
Smoothness Harmonic: A Graph-Based Approach to Reveal Spatiotemporal Patterns of Cortical Dynamics in fMRI Data

Abstract
:Featured Application
Abstract
1. Introduction
2. Graph-Based Smoothness Harmonic
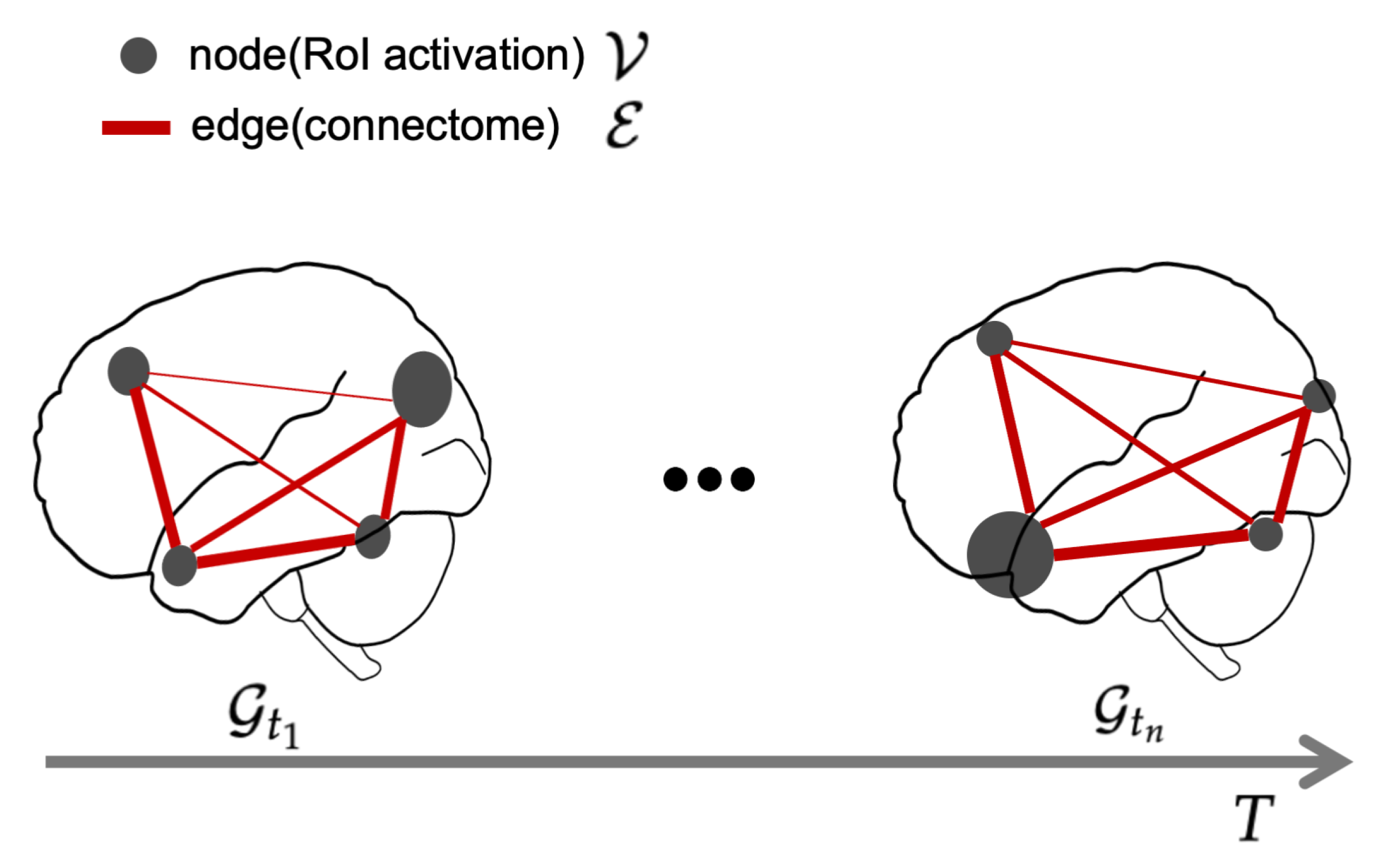
2.1. Time-Varying Brain Activity as a Graph
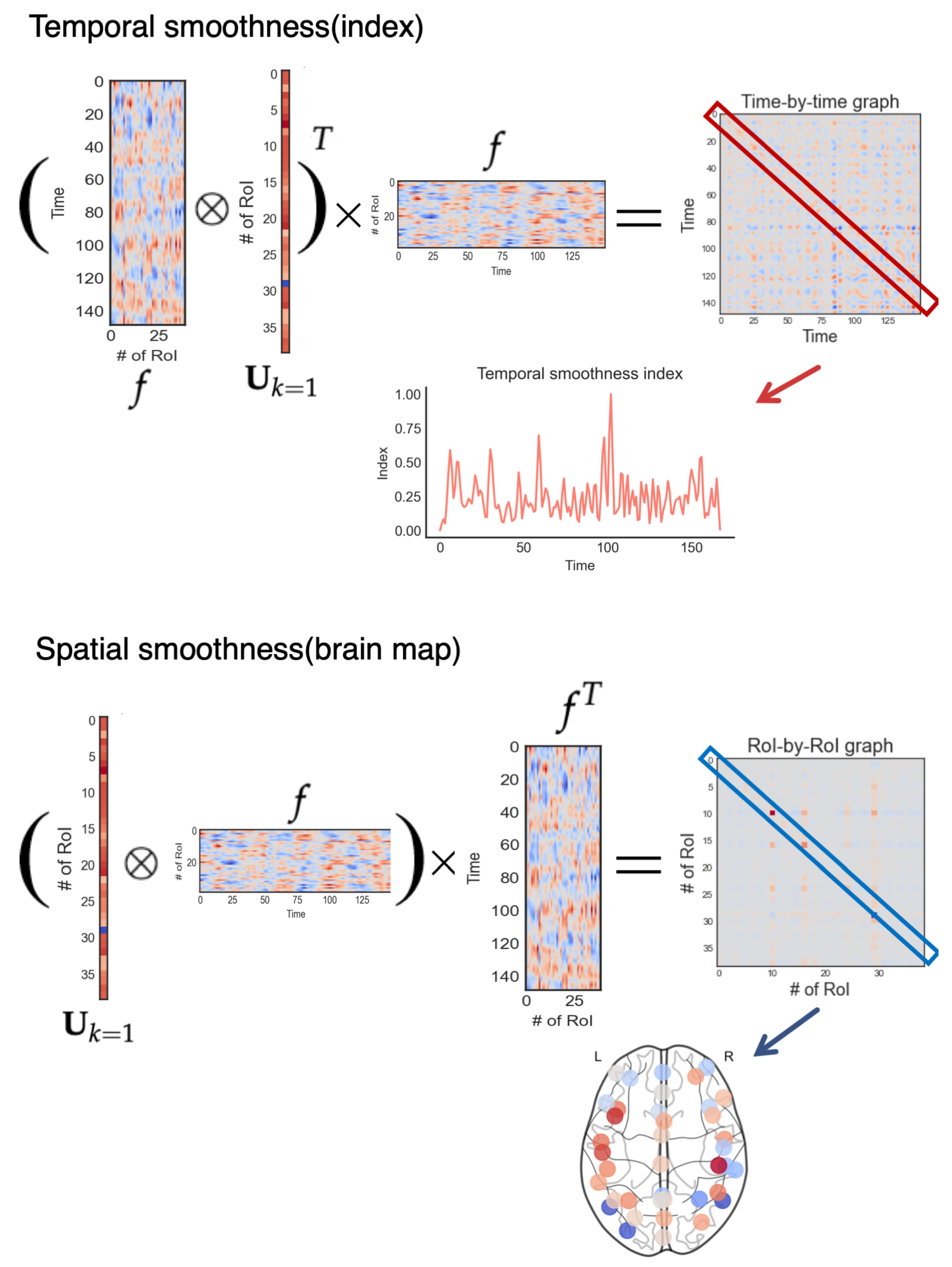
2.2. Smoothness Harmonic
2.3. Temporal Smoothness Index and Spatial Smoothness Brain Map
3. Application in Age-Development fMRI Data
3.1. Dataset and Preprocessing
3.2. f and Estimated
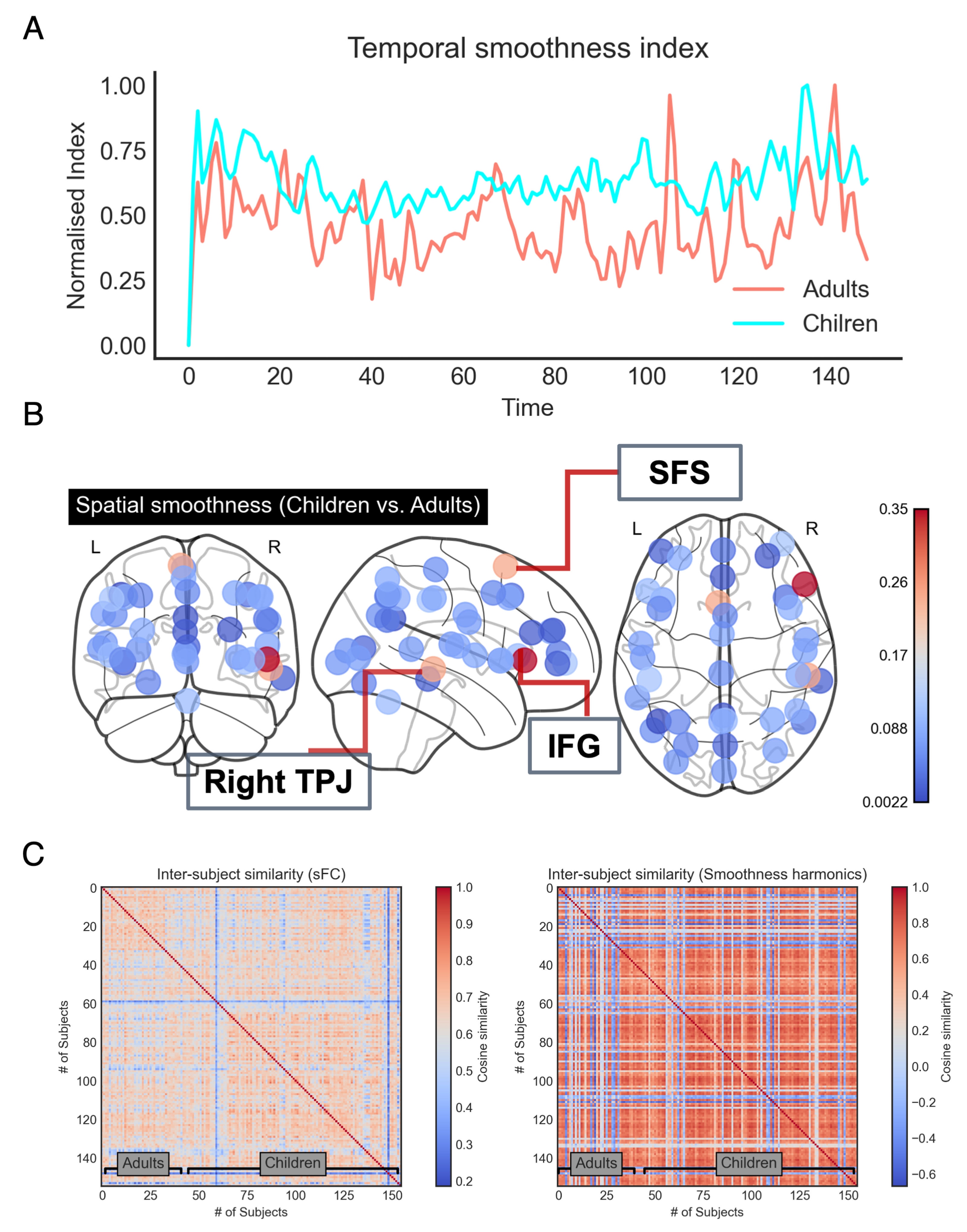
3.3. Extracted Smoothness Harmonics and Their Merits
4. Discussion
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Friston, K.J.; Kahan, J.; Razi, A.; Stephan, K.E.; Sporns, O. On nodes and modes in resting state fMRI. Neuroimage 2014, 99, 533–547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Logothetis, N.K. What we can do and what we cannot do with fMRI. Nature 2008, 453, 869–878. [Google Scholar] [CrossRef] [PubMed]
- Friston, K.J.; Holmes, A.P.; Poline, J.; Grasby, P.; Williams, S.; Frackowiak, R.S.; Turner, R. Analysis of fMRI time-series revisited. Neuroimage 1995, 2, 45–53. [Google Scholar] [CrossRef]
- Borogovac, A.; Asllani, I. Arterial spin labeling (ASL) fMRI: Advantages, theoretical constrains and experimental challenges in neurosciences. Int. J. Biomed. Imaging 2012, 2012, 818456. [Google Scholar]
- Niazy, R.K.; Xie, J.; Miller, K.; Beckmann, C.F.; Smith, S.M. Spectral characteristics of resting state networks. In Progress in Brain Research; Elsevier: Amsterdam, The Netherlands, 2011; Volume 13, pp. 259–276. [Google Scholar]
- Bullmore, E.; Sporns, O. Complex brain networks: Graph theoretical analysis of structural and functional systems. Nat. Rev. Neurosci. 2009, 10, 186–198. [Google Scholar] [CrossRef] [PubMed]
