
Green Leaf Volatile Function in Both the Natural Defense System of Rumex confertus and Associated Insects’ Behavior
 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plants and Z-Jasmone (ZJA) and Dihydrojasmone (DJA) Application
2.2. Rumex Confertus Volatile Organic Compounds Collection and Analysis
2.3. Insects
2.4. Preparation and Application of Green Leaf Volatiles Organic Compounds Blends
2.5. Evaluation of the Influence of Synthetic GLVs on Insects Behavior with the Use of Y-Tube Olfactometer
2.6. Statistical Analysis
3. Results
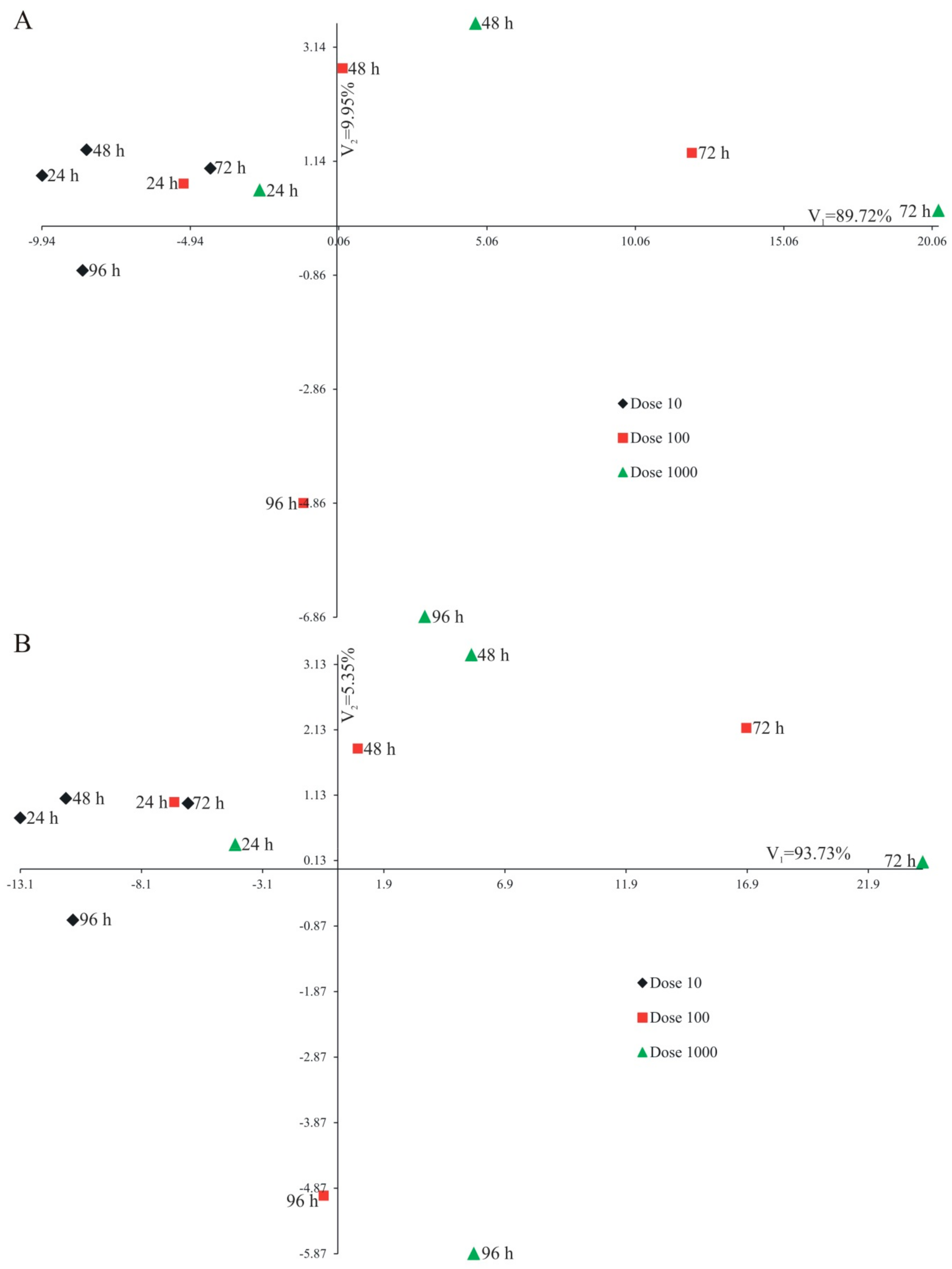
3.1. Plant Volatile Emissions Caused by ZJA and DJA Applications
3.2. Apion Miniatum Behavior in Response to Feed or Reproduction Stimuli
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Weber, E. Invasive Plant Species of the World: A Reference Guide to Environmental Weeds; Cabi: Wallingford, UK, 2017. [Google Scholar]
- Weber, E.; Gut, D. Assessing the Risk of Potentially Invasive Plant Species in Central Europe. J. Nat. Conserv. 2004, 12, 171–179. [Google Scholar] [CrossRef]
- Kołodziejek, J.; Patykowski, J. Effect of Environmental Factors on Germination and Emergence of Invasive Rumex Confertus in Central Europe. Sci. World J. 2015, 2015, 170176. [Google Scholar] [CrossRef] [PubMed]
- Raycheva, T. Rumex Confertus (Polygonaceae) in the Bulgarian Flora. Bot. Serbica 2011, 35, 55–59. [Google Scholar]
- Rutkovska, S.; Pučka, I.; Novicka, I.; Evarts-bunders, P. Relationship of Geographic Distribution of the Most Characteristical Invasive Plant Species in Habitats Adjacent To the River Daugava Within the Territory of Daugavpils City. Acta Biol. Univ. Daugavp. 2011, 11, 163–175. [Google Scholar]
- USDA Weed Risk Assessment for Rumex Confertus Willd. (Polygonaceae)—Russian Dock; United States Department of Agriculture: Washington, DC, USA, 2016. [Google Scholar]
- Vasas, A.; Orbán-Gyapai, O.; Hohmann, J. The Genus Rumex: Review of Traditional Uses, Phytochemistry and Pharmacology. J. Ethnopharmacol. 2015, 175, 198–228. [Google Scholar] [CrossRef]
- Kołodziejek, J. Growth Performance and Emergence of Invasive Alien Rumex Confertus in Different Soil Types. Sci. Rep. 2019, 9, 19678. [Google Scholar] [CrossRef]
- Cloyd, R.A. How Effective Is Conservation Biological Control in Regulating Insect Pest Populations in Organic Crop Production Systems? Insects 2020, 11, 744. [Google Scholar] [CrossRef]
