Multiple Sclerosis: Conformational and Binding Studies to MHC Class II. Brain Sci. 2018, 8, 213. [Google Scholar] [CrossRef] [Green Version]
- Katsara, M.; Yuriev, E.; Ramsland, P.A.; Deraos, G.; Tselios, T.; Matsoukas, J.; Apostolopoulos, V. Mannosylation of mutated MBP83–99 peptides diverts immune responses from Th1 to Th2. Mol. Immunol. 2008, 45, 3661–3670. [Google Scholar] [CrossRef]
- Tseveleki, V.; Tselios, T.; Kanistras, I.; Koutsoni, O.; Karamita, M.; Vamvakas, S.-S.; Apostolopoulos, V.; Dotsika, E.; Matsoukas, J.; Lassmann, H.; et al. Mannan-conjugated myelin peptides prime non-pathogenic Th1 and Th17 cells and ameliorate experimental autoimmune encephalomyelitis. Exp. Neurol. 2015, 267, 254–267. [Google Scholar] [CrossRef] [Green Version]
- Dagkonaki, A.; Avloniti, M.; Evangelidou, M.; Papazian, I.; Kanistras, I.; Tseveleki, V.; Lampros, F.; Tselios, T.; Jensen, L.T.; Möbius, W.; et al. Mannan-MOG35-55 Reverses Experimental Autoimmune Encephalomyelitis, Inducing a Peripheral Type 2 Myeloid Response, Reducing CNS Inflammation, and Preserving Axons in Spinal Cord Lesions. Front. Immunol. 2020, 11. [Google Scholar] [CrossRef]
- Katsara, M.; Deraos, G.; Tselios, T.; Matsoukas, J.; Apostolopoulos, V. Design of Novel Cyclic Altered Peptide Ligands of Myelin Basic Protein MBP83−99That Modulate Immune Responses in SJL/J Mice. J. Med. Chem. 2008, 51, 3971–3978. [Google Scholar] [CrossRef]
- Moharregh-Khiabani, D.; Linker, R.; Gold, R.; Stangel, M. Fumaric Acid and its Esters: An Emerging Treatment for Multiple Sclerosis. Curr. Neuropharmacol. 2009, 7, 60–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zécri, F.J. From Natural Product to the First Oral Treatment for Multiple Sclerosis: The Discovery of FTY720 (Gilenya™)? Curr. Opin. Chem. Biol. 2016, 32, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Berger, A.A.; Sottosanti, E.R.; Winnick, A.; Izygon, J.; Berardino, K.; Cornett, E.M.; Kaye, A.D.; Varrassi, G.; Viswanath, O.; Urits, I. Monomethyl Fumarate (MMF, Bafiertam) for the Treatment of Relapsing Forms of Multiple Sclerosis (MS). Neurol. Int. 2021, 13, 207–223. [Google Scholar] [CrossRef] [PubMed]
- Yannakakis, M.-P.; Simal, C.; Tzoupis, H.; Rodi, M.; Dargahi, N.; Prakash, M.; Mouzaki, A.; Platts, J.; Apostolopoulos, V.; Tselios, T. Design and Synthesis of Non-Peptide Mimetics Mapping the Immunodominant Myelin Basic Protein (MBP83–96) Epitope to Function as T-Cell Receptor Antagonists. Int. J. Mol. Sci. 2017, 18, 1215. [Google Scholar] [CrossRef] [PubMed]
- Rayatpour, A.; Javan, M. Targeting the brain lesions using peptides: A review focused on the possibility of targeted drug delivery to multiple sclerosis lesions. Pharmacol. Res. 2021, 167, 105441. [Google Scholar] [CrossRef]