- Mears, D.; Pollard, H.B. Network science and the human brain: Using graph theory to understand the brain and one of its hubs, the amygdala, in health and disease. J. Neurosci. Res. 2016, 94, 590–605. [Google Scholar] [CrossRef]
- Telesford, Q.K.; Simpson, S.L.; Burdette, J.H.; Hayasaka, S.; Laurienti, P.J. The brain as a complex system: Using network science as a tool for understanding the brain. Brain Connect. 2011, 1, 295–308. [Google Scholar] [CrossRef] [Green Version]
- Farahani, F.V.; Karwowski, W.; Lighthall, N.R. Application of graph theory for identifying connectivity patterns in human brain networks: A systematic review. Front. Neurosci. 2019, 13, 585. [Google Scholar] [CrossRef]
- Grohe, M. word2vec, node2vec, graph2vec, x2vec: Towards a theory of vector embeddings of structured data. In Proceedings of the 39th ACM SIGMOD-SIGACT-SIGAI Symposium on Principles of Database Systems, Portland, OR, USA, 14–19 June 2020; pp. 1–16. [Google Scholar]
- Levakov, G.; Faskowitz, J.; Avidan, G.; Sporns, O. Mapping individual differences across brain network structure to function and behavior with connectome embedding. Neuroimage 2021, 242, 118469. [Google Scholar] [CrossRef]
- Narayanan, A.; Chandramohan, M.; Venkatesan, R.; Chen, L.; Liu, Y.; Jaiswal, S. graph2vec: Learning distributed representations of graphs. arXiv 2017, arXiv:1707.05005. [Google Scholar]
- Levy, B. Laplace-beltrami eigenfunctions towards an algorithm that “understands” geometry. In Proceedings of the IEEE International Conference on Shape Modeling and Applications 2006 (SMI’06), Matsushima, Japan, 14 June 2006; p. 13. [Google Scholar]