- Piesik, D. Uszkodzenia Powodowane Przez Owady Zasiedlające Rumex Confertus Willd. Acta Sci. Pol. Agric. 2004, 3, 247–256. [Google Scholar]
- Piesik, D.; Wenda-Piesik, A. Gastroidea Viridula Deg. Potential to Control Mossy Sorrel (Rumex Confertus Willd.). J. Plant Prot. Res. 2005, 45. [Google Scholar]
- Xie, Y.; Tian, L.; Han, X.; Yang, Y. Research Advances in Allelopathy of Volatile Organic Compounds (VOCs) of Plants. Horticulturae 2021, 7, 278. [Google Scholar] [CrossRef]
- Rosenkranz, M.; Chen, Y.; Zhu, P.; Vlot, A.C. Volatile Terpenes—Mediators of Plant-to-Plant Communication. Plant J. 2021, 108, 617–631. [Google Scholar] [CrossRef]
- Ameye, M.; Allmann, S.; Verwaeren, J.; Smagghe, G.; Haesaert, G.; Schuurink, R.C.; Audenaert, K. Green Leaf Volatile Production by Plants: A Meta-Analysis. New Phytol. 2018, 220, 666–683. [Google Scholar] [CrossRef]
- Bouwmeester, H.; Schuurink, R.C.; Bleeker, P.M.; Schiestl, F. The Role of Volatiles in Plant Communication. Plant J. 2019, 100, 892–907. [Google Scholar] [CrossRef]
- Gaffke, A.M.; Alborn, H.T.; Dudley, T.L.; Bean, D.W. Using Chemical Ecology to Enhance Weed Biological Control. Insects 2021, 12, 695. [Google Scholar] [CrossRef]
- Sharifi, R.; Ryu, C.M. Social Networking in Crop Plants: Wired and Wireless Cross-Plant Communications. Plant Cell Environ. 2021, 44, 1095–1110. [Google Scholar] [CrossRef]
- Hammerbacher, A.; Coutinho, T.A.; Gershenzon, J. Roles of Plant Volatiles in Defence against Microbial Pathogens and Microbial Exploitation of Volatiles. Plant Cell Environ. 2019, 42, 2827–2843. [Google Scholar] [CrossRef]
- Hu, L.; Zhang, K.; Wu, Z.; Xu, J.; Erb, M. Plant Volatiles as Regulators of Plant Defense and Herbivore Immunity: Molecular Mechanisms and Unanswered Questions. Curr. Opin. Insect Sci. 2021, 44, 82–88. [Google Scholar] [CrossRef]
- Bertea, C.M.; Casacci, L.; Bonelli, S.P.; Zampollo, A.; Barbero, F. Chemical, Physiological and Molecular Responses of Host Plants to Lepidopteran Egg-Laying. Front. Plant Sci. 2020, 10, 1768. [Google Scholar] [CrossRef]
- Lev-Yadun, S. Avoiding Rather than Resisting Herbivore Attacks Is Often the First Line of Plant Defence. Biol. J. Linn. Soc. 2021, 134, 775–802. [Google Scholar] [CrossRef]