- Sestak, J.O.; Fakhari, A.; Badawi, A.H.; Siahaan, T.J.; Berkland, C. Structure, Size, and Solubility of Antigen Arrays Determines Efficacy in Experimental Autoimmune Encephalomyelitis. AAPS J. 2014, 16, 1185–1193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeste, A.; Nadeau, M.; Burns, E.J.; Weiner, H.L.; Quintana, F.J. Nanoparticle-mediated codelivery of myelin antigen and a tolerogenic small molecule suppresses experimental autoimmune encephalomyelitis. Proc. Natl. Acad. Sci. USA 2012, 109, 11270–11275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, Y.; Li, Z.; Zhu, H.; Gu, Z.; Zhang, H.; Luo, K. Recent Advances in Nanomedicines for Multiple Sclerosis Therapy. ACS Appl. Bio Mater. 2020, 3, 6571–6597. [Google Scholar] [CrossRef]
- Ahlawat, J.; Guillama Barroso, G.; Masoudi Asil, S.; Alvarado, M.; Armendariz, I.; Bernal, J.; Carabaza, X.; Chavez, S.; Cruz, P.; Escalante, V.; et al. Nanocarriers as Potential Drug Delivery Candidates for Overcoming the Blood–Brain Barrier: Challenges and Possibilities. ACS Omega 2020, 5, 12583–12595. [Google Scholar] [CrossRef]
- Warren, K.G.; Catz, I.; Ferenczi, L.Z.; Krantz, M.J. Intravenous synthetic peptide MBP8298 delayed disease progression in an HLA Class II-defined cohort of patients with progressive multiple sclerosis: Results of a 24-month double-blind placebo-controlled clinical trial and 5 years of follow-up treatment. Eur. J. Neurol. 2006, 13, 887–895. [Google Scholar] [CrossRef]
- Freedman, M.S.; Bar-Or, A.; Oger, J.; Traboulsee, A.; Patry, D.; Young, C.; Olsson, T.; Li, D.; Hartung, H.P.; Krantz, M.; et al. A phase III study evaluating the efficacy and safety of MBP8298 in secondary progressive MS. Neurology 2011, 77, 1551–1560. [Google Scholar] [CrossRef] [PubMed]
- Lourbopoulos, A.; Matsoukas, M.-T.; Katsara, M.; Deraos, G.; Giannakopoulou, A.; Lagoudaki, R.; Grigoriadis, N.; Matsoukas, J.; Apostolopoulos, V. Cyclization of PLP139-151 peptide reduces its encephalitogenic potential in experimental autoimmune encephalomyelitis. Bioorganic Med. Chem. 2018, 26, 2221–2228. [Google Scholar] [CrossRef] [PubMed]
- Katsara, M.; Tselios, T.; Deraos, S.; Deraos, G.; Matsoukas, M.T.; Lazoura, E.; Matsoukas, J.; Apostolopoulos, V. Round and round we go: Cyclic peptides in disease. Curr. Med. Chem. 2006, 13, 2221–2232. [Google Scholar] [CrossRef] [PubMed]
- Lourbopoulos, A.; Deraos, G.; Matsoukas, M.T.; Touloumi, O.; Giannakopoulou, A.; Kalbacher, H.; Grigoriadis, N.; Apostolopoulos, V.; Matsoukas, J. Cyclic MOG35-55 ameliorates clinical and neuropathological features of experimental autoimmune encephalomyelitis. Bioorg. Med. Chem. 2017, 25, 4163–4174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Apostolopoulos, V.; Barnes, N.; Pietersz, G.A.; McKenzie, I.F. Ex vivo targeting of the macrophage mannose receptor generates anti-tumor CTL responses. Vaccine 2000, 18, 3174–3184. [Google Scholar] [CrossRef]
- Sheng, K.C.; Kalkanidis, M.; Pouniotis, D.S.; Esparon, S.; Tang, C.K.; Apostolopoulos, V.; Pietersz, G.A. Delivery of antigen using a novel mannosylated dendrimer potentiates immunogenicity in vitro and in vivo. Eur. J. Immunol. 2008, 38, 424–436. [Google Scholar] [CrossRef] [PubMed]
