
Prefrontal Transcranial Direct Current Stimulation Globally Improves Learning but Does Not Selectively Potentiate the Benefits of Targeted Memory Reactivation on Awake Memory Consolidation


{kind=link}
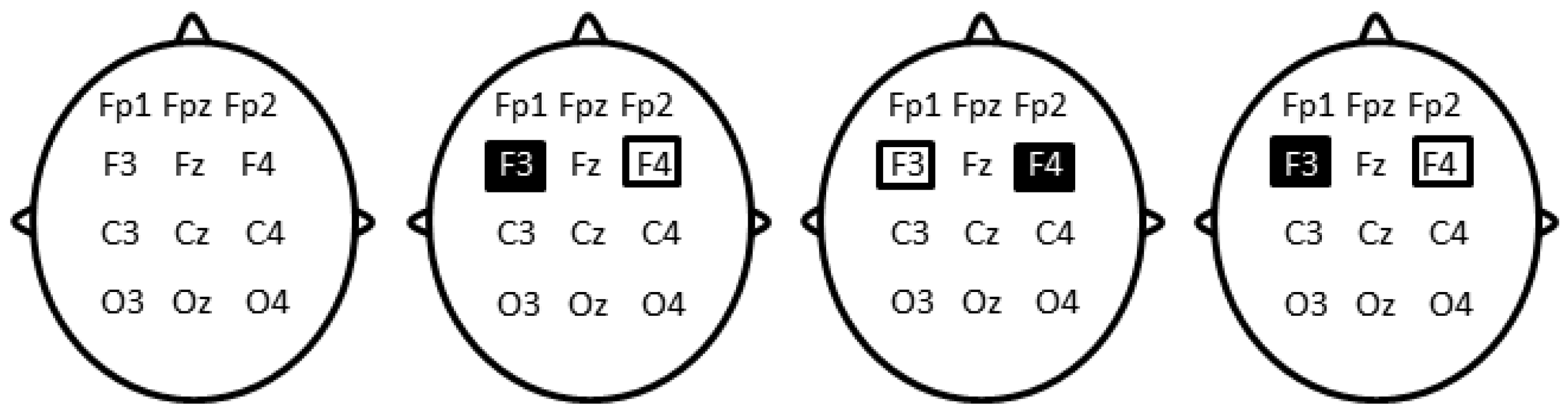{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Material
2.3. Procedure
2.4. Transcranial Direct Current Stimulation
3. Results
3.1. Sleepiness and Vigilance
3.2. Pre-Stimulation Learning Session (IRT)
3.3. Immediate Post-Stimulation Testing Session (RT1)
3.4. Long Term Memory Consolidation (RT2)
4. Discussion
4.1. The Benefits of Auditory Cueing on Memory Consolidation
4.2. The Effects of tDCS on Memory Consolidation
4.3. Consolidation of Emotional Memories and Lateralisation of tDCS Polarity
4.4. No Long-Term Benefits of TMR and tDCS
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mcgaugh, J.L. Time-dependent processes in memory storage. Science 1966, 153, 1351–1358. [Google Scholar] [CrossRef]
- Buzsaki, G. The hippocampo-neocortical dialogue. Cerebral Cortex 1996, 6, 81–92. [Google Scholar] [CrossRef] [Green Version]
- Peigneux, P.; Orban, P.; Balteau, E.; Degueldre, C.; Luxen, A.; Laureys, S.; Maquet, P. Offline persistence of memory-related cerebral activity during active wakefulness. PLoS Biol. 2006, 4, E100. [Google Scholar] [CrossRef]
- Tambini, A.; Davachi, L. Awake reactivation of prior experiences consolidates memories and biases cognition. Trends Cogn. Sci. 2019, 23, 876–890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wamsley, E.J. Memory consolidation during waking rest. Trends Cogn. Sci. 2019, 23, 171–173. [Google Scholar] [CrossRef] [PubMed]
- Humiston, G.B.; Tucker, M.A.; Summer, T.; Wamsley, E.J. Resting states and memory consolidation: A preregistered replication and meta-analysis. Sci. Rep. 2019, 9, 19345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sara, S.J. Reactivation, retrieval, replay and reconsolidation in and out of sleep: Connecting the dots. Front. Behav. Neurosci. 2010, 4, 185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skaggs, W.E.; Mcnaughton, B.L. Replay of neuronal firing sequences in rat hippocampus during sleep following spatial experience. Science 1996, 271, 1870–1873. [Google Scholar] [CrossRef]
- Valdes, J.L.; Mcnaughton, B.L.; Fellous, J.M. Offline reactivation of experience-dependent neuronal firing patterns in the rat ventral tegmental area. J. Neurophysiol. 2015, 114, 1183–1195. [Google Scholar] [CrossRef] [Green Version]
- Peigneux, P.; Laureys, S.; Fuchs, S.; Collette, F.; Perrin, F.; Reggers, J.; Phillips, C.; Degueldre, C.; Del Fiore, G.; Aerts, J.; et al. Are spatial memories strengthened in the human hippocampus during slow wave sleep? Neuron 2004, 44, 535–545. [Google Scholar] [CrossRef] [Green Version]
- Jegou, A.; Schabus, M.; Gosseries, O.; Dahmen, B.; Albouy, G.; Desseilles, M.; Sterpenich, V.; Phillips, C.; Maquet, P.; Grova, C.; et al. Cortical reactivations during sleep spindles following declarative learning. Neuroimage 2019, 195, 104–112. [Google Scholar] [CrossRef] [Green Version]
- Schonauer, M.; Alizadeh, S.; Jamalabadi, H.; Abraham, A.; Pawlizki, A.; Gais, S. Decoding material-specific memory reprocessing during sleep in humans. Nat. Commun. 2017, 8, 15404. [Google Scholar] [CrossRef] [Green Version]
