
Interactions among Relevant Non-Saccharomyces, Saccharomyces, and Lactic Acid Bacteria Species of the Wine Microbial Consortium: Towards Advances in Antagonistic Phenomena and Biocontrol Potential
 , ,
, ,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
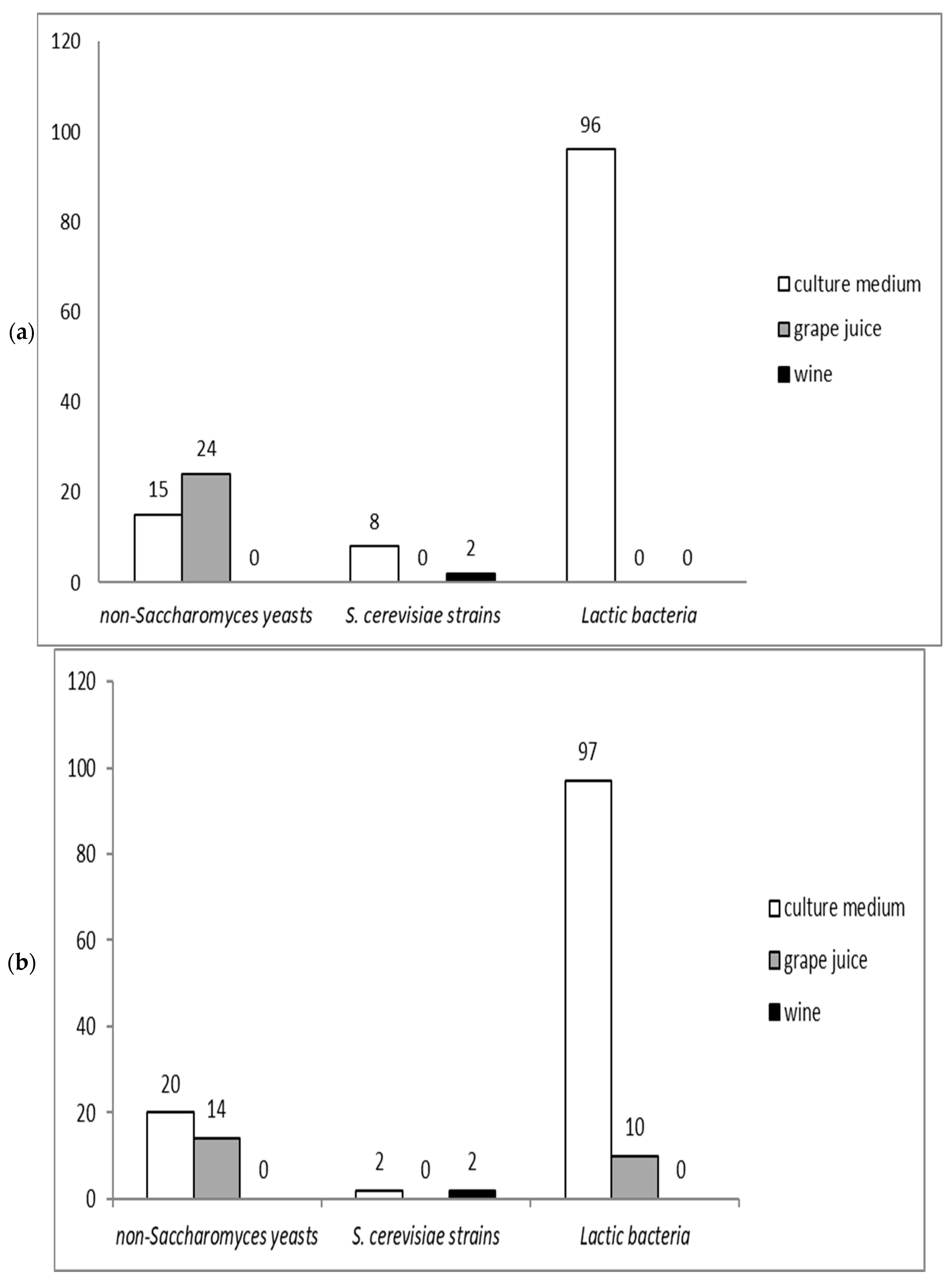
2.1. Microorganisms
2.2. Culture Medium and Growth Conditions
2.3. Interaction Plate Screening: Double-Layer Agar Diffusion Assay
2.4. Interaction Plate Screening: Agar Diffusion Assay
2.5. Interaction Plate Screening: Results Interpretation
3. Results and Discussion
3.1. Non-Saccharomyces as Inhibiting Species
3.2. Saccharomyces as Inhibiting Species
3.3. LAB as Inhibiting Species
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Liu, Y.; Rousseaux, S.; Tourdot-Maréchal, R.; Sadoudi, M.; Gougeon, R.; Schmitt-Kopplin, P.; Alexandre, H. Wine Microbiome: A Dynamic World of Microbial Interactions. Crit. Rev. Food Sci. Nutr. 2017, 57, 856–873. [Google Scholar] [CrossRef] [PubMed]
- Camilo, S.; Chandra, M.; Branco, P.; Malfeito-Ferreira, M. Wine Microbial Consortium: Seasonal Sources and Vectors Linking Vineyard and Winery Environments. Fermentation 2022, 8, 324. [Google Scholar] [CrossRef]
- Berbegal, C.; Spano, G.; Tristezza, M.; Grieco, F.; Capozzi, V. Microbial Resources and Innovation in the Wine Production Sector. South Afr. J. Enol. Vitic. 2017, 38, 156–166. [Google Scholar] [CrossRef]
- Belda, I.; Ruiz, J.; Esteban-Fernández, A.; Navascués, E.; Marquina, D.; Santos, A.; Moreno-Arribas, M.V. Microbial Contribution to Wine Aroma and Its Intended Use for Wine Quality Improvement. Molecules 2017, 22, 189. [Google Scholar] [CrossRef] [Green Version]
- Russo, P.; Capozzi, V.; Spano, G.; Corbo, M.R.; Sinigaglia, M.; Bevilacqua, A. Metabolites of Microbial Origin with an Impact on Health: Ochratoxin A and Biogenic Amines. Front. Microbiol. 2016, 7, 482. [Google Scholar] [CrossRef]
- Roudil, L.; Russo, P.; Berbegal, C.; Albertin, W.; Spano, G.; Capozzi, V. Non-Saccharomyces Commercial Starter Cultures: Scientific Trends, Recent Patents and Innovation in the Wine Sector. FNA 2019, 10, 27–39. [Google Scholar] [CrossRef]
- Petruzzi, L.; Capozzi, V.; Berbegal, C.; Corbo, M.R.; Bevilacqua, A.; Spano, G.; Sinigaglia, M. Microbial Resources and Enological Significance: Opportunities and Benefits. Front. Microbiol. 2017, 8, 995. [Google Scholar] [CrossRef] [Green Version]
- Bindon, K.A.; Kassara, S.; Solomon, M.; Bartel, C.; Smith, P.A.; Barker, A.; Curtin, C. Commercial Saccharomyces cerevisiae Yeast Strains Significantly Impact Shiraz Tannin and Polysaccharide Composition with Implications for Wine Colour and Astringency. Biomolecules 2019, 9, 466. [Google Scholar] [CrossRef] [Green Version]