- Sheng, K.C.; Kalkanidis, M.; Pouniotis, D.S.; Wright, M.D.; Pietersz, G.A.; Apostolopoulos, V. The adjuvanticity of a mannosylated antigen reveals TLR4 functionality essential for subset specialization and functional maturation of mouse dendritic cells. J. Immunol. 2008, 181, 2455–2464. [Google Scholar] [CrossRef] [Green Version]
- Sheng, K.C.; Pouniotis, D.S.; Wright, M.D.; Tang, C.K.; Lazoura, E.; Pietersz, G.A.; Apostolopoulos, V. Mannan derivatives induce phenotypic and functional maturation of mouse dendritic cells. Immunology 2006, 118, 372–383. [Google Scholar] [CrossRef]
- Tang, C.K.; Sheng, K.C.; Apostolopoulos, V.; Pietersz, G.A. Protein/peptide and DNA vaccine delivery by targeting C-type lectin receptors. Expert Rev. Vaccines 2008, 7, 1005–1018. [Google Scholar] [CrossRef]
- Tang, C.K.; Sheng, K.C.; Pouniotis, D.; Esparon, S.; Son, H.Y.; Kim, C.W.; Pietersz, G.A.; Apostolopoulos, V. Oxidized and reduced mannan mediated MUC1 DNA immunization induce effective anti-tumor responses. Vaccine 2008, 26, 3827–3834. [Google Scholar] [CrossRef]
- Acres, B.; Apostolopoulos, V.; Balloul, J.M.; Wreschner, D.; Xing, P.X.; Ali-Hadji, D.; Bizouarne, N.; Kieny, M.P.; McKenzie, I.F. MUC1-specific immune responses in human MUC1 transgenic mice immunized with various human MUC1 vaccines. Cancer Immunol. Immunother. 2000, 48, 588–594. [Google Scholar] [CrossRef] [PubMed]
- Apostolopoulos, V.; Lofthouse, S.A.; Popovski, V.; Chelvanayagam, G.; Sandrin, M.S.; McKenzie, I.F. Peptide mimics of a tumor antigen induce functional cytotoxic T cells. Nat. Biotechnol. 1998, 16, 276–280. [Google Scholar] [CrossRef]
- Apostolopoulos, V.; McKenzie, I.F. Role of the mannose receptor in the immune response. Curr. Mol. Med. 2001, 1, 469–474. [Google Scholar] [CrossRef] [PubMed]
- Apostolopoulos, V.; Pietersz, G.A.; Gordon, S.; Martinez-Pomares, L.; McKenzie, I.F. Aldehyde-mannan antigen complexes target the MHC class I antigen-presentation pathway. Eur. J. Immunol. 2000, 30, 1714–1723. [Google Scholar] [CrossRef]
- Apostolopoulos, V.; Pietersz, G.A.; Loveland, B.E.; Sandrin, M.S.; McKenzie, I.F. Oxidative/reductive conjugation of mannan to antigen selects for T1 or T2 immune responses. Proc. Natl. Acad. Sci. USA 1995, 92, 10128–10132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Apostolopoulos, V.; Pietersz, G.A.; McKenzie, I.F. Cell-mediated immune responses to MUC1 fusion protein coupled to mannan. Vaccine 1996, 14, 930–938. [Google Scholar] [CrossRef]
- Davis, W.C.; Konzek, R.L.; Haas, K.; Estes, D.M.; Hamilton, M.J.; Call, D.R.; Apostolopoulos, V.; McKenzie, I.F. Use of the mannan receptor to selectively target vaccine antigens for processing and antigen presentation through the MHC class I and class II pathways. Ann. N. Y. Acad. Sci. 2002, 969, 119–125. [Google Scholar] [CrossRef]
- Lees, C.J.; Apostolopoulos, V.; Acres, B.; Ong, C.S.; Popovski, V.; McKenzie, I.F. The effect of T1 and T2 cytokines on the cytotoxic T cell response to mannan-MUC1. Cancer Immunol. Immunother. 2000, 48, 644–652. [Google Scholar] [CrossRef]