- Diekelmann, S.; Born, J. The memory function of sleep. Nat. Rev. Neurosci. 2010, 11, 114–126. [Google Scholar] [CrossRef] [PubMed]
- Born, J.; Wilhelm, I. System consolidation of memory during sleep. Psychol. Res. 2012, 76, 192–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westermann, J.; Lange, T.; Textor, J.; Born, J. System consolidation during sleep—A common principle underlying psychological and immunological memory formation. Trends Neurosci. 2015, 38, 585–597. [Google Scholar] [CrossRef]
- Hu, X.; Cheng, L.Y.; Chiu, M.H.; Paller, K.A. Promoting memory consolidation during sleep: A meta-analysis of targeted memory reactivation. Psychol. Bull. 2020, 146, 218–244. [Google Scholar] [CrossRef]
- Craig, M.; Ottaway, G.; Dewar, M. Rest on it: Awake quiescence facilitates insight. Cortex 2018, 109, 205–214. [Google Scholar] [CrossRef]
- Dewar, M.; Alber, J.; Butler, C.; Cowan, N.; Della Sala, S. Brief wakeful resting boosts new memories over the long term. Psychol. Sci. 2012, 23, 955–960. [Google Scholar] [CrossRef] [PubMed]
- Dewar, M.; Garcia, Y.; Cowan, N.; Della Sala, S. Delaying interference enhances memory consolidation in amnesic patients. Neuropsychology 2009, 23, 627–634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tambini, A.; Ketz, N.; Davachi, L. Enhanced brain correlations during rest are related to memory for recent experiences. Neuron 2010, 65, 280–290. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.Y.; Baker, K.C.; Culbreth, J.L.; Tracy, O.; Arora, M.; Liu, T.; Morris, S.; Collins, M.B.; Wamsley, E.J. ‘Sleep-dependent’ memory consolidation? Brief periods of post-training rest and sleep provide an equivalent benefit for both declarative and procedural memory. Learn. Mem. 2021, 28, 195–203. [Google Scholar] [CrossRef]
- Buzsaki, G. Two-stage model of memory trace formation: A role for “noisy” brain states. Neuroscience 1989, 31, 551–570. [Google Scholar] [CrossRef]
- Brokaw, K.; Tishler, W.; Manceor, S.; Hamilton, K.; Gaulden, A.; Parr, E.; Wamsley, E.J. Resting state eeg correlates of memory consolidation. Neurobiol. Learn. Mem. 2016, 130, 17–25. [Google Scholar] [CrossRef]
- Oudiette, D.; Paller, K. Upgrading the sleeping brain with targeted memory reactivation. Trends Cogn. Sci. 2013, 17, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Farthouat, J.; Gilson, M.; Peigneux, P. New evidence for the necessity of a silent plastic period during sleep for a memory benefit of targeted memory reactivation. Sleep Spindl. Cortical Up States 2017, 1, 14–26. [Google Scholar] [CrossRef]
- Oudiette, D.; Antony, J.; Creery, J.; Paller, K. The role of memory reactivation during wakefulness and sleep in determining which memories endure. J. Neurosci. 2013, 33, 6672–6678. [Google Scholar] [CrossRef]
- Schreiner, T.; Rasch, B. Cueing vocabulary in awake subjects during the day has no effect on memory. Somnologie-Schlafforschung Und Schlafmed. 2015, 19, 133–140. [Google Scholar] [CrossRef]