- Tufariello, M.; Maiorano, G.; Rampino, P.; Spano, G.; Grieco, F.; Perrotta, C.; Capozzi, V.; Grieco, F. Selection of an Autochthonous Yeast Starter Culture for Industrial Production of Primitivo “Gioia Del Colle” PDO/DOC in Apulia (Southern Italy). LWT 2019, 99, 188–196. [Google Scholar] [CrossRef]
- Morata, A.; Loira, I.; Escott, C.; del Fresno, J.M.; Bañuelos, M.A.; Suárez-Lepe, J.A. Applications of Metschnikowia Pulcherrima in Wine Biotechnology. Fermentation 2019, 5, 63. [Google Scholar] [CrossRef]
- Morata, A.; Escott, C.; Bañuelos, M.A.; Loira, I.; del Fresno, J.M.; González, C.; Suárez-Lepe, J.A. Contribution of Non-Saccharomyces Yeasts to Wine Freshness. A Review. Biomolecules 2020, 10, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russo, P.; Tufariello, M.; Renna, R.; Tristezza, M.; Taurino, M.; Palombi, L.; Capozzi, V.; Rizzello, C.G.; Grieco, F. New Insights into the Oenological Significance of Candida zemplinina: Impact of Selected Autochthonous Strains on the Volatile Profile of Apulian Wines. Microorganisms 2020, 8, 628. [Google Scholar] [CrossRef] [PubMed]
- Tufariello, M.; Capozzi, V.; Spano, G.; Cantele, G.; Venerito, P.; Mita, G.; Grieco, F. Effect of Co-Inoculation of Candida Zemplinina, Saccharomyces Cerevisiae and Lactobacillus plantarum for the Industrial Production of Negroamaro Wine in Apulia (Southern Italy). Microorganisms 2020, 8, 726. [Google Scholar] [CrossRef] [PubMed]
- Du Plessis, H.; Du Toit, M.; Nieuwoudt, H.; Van der Rijst, M.; Hoff, J.; Jolly, N. Modulation of Wine Flavor Using Hanseniaspora Uvarum in Combination with Different Saccharomyces cerevisiae, Lactic Acid Bacteria Strains and Malolactic Fermentation Strategies. Fermentation 2019, 5, 64. [Google Scholar] [CrossRef] [Green Version]
- Escribano, R.; González-Arenzana, L.; Garijo, P.; Berlanas, C.; López-Alfaro, I.; López, R.; Gutiérrez, A.R.; Santamaría, P. Screening of Enzymatic Activities within Different Enological Non-Saccharomyces Yeasts. J. Food Sci. Technol. 2017, 54, 1555–1564. [Google Scholar] [CrossRef] [Green Version]
- Berbegal, C.; Khomenko, I.; Russo, P.; Spano, G.; Fragasso, M.; Biasioli, F.; Capozzi, V. PTR-ToF-MS for the Online Monitoring of Alcoholic Fermentation in Wine: Assessment of VOCs Variability Associated with Different Combinations of Saccharomyces/Non-Saccharomyces as a Case-Study. Fermentation 2020, 6, 55. [Google Scholar] [CrossRef]
- Bartowsky, E.J. Oenococcus oeni and Malolactic Fermentation—Moving into the Molecular Arena. Aust. J. Grape Wine Res. 2005, 11, 174–187. [Google Scholar] [CrossRef]
- Campbell-Sills, H.; Khoury, M.E.; Gammacurta, M.; Miot-Sertier, C.; Dutilh, L.; Vestner, J.; Capozzi, V.; Sherman, D.; Hubert, C.; Claisse, O.; et al. Two Different Oenococcus oeni Lineages Are Associated to Either Red or White Wines in Burgundy: Genomics and Metabolomics Insights. OENO One 2017, 51, 309. [Google Scholar] [CrossRef]