- Lees, C.J.; Apostolopoulos, V.; Acres, B.; Ramshaw, I.; Ramsay, A.; Ong, C.S.; McKenzie, I.F. Immunotherapy with mannan-MUC1 and IL-12 in MUC1 transgenic mice. Vaccine 2000, 19, 158–162. [Google Scholar] [CrossRef]
- Lees, C.J.; Apostolopoulos, V.; McKenzie, I.F. Cytokine production from murine CD4 and CD8 cells after mannan-MUC1 immunization. J. Interferon Cytokine Res. 1999, 19, 1373–1379. [Google Scholar] [CrossRef]
- Lofthouse, S.A.; Apostolopoulos, V.; Pietersz, G.A.; Li, W.; McKenzie, I.F. Induction of T1 (cytotoxic lymphocyte) and/or T2 (antibody) responses to a mucin-1 tumour antigen. Vaccine 1997, 15, 1586–1593. [Google Scholar] [CrossRef]
- Tang, C.K.; Lodding, J.; Minigo, G.; Pouniotis, D.S.; Plebanski, M.; Scholzen, A.; McKenzie, I.F.; Pietersz, G.A.; Apostolopoulos, V. Mannan-mediated gene delivery for cancer immunotherapy. Immunology 2007, 120, 325–335. [Google Scholar] [CrossRef] [PubMed]
- Sandrin, M.S.; Vaughan, H.A.; Xing, P.X.; McKenzie, I.F. Natural human anti-Gal alpha(1,3)Gal antibodies react with human mucin peptides. Glycoconj. J. 1997, 14, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Vaughan, H.A.; Ho, D.W.; Karanikas, V.; Sandrin, M.S.; McKenzie, I.F.; Pietersz, G.A. The immune response of mice and cynomolgus monkeys to macaque mucin 1-mannan. Vaccine 2000, 18, 3297–3309. [Google Scholar] [CrossRef]
- Vaughan, H.A.; Ho, D.W.; Karanikas, V.A.; Ong, C.S.; Hwang, L.A.; Pearson, J.M.; McKenzie, I.F.; Pietersz, G.A. Induction of humoral and cellular responses in cynomolgus monkeys immunised with mannan-human MUC1 conjugates. Vaccine 1999, 17, 2740–2752. [Google Scholar] [CrossRef]
- Apostolopoulos, V.; Osinski, C.; McKenzie, I.F. MUC1 cross-reactive Gal alpha(1,3)Gal antibodies in humans switch immune responses from cellular to humoral. Nat. Med. 1998, 4, 315–320. [Google Scholar] [CrossRef] [PubMed]
- Apostolopoulos, V.; Pietersz, G.A.; Tsibanis, A.; Tsikkinis, A.; Drakaki, H.; Loveland, B.E.; Piddlesden, S.J.; Plebanski, M.; Pouniotis, D.S.; Alexis, M.N.; et al. Pilot phase III immunotherapy study in early-stage breast cancer patients using oxidized mannan-MUC1 [ISRCTN71711835]. Breast Cancer Res. 2006, 8, R27. [Google Scholar] [CrossRef]
- Apostolopoulos, V.; Pietersz, G.A.; Tsibanis, A.; Tsikkinis, A.; Stojanovska, L.; McKenzie, I.F.; Vassilaros, S. Dendritic cell immunotherapy: Clinical outcomes. Clin. Transl. Immunology 2014, 3, e21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karanikas, V.; Hwang, L.A.; Pearson, J.; Ong, C.S.; Apostolopoulos, V.; Vaughan, H.; Xing, P.X.; Jamieson, G.; Pietersz, G.; Tait, B.; et al. Antibody and T cell responses of patients with adenocarcinoma immunized with mannan-MUC1 fusion protein. J. Clin. Investig. 1997, 100, 2783–2792. [Google Scholar] [CrossRef] [Green Version]
- Karanikas, V.; Lodding, J.; Maino, V.C.; McKenzie, I.F. Flow cytometric measurement of intracellular cytokines detects immune responses in MUC1 immunotherapy. Clin. Cancer Res. 2000, 6, 829–837. [Google Scholar]