- Gosset, V.; Harmel, N.; Göbel, C.; Francis, F.; Haubruge, E.; Wathelet, J.-P.; Du Jardin, P.; Feussner, I.; Fauconnier, M.-L. Attacks by a Piercing-Sucking Insect (Myzus Persicae Sultzer) or a Chewing Insect (Leptinotarsa Decemlineata Say) on Potato Plants (Solanum Tuberosum L.) Induce Differential Changes in Volatile Compound Release and Oxylipin Synthesis. J. Exp. Bot. 2009, 60, 1231–1240. [Google Scholar] [CrossRef]
- Li, T. Neighbour Recognition Through Volatile-Mediated Interactions. In Deciphering Chemical Language of Plant Communication; Blande, J.D., Glinwood, R., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 153–174. ISBN 978-3-319-33498-1. [Google Scholar]
- Guo, H.; Wang, C.-Z. The Ethological Significance and Olfactory Detection of Herbivore-Induced Plant Volatiles in Interactions of Plants, Herbivorous Insects, and Parasitoids. Arthropod. Plant. Interact. 2019, 13, 161–179. [Google Scholar] [CrossRef]
- Turlings, T.C.J.; Erb, M. Tritrophic Interactions Mediated by Herbivore-Induced Plant Volatiles: Mechanisms, Ecological Relevance, and Application Potential. Annu. Rev. Entomol. 2018, 63, 433–452. [Google Scholar] [CrossRef] [PubMed]
- Shao, X.; Cheng, K.; Wang, Z.; Zhang, Q.; Yang, X. Use of Odor by Host-Finding Insects: The Role of Real-Time Odor Environment and Odor Mixing Degree. Chemoecology 2021, 31, 149–158. [Google Scholar] [CrossRef]
- Sidorova, D.E.; Plyuta, V.A.; Padiy, D.A.; Kupriyanova, E.V.; Roshina, N.V.; Koksharova, O.A.; Khmel, I.A. The Effect of Volatile Organic Compounds on Different Organisms: Agrobacteria, Plants and Insects. Microorganisms 2022, 10, 69. [Google Scholar] [CrossRef] [PubMed]
- Thoming, G. Behavior Matters—Future Need for Insect Studies on Odor-Mediated Host Plant Recognition with the Aim of Making Use of Allelochemicals for Plant Protection. J. Agric. Food Chem. 2021, 69, 10469–10479. [Google Scholar] [CrossRef]
- Moreira, X.; Abdala-Roberts, L. Specificity and Context-Dependency of Plant–Plant Communication in Response to Insect Herbivory. Curr. Opin. Insect Sci. 2019, 32, 15–21. [Google Scholar] [CrossRef]
- Piesik, D.; Kalka, I.; Wenda-Piesik, A.; Bocianowski, J. Apion Miniatum Germ. Herbivory on the Mossy Sorrel, Rumex Confertus Willd.: Induced Plant Volatiles and Weevil Orientation Responses. Polish J. Environ. Stud. 2014, 23, 2149–2156. [Google Scholar] [CrossRef]
- Warzecha, T.; Skrzypek, E.; Bocianowski, J.; Sutkowska, A. Impact of Selected PSII Parameters on Barley DH Lines Biomass and Yield Elements. Agronomy 2021, 11, 1705. [Google Scholar] [CrossRef]