- Lasik, M. The Application of Malolactic Fermentation Process to Create Good-Quality Grape Wine Produced in Cool-Climate Countries: A Review. Eur. Food Res. Technol. 2013, 237, 843–850. [Google Scholar] [CrossRef] [Green Version]
- Campbell-Sills, H.; Capozzi, V.; Romano, A.; Cappellin, L.; Spano, G.; Breniaux, M.; Lucas, P.; Biasioli, F. Advances in Wine Analysis by PTR-ToF-MS: Optimization of the Method and Discrimination of Wines from Different Geographical Origins and Fermented with Different Malolactic Starters. Int. J. Mass Spectrom. 2016, 397–398, 42–51. [Google Scholar] [CrossRef]
- Bordet, F.; Joran, A.; Klein, G.; Roullier-Gall, C.; Alexandre, H. Yeast–Yeast Interactions: Mechanisms, Methodologies and Impact on Composition. Microorganisms 2020, 8, 600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alonso-del-Real, J.; Lairón-Peris, M.; Barrio, E.; Querol, A. Effect of Temperature on the Prevalence of Saccharomyces Non Cerevisiae Species against a S. Cerevisiae Wine Strain in Wine Fermentation: Competition, Physiological Fitness, and Influence in Final Wine Composition. Front. Microbiol. 2017, 8, 150. [Google Scholar] [CrossRef] [Green Version]
- Alonso-del-Real, J.; Pérez-Torrado, R.; Querol, A.; Barrio, E. Dominance of Wine Saccharomyces Cerevisiae Strains over S. Kudriavzevii in Industrial Fermentation Competitions Is Related to an Acceleration of Nutrient Uptake and Utilization. Environ. Microbiol. 2019, 21, 1627–1644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bagheri, B.; Bauer, F.F.; Setati, M.E. The Impact of Saccharomyces Cerevisiae on a Wine Yeast Consortium in Natural and Inoculated Fermentations. Front. Microbiol. 2017, 8, 1988. [Google Scholar] [CrossRef]
- Branco, P.; Francisco, D.; Chambon, C.; Hébraud, M.; Arneborg, N.; Almeida, M.G.; Caldeira, J.; Albergaria, H. Identification of Novel GAPDH-Derived Antimicrobial Peptides Secreted by Saccharomyces Cerevisiae and Involved in Wine Microbial Interactions. Appl. Microbiol. Biotechnol. 2014, 98, 843–853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Comitini, F.; De Ingeniis, J.; Pepe, L.; Mannazzu, I.; Ciani, M. Pichia Anomala and Kluyveromyces Wickerhamii Killer Toxins as New Tools against Dekkera/Brettanomyces Spoilage Yeasts. FEMS Microbiol. Lett. 2004, 238, 235–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oro, L.; Ciani, M.; Comitini, F. Antimicrobial Activity of Metschnikowia pulcherrima on Wine Yeasts. J. Appl. Microbiol. 2014, 116, 1209–1217. [Google Scholar] [CrossRef]
- Balmaseda, A.; Bordons, A.; Reguant, C.; Bautista-Gallego, J. Non-Saccharomyces in Wine: Effect Upon Oenococcus oeni and Malolactic Fermentation. Front. Microbiol. 2018, 9, 534. [Google Scholar] [CrossRef] [Green Version]