- Wilhelm, I.; Schreiner, T.; Beck, J.; Rasch, B. No effect of targeted memory reactivation during sleep on retention of vocabulary in adolescents. Sci. Rep. 2020, 10, 4255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diekelmann, S.; Buchel, C.; Born, J.; Rasch, B. Labile or stable: Opposing consequences for memory when reactivated during waking and sleep. Nat. Neurosci 2011, 14, 381–386. [Google Scholar] [CrossRef]
- Rudoy, J.; Voss, J.; Westerberg, C.; Paller, K. Strengthening individual memories by reactivating them during sleep. Science 2009, 326, 1079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schreiner, T.; Rasch, B. Boosting vocabulary learning by verbal cueing during sleep. Cereb. Cortex 2015, 25, 4169–4179. [Google Scholar] [CrossRef] [Green Version]
- Lewis, P.A.; Bendor, D. How targeted memory reactivation promotes the selective strengthening of memories in sleep. Curr. Biol. 2019, 29, R906–R912. [Google Scholar] [CrossRef] [PubMed]
- Nitsche, M.A.; Paulus, W. Excitability changes induced in the human motor cortex by weak transcranial direct current stimulation. J. Physiol. 2000, 527 Pt. 3, 633–639. [Google Scholar] [CrossRef]
- Huang, Y.Z.; Lu, M.K.; Antal, A.; Classen, J.; Nitsche, M.; Ziemann, U.; Ridding, M.; Hamada, M.; Ugawa, Y.; Jaberzadeh, S.; et al. Plasticity induced by non-invasive transcranial brain stimulation: A position paper. Clin. Neurophysiol. 2017, 128, 2318–2329. [Google Scholar] [CrossRef] [PubMed]
- Kuo, M.F.; Nitsche, M.A. Effects of transcranial electrical stimulation on cognition. Clin. Eeg. Neurosci. 2012, 43, 192–199. [Google Scholar] [CrossRef] [PubMed]
- Brunoni, A.R.; Nitsche, M.A.; Bolognini, N.; Bikson, M.; Wagner, T.; Merabet, L.; Edwards, D.J.; Valero-Cabre, A.; Rotenberg, A.; Pascual-Leone, A.; et al. Clinical research with transcranial direct current stimulation (Tdcs): Challenges and future directions. Brain Stimul. 2012, 5, 175–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westphal, A.J.; Chow, T.E.; Ngoy, C.; Zuo, X.; Liao, V.; Storozuk, L.A.; Peters, M.A.K.; Wu, A.D.; Rissman, J. Anodal transcranial direct current stimulation to the left rostrolateral prefrontal cortex selectively improves source memory retrieval. J. Cogn. Neurosci. 2019, 31, 1380–1391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Javadi, A.H.; Walsh, V. Transcranial direct current stimulation (tdcs) of the left dorsolateral prefrontal cortex modulates declarative memory. Brain Stimul. 2012, 5, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Lang, N.; Nitsche, M.A.; Paulus, W.; Rothwell, J.C.; Lemon, R.N. Effects of transcranial direct current stimulation over the human motor cortex on corticospinal and transcallosal excitability. Exp. Brain Res. 2004, 156, 439–443. [Google Scholar] [CrossRef]
- Bjekic, J.; Vulic, K.; Zivanovic, M.; Vujicic, J.; Ljubisavljevic, M.; Filipovic, S.R. The immediate and delayed effects of single tdcs session over posterior parietal cortex on face-word associative memory. Behav. Brain Res. 2019, 366, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Bjekic, J.; Colic, M.V.; Zivanovic, M.; Milanovic, S.D.; Filipovic, S.R. Transcranial direct current stimulation (tdcs) over parietal cortex improves associative memory. Neurobiol. Learn. Mem. 2019, 157, 114–120. [Google Scholar] [CrossRef] [PubMed]