- Karanikas, V.; Thynne, G.; Mitchell, P.; Ong, C.S.; Gunawardana, D.; Blum, R.; Pearson, J.; Lodding, J.; Pietersz, G.; Broadbent, R.; et al. Mannan Mucin-1 Peptide Immunization: Influence of Cyclophosphamide and the Route of Injection. J. Immunother. 2001, 24, 172–183. [Google Scholar] [CrossRef] [PubMed]
- Loveland, B.E.; Zhao, A.; White, S.; Gan, H.; Hamilton, K.; Xing, P.X.; Pietersz, G.A.; Apostolopoulos, V.; Vaughan, H.; Karanikas, V.; et al. Mannan-MUC1-pulsed dendritic cell immunotherapy: A phase I trial in patients with adenocarcinoma. Clin. Cancer Res. 2006, 12, 869–877. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, P.L.; Quinn, M.A.; Grant, P.T.; Allen, D.G.; Jobling, T.W.; White, S.C.; Zhao, A.; Karanikas, V.; Vaughan, H.; Pietersz, G.; et al. A phase 2, single-arm study of an autologous dendritic cell treatment against mucin 1 in patients with advanced epithelial ovarian cancer. J. Immunother. Cancer 2014, 2, 16. [Google Scholar] [CrossRef] [PubMed]
- Prince, H.M.; Wall, D.M.; Ritchie, D.; Honemann, D.; Harrrison, S.; Quach, H.; Thompson, M.; Hicks, R.; Lau, E.; Davison, J.; et al. In vivo tracking of dendritic cells in patients with multiple myeloma. J. Immunother. 2008, 31, 166–179. [Google Scholar] [CrossRef] [PubMed]
- Vassilaros, S.; Tsibanis, A.; Tsikkinis, A.; Pietersz, G.A.; McKenzie, I.F.; Apostolopoulos, V. Up to 15-year clinical follow-up of a pilot Phase III immunotherapy study in stage II breast cancer patients using oxidized mannan-MUC1. Immunotherapy 2013, 5, 1177–1182. [Google Scholar] [CrossRef] [Green Version]
- Agnes, M.C.; Tan, A.; Jordens, R.; Geluk, A.; Roep, B.O.; Ottenhoff, T.; Drijfhout, J.W.; Koning, F. Strongly increased efficiency of altered peptide ligands by mannosylation. Int. Immunol. 1998, 10, 1299–1304. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Fang, H.; Hu, Y.; Wu, J.; Zhang, S.; Feng, Y.; Lin, L.; Tian, H.; Chen, X. Combining mannose receptor mediated nanovaccines and gene regulated PD-L1 blockade for boosting cancer immunotherapy. Bioact. Mater. 2022, 7, 167–180. [Google Scholar] [CrossRef] [PubMed]
- Mommaas, A.M.; Mulder, A.A.; Jordens, R.; Out, C.; Tan, M.C.; Cresswell, P.; Kluin, P.M.; Koning, F. Human epidermal Langerhans cells lack functional mannose receptors and a fully developed endosomal/lysosomal compartment for loading of HLA class II molecules. Eur. J. Immunol. 1999, 29, 571–580. [Google Scholar] [CrossRef]
- Motoyama, K.; Mitsuyasu, R.; Akao, C.; Abu, H., II; Sato, N.; Tanaka, T.; Higashi, T.; Arima, H. Potential Use of Thioalkylated Mannose-Modified Dendrimer (G3)/alpha-Cyclodextrin Conjugate as an NF-kappaB siRNA Carrier for the Treatment of Fulminant Hepatitis. Mol. Pharm. 2015, 12, 3129–3136. [Google Scholar] [CrossRef]