- Richardson, H.; Lisandrelli, G.; Riobueno-Naylor, A.; Saxe, R. Development of the social brain from age three to twelve years. Nat. Commun. 2018, 9, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Preti, M.G.; Bolton, T.A.; Van De Ville, D. The dynamic functional connectome: State-of-the-art and perspectives. Neuroimage 2017, 160, 41–54. [Google Scholar] [CrossRef] [PubMed]
- Leonardi, N.; Van De Ville, D. On spurious and real fluctuations of dynamic functional connectivity during rest. Neuroimage 2015, 104, 430–436. [Google Scholar] [CrossRef] [Green Version]
- Atasoy, S.; Donnelly, I.; Pearson, J. Human brain networks function in connectome-specific harmonic waves. Nat. Commun. 2016, 7, 10340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, X.; Thanou, D.; Frossard, P.; Vandergheynst, P. Learning Laplacian matrix in smooth graph signal representations. IEEE Trans. Signal Process. 2016, 64, 6160–6173. [Google Scholar] [CrossRef] [Green Version]
- Stankovic, L.; Mandic, D.; Dakovic, M.; Brajovic, M.; Scalzo, B.; Constantinides, T. Graph Signal Processing—Part I: Graphs, Graph Spectra, and Spectral Clustering. arXiv 2019, arXiv:1907.03467. [Google Scholar]
- Belkin, M.; Niyogi, P. Laplacian eigenmaps for dimensionality reduction and data representation. Neural Comput. 2003, 15, 1373–1396. [Google Scholar] [CrossRef] [Green Version]
- Buzsaki, G.; Draguhn, A. Neuronal oscillations in cortical networks. Science 2004, 304, 1926–1929. [Google Scholar] [CrossRef] [Green Version]
- Vanderwal, T.; Kelly, C.; Eilbott, J.; Mayes, L.C.; Castellanos, F.X. Inscapes: A movie paradigm to improve compliance in functional magnetic resonance imaging. Neuroimage 2015, 122, 222–232. [Google Scholar] [CrossRef] [Green Version]
- Haxby, J.V.; Guntupalli, J.S.; Connolly, A.C.; Halchenko, Y.O.; Conroy, B.R.; Gobbini, M.I.; Hanke, M.; Ramadge, P.J. A common, high-dimensional model of the representational space in human ventral temporal cortex. Neuron 2011, 72, 404–416. [Google Scholar] [CrossRef] [Green Version]