- Vulic, K.; Bjekic, J.; Paunovic, D.; Jovanovic, M.; Milanovic, S.; Filipovic, S.R. Theta-modulated oscillatory transcranial direct current stimulation over posterior parietal cortex improves associative memory. Sci. Rep. 2021, 11, 3013. [Google Scholar] [CrossRef]
- Sandrini, M.; Manenti, R.; Gobbi, E.; Rusich, D.; Bartl, G.; Cotelli, M. Transcranial direct current stimulation applied after encoding facilitates episodic memory consolidation in older adults. Neurobiol. Learn. Mem. 2019, 163, 107037. [Google Scholar] [CrossRef]
- Leach, R.C.; Mccurdy, M.P.; Trumbo, M.C.; Matzen, L.E.; Leshikar, E.D. Differential age effects of transcranial direct current stimulation on associative memory. J. Gerontol. B Psychol. Sci. Soc. Sci. 2019, 74, 1163–1173. [Google Scholar] [CrossRef]
- Floel, A.; Suttorp, W.; Kohl, O.; Kurten, J.; Lohmann, H.; Breitenstein, C.; Knecht, S. Non-invasive brain stimulation improves object-location learning in the elderly. Neurobiol. Aging 2012, 33, 1682–1689. [Google Scholar] [CrossRef] [PubMed]
- Nitsche, M.; Jakoubkova, M.; Thirugnanasambandam, N.; Schmalfuss, L.; Hullemann, S.; Sonka, K.; Paulus, W.; Trenkwalder, C.; Happe, S. Contribution of the premotor cortex to consolidation of motor sequence learning in humans during sleep. J. Neurophysiol. 2010, 104, 2603–2614. [Google Scholar] [CrossRef] [PubMed]
- Krause, V.; Meier, A.; Dinkelbach, L.; Pollok, B. Beta band transcranial alternating (tacs) and direct current stimulation (tdcs) applied after initial learning facilitate retrieval of a motor sequence. Front. Behav. Neurosci. 2016, 10, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galli, G.; Vadillo, M.A.; Sirota, M.; Feurra, M.; Medvedeva, A. A systematic review and meta-analysis of the effects of transcranial direct current stimulation (tdcs) on episodic memory. Brain Stimul. 2019, 12, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Deliens, G.; Gilson, M.; Peigneux, P. Sleep and the processing of emotions. Exp. Brain Res. 2014, 232, 1403–1414. [Google Scholar] [CrossRef]
- Borod, J.C.; Bloom, R.L.; Brickman, A.M.; Nakhutina, L.; Curko, E.A. Emotional processing deficits in individuals with unilateral brain damage. Appl. Neuropsychol. 2002, 9, 23–36. [Google Scholar] [CrossRef]
- Adolphs, R.; Jansari, A.; Tranel, D. Hemispheric perception of emotional valence from facial expressions. Neuropsychology 2001, 15, 516–524. [Google Scholar] [CrossRef]
- Balconi, M.; Ferrari, C. Repeated transcranial magnetic stimulation on dorsolateral prefrontal cortex improves performance in emotional memory retrieval as a function of level of anxiety and stimulus valence. Psychiatry Clin. Neurosci. 2013, 67, 210–218. [Google Scholar] [CrossRef] [PubMed]
- Borod, J.C.; Cicero, B.A.; Obler, L.K.; Welkowitz, J.; Erhan, H.M.; Santschi, C.; Grunwald, I.S.; Agosti, R.M.; Whalen, J.R. Right hemisphere emotional perception: Evidence across multiple channels. Neuropsychology 1998, 12, 446–458. [Google Scholar] [CrossRef] [PubMed]
- Faul, F.; Erdfelder, E.; Buchner, A.; Lang, A.G. Statistical power analyses using g*power 3.1: Tests for correlation and regression analyses. Behav. Res. Methods 2009, 41, 1149–1160. [Google Scholar] [CrossRef] [Green Version]