- Motoyama, K.; Mitsuyasu, R.; Akao, C.; Tanaka, T.; Ohyama, A.; Sato, N.; Higashi, T.; Arima, H. Design and evaluation of thioalkylated mannose-modified dendrimer (G3)/alpha-cyclodextrin conjugates as antigen-presenting cell-selective siRNA carriers. AAPS J. 2014, 16, 1298–1308. [Google Scholar] [CrossRef] [Green Version]
- Tan, M.C.; Mommaas, A.M.; Drijfhout, J.W.; Jordens, R.; Onderwater, J.J.; Verwoerd, D.; Mulder, A.A.; van der Heiden, A.N.; Ottenhoff, T.H.; Cella, M.; et al. Mannose receptor mediated uptake of antigens strongly enhances HLA-class II restricted antigen presentation by cultured dendritic cells. Adv. Exp. Med. Biol. 1997, 417, 171–174. [Google Scholar] [CrossRef] [PubMed]
- Degano, M.; Garcia, K.C.; Apostolopoulos, V.; Rudolph, M.G.; Teyton, L.; Wilson, I.A. A functional hot spot for antigen recognition in a superagonist TCR/MHC complex. Immunity 2000, 12, 251–261. [Google Scholar] [CrossRef] [Green Version]
- Baka, Z.; Gyorgy, B.; Geher, P.; Buzas, E.I.; Falus, A.; Nagy, G. Citrullination under physiological and pathological conditions. Joint Bone Spine 2012, 79, 431–436. [Google Scholar] [CrossRef] [PubMed]
- Bradford, C.M.; Ramos, I.; Cross, A.K.; Haddock, G.; McQuaid, S.; Nicholas, A.P.; Woodroofe, M.N. Localisation of citrullinated proteins in normal appearing white matter and lesions in the central nervous system in multiple sclerosis. J. Neuroimmunol. 2014, 273, 85–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horibata, S.; Coonrod, S.A.; Cherrington, B.D. Role for peptidylarginine deiminase enzymes in disease and female reproduction. J. Reprod. Dev. 2012, 58, 274–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohanan, S.; Cherrington, B.D.; Horibata, S.; McElwee, J.L.; Thompson, P.R.; Coonrod, S.A. Potential role of peptidylarginine deiminase enzymes and protein citrullination in cancer pathogenesis. Biochem. Res. Int. 2012, 2012, 895343. [Google Scholar] [CrossRef] [Green Version]
- Pritzker, L.B.; Joshi, S.; Gowan, J.J.; Harauz, G.; Moscarello, M.A. Deimination of myelin basic protein. 1. Effect of deimination of arginyl residues of myelin basic protein on its structure and susceptibility to digestion by cathepsin D. Biochemistry 2000, 39, 5374–5381. [Google Scholar] [CrossRef] [PubMed]
- Pritzker, L.B.; Joshi, S.; Harauz, G.; Moscarello, M.A. Deimination of myelin basic protein. 2. Effect of methylation of MBP on its deimination by peptidylarginine deiminase. Biochemistry 2000, 39, 5382–5388. [Google Scholar] [CrossRef]
- Whitaker, J.N.; Kirk, K.A.; Herman, P.K.; Zhou, S.R.; Goodin, R.R.; Moscarello, M.A.; Wood, D.D. An immunochemical comparison of human myelin basic protein and its modified, citrullinated form, C8. J. Neuroimmunol. 1992, 36, 135–146. [Google Scholar] [CrossRef]
- Hohlfeld, R. Biotechnological agents for the immunotherapy of multiple sclerosis. Principles, problems and perspectives. Brain 1997, 120, 865–916. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide Analog [Reference] | Major Effects |
|---|---|