- Ferrando, N.; Araque, I.; Ortís, A.; Thornes, G.; Bautista-Gallego, J.; Bordons, A.; Reguant, C. Evaluating the Effect of Using Non-Saccharomyces on Oenococcus Oeni and Wine Malolactic Fermentation. Food Res. Int. 2020, 138, 109779. [Google Scholar] [CrossRef]
- Berbegal, C.; Borruso, L.; Fragasso, M.; Tufariello, M.; Russo, P.; Brusetti, L.; Spano, G.; Capozzi, V. A Metagenomic-Based Approach for the Characterization of Bacterial Diversity Associated with Spontaneous Malolactic Fermentations in Wine. Int. J. Mol. Sci. 2019, 20, 3980. [Google Scholar] [CrossRef]
- Englezos, V.; Torchio, F.; Vagnoli, P.; Krieger-Weber, S.; Rantsiou, K.; Cocolin, L. Impact of Saccharomyces Cerevisiae Strain Selection on Malolactic Fermentation by Lactobacillus Plantarum and Oenococcus Oeni. Am. J. Enol. Vitic. 2020, 71, 157–165. [Google Scholar] [CrossRef]
- Berbegal, C.; Garofalo, C.; Russo, P.; Pati, S.; Capozzi, V.; Spano, G. Use of Autochthonous Yeasts and Bacteria in Order to Control Brettanomyces bruxellensis in Wine. Fermentation 2017, 3, 65. [Google Scholar] [CrossRef] [Green Version]
- Berbegal, C.; Spano, G.; Fragasso, M.; Grieco, F.; Russo, P.; Capozzi, V. Starter Cultures as Biocontrol Strategy to Prevent Brettanomyces bruxellensis Proliferation in Wine. Appl. Microbiol. Biotechnol. 2018, 102, 569–576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.-C.; Edwards, C.G.; Peterson, J.C.; Haag, K.M. Relationship Between Sluggish Fermentations and the Antagonism of Yeast by Lactic Acid Bacteria. Am. J. Enol. Vitic. 1996, 47, 1–10. [Google Scholar]
- Yurdugül, S.; Bozoglu, F. Studies on an Inhibitor Produced by Lactic Acid Bacteria of Wines on the Control of Malolactic Fermentation. Eur. Food Res. Technol. 2002, 215, 38–41. [Google Scholar] [CrossRef]
- Comitini, F.; Ciani, M. The Inhibitory Activity of Wine Yeast Starters on Malolactic Bacteria. Ann. Microbiol. 2007, 57, 61. [Google Scholar] [CrossRef]
- Sipiczki, M. Overwintering of Vineyard Yeasts: Survival of Interacting Yeast Communities in Grapes Mummified on Vines. Front. Microbiol. 2016, 7, 212. [Google Scholar] [CrossRef] [Green Version]
- Mehlomakulu, N.N.; Setati, M.E.; Divol, B. Characterization of Novel Killer Toxins Secreted by Wine-Related Non-Saccharomyces Yeasts and Their Action on Brettanomyces Spp. Int. J. Food Microbiol. 2014, 188, 83–91. [Google Scholar] [CrossRef]
- Pinto, L.; Malfeito-Ferreira, M.; Quintieri, L.; Silva, A.C.; Baruzzi, F. Growth and Metabolite Production of a Grape Sour Rot Yeast-Bacterium Consortium on Different Carbon Sources. Int. J. Food Microbiol. 2019, 296, 65–74. [Google Scholar] [CrossRef]