- Bagby, R.M.; Parker, J.D.A.; Taylor, G.J. The twenty-item toronto alexithymia scale-i. Item selection and cross-validation of the factor structure. J. Psychosom. Res. 1994, 38, 23–32. [Google Scholar] [CrossRef]
- Deltour, J.J. Echelle De Vocabulaire Mill Hill De J. C. Raven: Adaptation Française Et Normes Comparées Du Mill Hill Et Du Standard Progressive Matrices (Pm 38); Editions L’application Des Techniques Modernes: Braine-Le-Château, Belgium, 1993. [Google Scholar]
- Syssau, A.; Font, N. Evaluations des caractéristique émotionnelles d’un corpus de 604 mots. Bull. De Psychol. 2005, 58, 361–367. [Google Scholar] [CrossRef]
- Content, A.; Mousty, P.; Radeau, M. Brulex: Une base de données lexicales informatisée pour le français écrit et parlé. L’année Psychol. 1990, 90, 551–566. [Google Scholar] [CrossRef] [Green Version]
- Desrochers, A.; Bergeron, M. Valeurs de fréquence subjective et d’imagerie pour un échantillon de 1916 substantifs de la langue française. Revue Can. De Psychol. Expérimentale 2000, 54, 274–325. [Google Scholar] [CrossRef]
- Bradley, M.M.; Lang, P.J. Measuring emotion: The self-assessment manikin and the semantic differential. J. Behav. Ther. Exp. Psychiatry 1994, 25, 49–59. [Google Scholar] [CrossRef]
- Akerstedt, T.; Gillberg, M. Subjective and objective sleepiness in the active individual. Int. J. Neurosci. 1990, 52, 29–37. [Google Scholar] [CrossRef]
- Dinges, D.F.; Powell, J.W. Microcomputer analyses of per- formance on a portable, simple visual rt task during sustained op- erations. Behav. Res. Methods Instr. Comput. 1985, 17, 652–655. [Google Scholar] [CrossRef]
- Shin, Y.I.; Foerster, A.; Nitsche, M.A. Transcranial direct current stimulation (tdcs)—Application in neuropsychology. Neuropsychologia 2015, 69, 154–175. [Google Scholar] [CrossRef] [PubMed]
- Beam, W.; Borckardt, J.J.; Reeves, S.T.; George, M.S. An efficient and accurate new method for locating the f3 position for prefrontal tms applications. Brain Stimul. 2009, 2, 50–54. [Google Scholar] [CrossRef] [Green Version]
- Antal, A.; Alekseichuk, I.; Bikson, M.; Brockmoller, J.; Brunoni, A.R.; Chen, R.; Cohen, L.G.; Dowthwaite, G.; Ellrich, J.; Floel, A.; et al. Low intensity transcranial electric stimulation: Safety, ethical, legal regulatory and application guidelines. Clin. Neurophysiol. 2017, 128, 1774–1809. [Google Scholar] [CrossRef] [Green Version]
- Basner, M.; Dinges, D. Maximizing sensitivity of the psychomotor vigilance test (Pvt) to sleep loss. Sleep 2011, 34, 581–591. [Google Scholar] [CrossRef] [PubMed]
- Creery, J.D.; Oudiette, D.; Antony, J.W.; Paller, K.A. Targeted memory reactivation during sleep depends on prior learning. Sleep 2015, 38, 755–763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandrini, M.; Brambilla, M.; Manenti, R.; Rosini, S.; Cohen, L.G.; Cotelli, M. Noninvasive stimulation of prefrontal cortex strengthens existing episodic memories and reduces forgetting in the elderly. Front. Aging Neurosci. 2014, 6, 289. [Google Scholar] [CrossRef] [PubMed]
- Kirov, R.; Weiss, C.; Siebner, H.R.; Born, J.; Marshall, L. Slow oscillation electrical brain stimulation during waking promotes eeg theta activity and memory encoding. Proc. Natl. Acad. Sci. USA 2009, 106, 15460–15465. [Google Scholar] [CrossRef] [Green Version]