- Mahalanobis, P.C. On the Generalised Distance in Statistics. Proc. Natl. Inst. Sci. India 1936, 12, 49–55. [Google Scholar]
- Seidler-Łożykowska, K.; Bocianowski, J. Evaluation of Variability of Morphological Traits of Selected Caraway (Carum Carvi L.) Genotypes. Ind. Crops Prod. 2012, 35, 140–145. [Google Scholar] [CrossRef]
- Bocianowski, J.; Liersch, A. Multidimensional Analysis of Diversity in Genotypes of Winter Oilseed Rape (Brassica Napus L.). Agronomy 2022, 12, 633. [Google Scholar] [CrossRef]
- Ayelo, P.M.; Pirk, C.W.W.; Yusuf, A.A.; Chailleux, A.; Mohamed, S.A.; Deletre, E. Exploring the Kairomone-Based Foraging Behaviour of Natural Enemies to Enhance Biological Control: A Review. Front. Ecol. Evol. 2021, 9, 641974. [Google Scholar] [CrossRef]
- Cusumano, A.; Harvey, J.A.; Bourne, M.E.; Poelman, E.H.; De Boer, J. Exploiting Chemical Ecology to Manage Hyperparasitoids in Biological Control of Arthropod Pests. Pest Manag. Sci. 2020, 76, 432–443. [Google Scholar] [CrossRef]
- Piesik, D.; Wenda-piesik, A.; Kotwica, K.; Łyszczarz, A.; Delaney, K.J. Gastrophysa Polygoni Herbivory on Rumex Confertus: Single Leaf VOC Induction and Dose Dependent Herbivore Attraction/Repellence to Individual Compounds. J. Plant Physiol. 2011, 168, 2134–2138. [Google Scholar] [CrossRef]
- Piesik, D.; Bocianowski, J.; Sendel, S.; Krawczyk, K.; Kotwica, K. Beetle Orientation Responses of Gastrophysa Viridula and Gastrophysa Polygoni (Coleoptera: Chrysomelidae) to a Blend of Synthetic Volatile Organic Compounds. Environ. Entomol. 2020, 49, 1071–1076. [Google Scholar] [CrossRef]
- Piesik, D.; Wenda-Piesik, A.; Krasińska, A.; Wrzesińska, D.; Delaney, K.J. Volatile Organic Compounds Released by R Umex Confertus Following H Ypera Rumicis Herbivory and Weevil Responses to Volatiles. J. Appl. Entomol. 2016, 140, 308–316. [Google Scholar] [CrossRef]
- Kohout, V.; Kohoutova, S. Possibilities of Utilization of Species Apion Miniatum Germar in Biological Control of Genus Rumex. Sonderheft 1994, 14, 217–220. [Google Scholar]


{kind=link}
| VOCs | Z-3-HAL | Z-3-HOL | Z-3-HAC | Z-OCI | LIN | BAC | MAT | β-CAR | β-FAR |
|---|---|---|---|---|---|---|---|---|---|
| Z-3-HAL | |||||||||
| Z-3-HOL | 0.87 # | ||||||||
| Z-3-HAC | 0.87 | 0.87 | |||||||
| Z-OCI | 0.76 | 0.77 | 0.79 | ||||||