- Reher, K.; Sohn, P. Partly Cloudy [Motion Picture]; Pixar Animation Studios and Walt Disney Pictures: Burbank, CA, USA, 2009. [Google Scholar]
- Varoquaux, G.; Gramfort, A.; Pedregosa, F.; Michel, V.; Thirion, B. Multi-subject dictionary learning to segment an atlas of brain spontaneous activity. In Proceedings of the Biennial International Conference on Information Processing in Medical Imaging, Irsee, Germany, 3 July 2011; pp. 562–573. [Google Scholar]
- Saxe, R.; Kanwisher, N. People thinking about thinking people: The role of the temporo-parietal junction in “theory of mind”. Neuroimage 2003, 19, 1835–1842. [Google Scholar] [CrossRef]
- Gallagher, H.L.; Frith, C.D. Functional imaging of ’theory of mind’. Trends Cogn. Sci. 2003, 7, 77–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, C.; Glover, G.H.; Temple, E. Children’s and adult’s neural bases of verbal and nonverbal theory of mind. Neuropsychologia 2007, 45, 1522–1532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallagher, H.L.; Happé, F.; Brunswick, N.; Fletcher, P.C.; Frith, U.; Frith, C.D. Reading the mind in cartoons and stories: An fMRI study of ‘theory of mind’in verbal and nonverbal tasks. Neuropsychologia 2000, 38, 11–21. [Google Scholar] [CrossRef]
- Gweon, H.; Dodell-Feder, D.; Bedny, M.; Saxe, R. Theory of mind performance in children correlates with functional specialization of a brain region for thinking about thoughts. Child Dev. 2012, 83, 1853–1868. [Google Scholar] [CrossRef]
- Kwon, O.; Sim, J.M. Effects of data set features on the performances of classification algorithms. Expert Syst. Appl. 2013, 40, 1847–1857. [Google Scholar] [CrossRef]
- Rubinov, M.; Sporns, O. Complex network measures of brain connectivity: Uses and interpretations. Neuroimage 2010, 52, 1059–1069. [Google Scholar] [CrossRef]