- Javadi, A.H.; Cheng, P. Transcranial direct current stimulation (tdcs) enhances reconsolidation of long-term memory. Brain Stimul. 2013, 6, 668–674. [Google Scholar] [CrossRef]
- Nitsche, M.A.; Fricke, K.; Henschke, U.; Schlitterlau, A.; Liebetanz, D.; Lang, N.; Henning, S.; Tergau, F.; Paulus, W. Pharmacological modulation of cortical excitability shifts induced by transcranial direct current stimulation in humans. J. Physiol. 2003, 553, 293–301. [Google Scholar] [CrossRef]
- Yavari, F.; Jamil, A.; Mosayebi Samani, M.; Vidor, L.P.; Nitsche, M.A. Basic and functional effects of transcranial electrical stimulation (tes)-an introduction. Neurosci. Biobehav. Rev. 2018, 85, 81–92. [Google Scholar] [CrossRef]
- Nitsche, M.A.; Paulus, W. Sustained excitability elevations induced by transcranial dc motor cortex stimulation in humans. Neurology 2001, 57, 1899–1901. [Google Scholar] [CrossRef] [PubMed]
- Liebetanz, D.; Nitsche, M.A.; Tergau, F.; Paulus, W. Pharmacological approach to the mechanisms of transcranial dc-stimulation-induced after-effects of human motor cortex excitability. Brain 2002, 125, 2238–2247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stagg, C.J.; Antal, A.; Nitsche, M.A. Physiology of transcranial direct current stimulation. J. Ect. 2018, 34, 144–152. [Google Scholar] [CrossRef]
- Wang, S.; Redondo, R.; Morris, R. Relevance of synaptic tagging and capture to the persistence of long-term potentiation and everyday spatial memory. Proc. Natl. Acad. Sci. USA 2010, 107, 19537–19542. [Google Scholar] [CrossRef] [Green Version]
- Polania, R.; Paulus, W.; Nitsche, M.A. Modulating cortico-striatal and thalamo-cortical functional connectivity with transcranial direct current stimulation. Hum. Brain Mapp. 2012, 33, 2499–2508. [Google Scholar] [CrossRef] [PubMed]
- Tulving, E.; Kapur, S.; Craik, F.; Moscovitch, M.; Houle, S. Hemispheric encoding/retrieval asymmetry in episodic memory: Positron emission tomography findings. Proc. Natl. Acad. Sci. USA 1994, 91, 2016–2020. [Google Scholar] [CrossRef] [Green Version]
- Cabeza, R.; Nyberg, L. Imaging cognition ii: An empirical review of 275 pet an fmri studies. J. Cogn. Neurosci. 2000, 12, 1–47. [Google Scholar] [CrossRef]
- Sandrini, M.; Censor, N.; Mishoe, J.; Cohen, L.G. Causal role of prefrontal cortex in strengthening of episodic memories through reconsolidation. Curr. Biol. 2013, 23, 2181–2184. [Google Scholar] [CrossRef] [Green Version]
- Nitsche, M.A.; Koschack, J.; Pohlers, H.; Hullemann, S.; Paulus, W.; Happe, S. Effects of frontal transcranial direct current stimulation on emotional state and processing in healthy humans. Front. Psychiatry 2012, 3, 58. [Google Scholar] [CrossRef] [Green Version]
- Penolazzi, B.; Di Domenico, A.; Marzoli, D.; Mammarella, N.; Fairfield, B.; Franciotti, R.; Brancucci, A.; Tommasi, L. Effects of transcranial direct current stimulation on episodic memory related to emotional visual stimuli. PLoS ONE 2010, 5, E10623. [Google Scholar] [CrossRef]
- Kilpatrick, L.; Cahill, L. Amygdala modulation of parahippocampal and frontal regions during emotionally influenced memory storage. Neuroimage 2003, 20, 2091–2099. [Google Scholar] [CrossRef]