| LIN | 0.70 | 0.79 | 0.80 | 0.90 | |||||
| BAC | 0.65 | 0.71 | 0.66 | 0.80 | 0.78 | ||||
| MAT | 0.78 | 0.77 | 0.82 | 0.88 | 0.87 | 0.85 | |||
| B-CAR | 0.79 | 0.80 | 0.82 | 0.91 | 0.90 | 0.87 | 0.92 | ||
| B-FAR | 0.72 | 0.73 | 0.76 | 0.88 | 0.82 | 0.85 | 0.89 | 0.88 |
| VOCs | Z-3-HAL | E-2-HAL | Z-3-HOL | E-2-HOL | Z-3-HAC | Z-OCI | LIN | BAC | MAT | β-CAR | β-FAR |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Z-3-HAL | |||||||||||
| E-2-HAL | 0.93 # | ||||||||||
| Z-3-HOL | 0.89 | 0.90 | |||||||||
| E-2-HOL | 0.92 | 0.92 | 0.92 | ||||||||
| Z-3-HAC | 0.95 | 0.93 | 0.88 | 0.90 | |||||||
| Z-OCI | 0.79 | 0.79 | 0.84 | 0.79 | 0.77 | ||||||
| LIN | 0.80 | 0.77 | 0.78 | 0.78 | 0.77 | 0.91 | |||||
| BAC | 0.72 | 0.73 | 0.73 | 0.74 | 0.72 | 0.87 | 0.86 | ||||
| MAT | 0.83 | 0.81 | 0.83 | 0.81 | 0.80 | 0.85 | 0.85 | 0.91 | |||
| B-CAR | 0.84 | 0.84 | 0.84 | 0.84 | 0.82 | 0.89 | 0.86 | 0.89 | 0.90 | ||
| B-FAR | 0.74 | 0.76 | 0.79 | 0.77 | 0.73 | 0.88 | 0.86 | 0.88 | 0.86 | 0.94 |
| No. of Females | No. of Males | ||||||
|---|---|---|---|---|---|---|---|
| Name of Mixed Compounds | Dosage ng min−1 | + (2) | – (3) | χ2 (1) | + (2) | – (3) | χ2 (1) |
| (Z)-3-HAL | control (0.0) | 10 | 10 | 0.05 ns | 7 | 13 | 1.25 ns |
| + (Z)-3-HOL | 1 | 16 | 4 | 6.05 * (a) | 11 | 9 | 0.05 ns |
| + (Z)-3-HAC | 10 | 18 | 2 | 11.3 *** (a) | 16 | 4 | 6.05 * (a) |
| 100 | 13 | 7 | 1.25 ns | 10 | 10 | 0.05 ns | |
| 1000 | 1 | 19 | 14.45 *** (r) | 2 | 18 | 11.3 *** (r) | |
| No. of Females | No. of Males | ||||||
|---|---|---|---|---|---|---|---|
| Name of Mixed Compounds | Dosage ng min−1 | + (2) | – (3) | χ2 (1) | + (2) | – (3) | χ2 (1) |
| (Z)-3-HAL | control (0.0) | 11 | 9 | 0.05 ns | 9 | 11 | 0.05 ns |
| + (E)-2-HAL | 1 | 7 | 13 | 1.25 ns | 13 | 7 | 1.25 ns |
| + (Z)-3-HOL | 10 | 15 | 5 | 4.05 * (a) | 16 | 4 | 6.05 * (a) |
| + (E)-2-HOL | 100 | 17 | 3 | 8.45 ** (a) | 17 | 3 | 8.45 ** (a) |
| + (Z)-3-HAC | 1000 | 3 | 17 | 8.45 ** (r) | 3 | 17 | 8.45 ** (r) |
| No. of Unmated Females | No. of Unmated Males | ||||||
|---|---|---|---|---|---|---|---|
| Name of Mixed Compounds after ZJA Application | Dosage ng min−1 | + (2) B1 | – (3) Unmated Males | χ2 (1) | + (2) B1 | – (3) Unmated Females | χ2 (1) |
| (Z)-3-HAL | control (0.0) | 11 | 9 | 0.05 ns | 10 | 10 | 0.05 ns |
| + (Z)-3-HOL | 1 | 9 | 11 | 0.05 ns | 4 | 16 | 6.05 * |