- Yousefian, A.; Shayegh, F.; Maleki, Z. Detection of autism spectrum disorder using graph representation learning algorithms and deep neural network, based on fMRI signals. Front. Syst. Neurosci. 2022, 16, 904770. [Google Scholar] [CrossRef]
- Suo, X.; Lei, D.; Li, K.; Chen, F.; Li, F.; Li, L.; Huang, X.; Lui, S.; Li, L.; Kemp, G.J.; et al. Disrupted brain network topology in pediatric posttraumatic stress disorder: A resting-state fMRI study. Hum. Brain Mapp. 2015, 36, 3677–3686. [Google Scholar] [CrossRef]
- Lurie, D.J.; Kessler, D.; Bassett, D.S.; Betzel, R.F.; Breakspear, M.; Kheilholz, S.; Kucyi, A.; Liégeois, R.; Lindquist, M.A.; McIntosh, A.R.; et al. Questions and controversies in the study of time-varying functional connectivity in resting fMRI. Netw. Neurosci. 2020, 4, 30–69. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| sFCs | Smoothness Harmonics | |
|---|---|---|
| Inter-subject similarity | Overall: | Overall: |
| Between-adults: | Between-adults: | |
| Between-children: | Between-children: | |
| Between-group classification accuracy (%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bai, W. Smoothness Harmonic: A Graph-Based Approach to Reveal Spatiotemporal Patterns of Cortical Dynamics in fMRI Data. Appl. Sci. 2023, 13, 7130. https://doi.org/10.3390/app13127130
Bai W. Smoothness Harmonic: A Graph-Based Approach to Reveal Spatiotemporal Patterns of Cortical Dynamics in fMRI Data. Applied Sciences. 2023; 13(12):7130. https://doi.org/10.3390/app13127130
Chicago/Turabian StyleBai, Wenjun. 2023. "Smoothness Harmonic: A Graph-Based Approach to Reveal Spatiotemporal Patterns of Cortical Dynamics in fMRI Data" Applied Sciences 13, no. 12: 7130. https://doi.org/10.3390/app13127130