- Ochsner, K.N. Are affective events richly recollected or simply familiar? The experience and process of recognizing feelings past. J. Exp. Psychol. Gen. 2000, 129, 242–261. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Lin, W.; Liu, C.; Luo, Y.; Wu, J.; Bayley, P.J.; Qin, S. Memory consolidation reconfigures neural pathways involved in the suppression of emotional memories. Nat. Commun. 2016, 7, 13375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Voogd, L.D.; Fernandez, G.; Hermans, E.J. Awake reactivation of emotional memory traces through hippocampal-neocortical interactions. Neuroimage 2016, 134, 563–572. [Google Scholar] [CrossRef] [PubMed]
- Okada, G.; Okamoto, Y.; Kunisato, Y.; Aoyama, S.; Nishiyama, Y.; Yoshimura, S.; Onoda, K.; Toki, S.; Yamashita, H.; Yamawaki, S. The effect of negative and positive emotionality on associative memory: An fmri study. PLoS ONE 2011, 6, e24862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.; Liu, X.; An, W.; Yang, Y.; Wang, Y. Recognition memory of neutral words can be impaired by task-irrelevant emotional encoding contexts: Behavioral and electrophysiological evidence. Front. Hum. Neurosci. 2015, 9, 73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nashiro, K.; Mather, M. Effects of emotional arousal on memory binding in normal aging and alzheimer’s disease. Am. J. Psychol. 2011, 124, 301–312. [Google Scholar] [CrossRef] [Green Version]
- Guez, J.; Saar-Ashkenazy, R.; Mualem, L.; Efrati, M.; Keha, E. Negative emotional arousal impairs associative memory performance for emotionally neutral content in healthy participants. PLoS ONE 2015, 10, e0132405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bisby, J.A.; Burgess, N. Negative affect impairs associative memory but not item memory. Learn. Mem. 2013, 21, 21–27. [Google Scholar] [CrossRef] [Green Version]
- Nader, K. Memory traces unbound. Trends Neurosci. 2003, 26, 65–72. [Google Scholar] [CrossRef]
- Crossman, M.; Bartl, G.; Soerum, R.; Sandrini, M. Effects of transcranial direct current stimulation over the posterior parietal cortex on episodic memory reconsolidation. Cortex 2019, 121, 78–88. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gilson, M.; Nitsche, M.A.; Peigneux, P. Prefrontal Transcranial Direct Current Stimulation Globally Improves Learning but Does Not Selectively Potentiate the Benefits of Targeted Memory Reactivation on Awake Memory Consolidation. Brain Sci. 2021, 11, 1104. https://doi.org/10.3390/brainsci11081104
Gilson M, Nitsche MA, Peigneux P. Prefrontal Transcranial Direct Current Stimulation Globally Improves Learning but Does Not Selectively Potentiate the Benefits of Targeted Memory Reactivation on Awake Memory Consolidation. Brain Sciences. 2021; 11(8):1104. https://doi.org/10.3390/brainsci11081104
Chicago/Turabian StyleGilson, Médhi, Michael A. Nitsche, and Philippe Peigneux. 2021. "Prefrontal Transcranial Direct Current Stimulation Globally Improves Learning but Does Not Selectively Potentiate the Benefits of Targeted Memory Reactivation on Awake Memory Consolidation" Brain Sciences 11, no. 8: 1104. https://doi.org/10.3390/brainsci11081104