| + (Z)-3-HAC | 10 | 15 | 5 | 4.05 * | 2 | 18 | 11.3 *** |
| No. of Mated Females | No. of Mated Males | ||||||
|---|---|---|---|---|---|---|---|
| Name of Mixed Compounds after ZJA Application | Dosage ng min−1 | + (2) B1 | – (3) Mated Males | χ2 (1) | + (2) B1 | – (3) Mated Females | χ2 (1) |
| (Z)-3-HAL | control (0.0) | 11 | 9 | 0.05 ns | 9 | 11 | 0.05 ns |
| + (Z)-3-HOL | 1 | 15 | 5 | 4.05 * | 4 | 16 | 6.05 * |
| + (Z)-3-HAC | 10 | 17 | 3 | 8.45 ** | 3 | 17 | 8.45 ** |
| No. of Unmated Females | No. of Unmated Males | ||||||
|---|---|---|---|---|---|---|---|
| Name of Mixed Compounds after DJA Application | Dosage ng min−1 | + (2) B2 | – (3) Unmated Males | χ2 (1) | + (2) B2 | – (3) Unmated Females | χ2 (1) |
| (Z)-3-HAL | control (0.0) | 13 | 7 | 1.25 ns | 11 | 9 | 0.05 ns |
| + (E)-2-HAL | 1 | 17 | 3 | 8.45 ** | 2 | 18 | 11.3 *** |
| + (Z)-3-HOL | 100 | 18 | 2 | 11.3 *** | 0 | 20 | 18.5 *** |
| + (E)-2-HOL | |||||||
| + (Z)-3-HAC | |||||||
| No. of Mated Females | No. of Mated Males | ||||||
|---|---|---|---|---|---|---|---|
| Name of Mixed Compounds after Dihydrojasmone Application | Dosage ng min−1 | + (2) B2 | – (3) Mated Males | χ2 (1) | + (2) B2 | – (3) Mated Females | χ2 (1) |
| (Z)-3-HAL | control (0.0) | 13 | 7 | 1.25 ns | 11 | 9 | 0.05 ns |
| + (E)-2-HAL | 1 | 16 | 4 | 6.05 * | 4 | 16 | 6.05 * |
| + (Z)-3-HOL | 100 | 20 | 0 | 18.5 *** | 2 | 18 | 11.3 *** |
| + (E)-2-HOL | |||||||
| + (Z)-3-HAC | |||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piesik, D.; Łyczko, J.; Krawczyk, K.; Gantner, M.; Bocianowski, J.; Ruzsanyi, V.; Mayhew, C.A. Green Leaf Volatile Function in Both the Natural Defense System of Rumex confertus and Associated Insects’ Behavior. Appl. Sci. 2023, 13, 2253. https://doi.org/10.3390/app13042253
Piesik D, Łyczko J, Krawczyk K, Gantner M, Bocianowski J, Ruzsanyi V, Mayhew CA. Green Leaf Volatile Function in Both the Natural Defense System of Rumex confertus and Associated Insects’ Behavior. Applied Sciences. 2023; 13(4):2253. https://doi.org/10.3390/app13042253
Chicago/Turabian StylePiesik, Dariusz, Jacek Łyczko, Krzysztof Krawczyk, Magdalena Gantner, Jan Bocianowski, Veronika Ruzsanyi, and Chris A. Mayhew. 2023. "Green Leaf Volatile Function in Both the Natural Defense System of Rumex confertus and Associated Insects’ Behavior" Applied Sciences 13, no. 4: 2253. https://doi.org/10.3390/app13042253