| MBP83–99 and PLP139–151 [10,11,12,13,14,15,16,17,18] | These agonist peptides are involved in the pathophysiology of MS and also induce EAE in animal models. |
| MBP82–98 [40,41] | Dirucotide in animal models inhibits disease and in early human clinical trials showed efficacy; however, the peptide did not meet primary endpoints in phase III-trials. |
| cyclic(87–99)[MBP87–99] [23] | Stimulates Th2 cytokines and inhibits EAE in mice. |
| MBP87–99(R91,A96), MBP87–99(A91,A96) [21] | Induces IL-4 and antagonizes IFNγ responses in mice. |
| MBP72–85 [22] | These agonist peptides induce EAE in mice and Th1 responses in humans. |
| MBP72–85(A79) [22] | Suppresses EAE in mice. |
| PLP139–151(L144, R147) [42] | Antagonizes PLP-specific T-clones in vitro. |
| cyclic-MBP82–98 | Exerts strong binding to the HLA-DR2 and lowers binding to the HLA-DR4 allele in vitro |
| cyclic-MBP87–99(A96) or (R91A96) [21,30] | Suppresses proliferation of CD4+ T cells and exerts IL-10 selectivity in vitro. Binds to HLA-DR4 and is stable to lysosomal enzymes and cathepsins B, D, and H. |
| cyclic-MOG35–55 [44] cyclic-PLP131–151 [42] linear and cyclic-MBP83–99(A91,A96) [27] Mannan-linear and cyclic-MBP83–99(A91,A96) [27] | Reduces EAE, demyelination, and chronic axonopathy in acute and chronic phases of EAE in mice. Low disease burden in regards to EAE in mice with minimal inflammatory, demyelinating, and axonopathic pathology compared to its linear counterpart. Decreases IFNγ responses in mice. Diverts the “bad” IFNγ to “good” IL-4 cytokine in mice. |
| Mannan-MOG35–55 [28] | Protects mice against EAE in prophylactic and therapeutic protocols, with oxidized-conjugated peptides giving the best results. |
| Cyclo(87–99)MBP87-99(A91,A96) [27,30] Mannan-cyclo(87–99)MBP87-99(A91,A96) [27,30] | Decreases Th1 responses. Shifts Th1 responses to Th2 responses. |
| MBP87–99[Cit91,A96,Cit97] [25] cyclic-MBP87–99[Cit91,A96,Cit97] [25] | Induces T-cell proliferation and IFNγ secretion in mice. Activates T cells and increases IFNγ secretion in mice. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matsoukas, J.M.; Ligielli, I.; Chasapis, C.T.; Kelaidonis, K.; Apostolopoulos, V.; Mavromoustakos, T. Novel Approaches in the Immunotherapy of Multiple Sclerosis: Cyclization of Myelin Epitope Peptides and Conjugation with Mannan. Brain Sci. 2021, 11, 1583. https://doi.org/10.3390/brainsci11121583
Matsoukas JM, Ligielli I, Chasapis CT, Kelaidonis K, Apostolopoulos V, Mavromoustakos T. Novel Approaches in the Immunotherapy of Multiple Sclerosis: Cyclization of Myelin Epitope Peptides and Conjugation with Mannan. Brain Sciences. 2021; 11(12):1583. https://doi.org/10.3390/brainsci11121583
Chicago/Turabian StyleMatsoukas, John M., Irene Ligielli, Christos T. Chasapis, Konstantinos Kelaidonis, Vasso Apostolopoulos, and Thomas Mavromoustakos. 2021. "Novel Approaches in the Immunotherapy of Multiple Sclerosis: Cyclization of Myelin Epitope Peptides and Conjugation with Mannan" Brain Sciences 11, no. 12: 1583. https://doi.org/10.3390/brainsci11121583