- Kemsawasd, V.; Branco, P.; Almeida, M.G.; Caldeira, J.; Albergaria, H.; Arneborg, N. Cell-to-Cell Contact and Antimicrobial Peptides Play a Combined Role in the Death of Lachanchea Thermotolerans during Mixed-Culture Alcoholic Fermentation with Saccharomyces cerevisiae. FEMS Microbiol. Lett. 2015, 362, fnv103. [Google Scholar] [CrossRef]
- Kapetanakou, A.E.; Kollias, J.N.; Drosinos, E.H.; Skandamis, P.N. Inhibition of A. Carbonarius Growth and Reduction of Ochratoxin A by Bacteria and Yeast Composites of Technological Importance in Culture Media and Beverages. Int. J. Food Microbiol. 2012, 152, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Garofalo, C.; Khoury, M.E.; Lucas, P.; Bely, M.; Russo, P.; Spano, G.; Capozzi, V. Autochthonous Starter Cultures and Indigenous Grape Variety for Regional Wine Production. J. Appl. Microbiol. 2015, 118, 1395–1408. [Google Scholar] [CrossRef] [PubMed]
- Berbegal, C.; Peña, N.; Russo, P.; Grieco, F.; Pardo, I.; Ferrer, S.; Spano, G.; Capozzi, V. Technological Properties of Lactobacillus Plantarum Strains Isolated from Grape Must Fermentation. Food Microbiol. 2016, 57, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Englezos, V.; Jolly, N.P.; Di Gianvito, P.; Rantsiou, K.; Cocolin, L. Microbial Interactions in Winemaking: Ecological Aspects and Effect on Wine Quality. Trends Food Sci. Technol. 2022, 127, 99–113. [Google Scholar] [CrossRef]
- Di Toro, M.R.; Capozzi, V.; Beneduce, L.; Alexandre, H.; Tristezza, M.; Durante, M.; Tufariello, M.; Grieco, F.; Spano, G. Intraspecific Biodiversity and ‘Spoilage Potential’ of Brettanomyces bruxellensis in Apulian Wines. LWT Food Sci. Technol. 2015, 60, 102–108. [Google Scholar] [CrossRef]
- Lamontanara, A.; Caggianiello, G.; Orrù, L.; Capozzi, V.; Michelotti, V.; Bayjanov, J.R.; Renckens, B.; van Hijum, S.A.F.T.; Cattivelli, L.; Spano, G. Draft Genome Sequence of Lactobacillus plantarum Lp90 Isolated from Wine. Genome Announc. 2015, 3, e00097-15. [Google Scholar] [CrossRef] [Green Version]
- Lamontanara, A.; Orrù, L.; Cattivelli, L.; Russo, P.; Spano, G.; Capozzi, V. Genome Sequence of Oenococcus oeni OM27, the First Fully Assembled Genome of a Strain Isolated from an Italian Wine. Genome Announc. 2014, 2, e00658-14. [Google Scholar] [CrossRef] [Green Version]
- Capozzi, V.; Russo, P.; Lamontanara, A.; Orrù, L.; Cattivelli, L.; Spano, G. Genome Sequences of Five Oenococcus oeni Strains Isolated from Nero Di Troia Wine from the Same Terroir in Apulia, Southern Italy. Genome Announc. 2014, 2, e01077-14. [Google Scholar] [CrossRef] [Green Version]
- Garofalo, C.; Russo, P.; Beneduce, L.; Massa, S.; Spano, G.; Capozzi, V. Non-Saccharomyces Biodiversity in Wine and the ‘Microbial Terroir’: A Survey on Nero Di Troia Wine from the Apulian Region, Italy. Ann. Microbiol. 2016, 66, 143–150. [Google Scholar] [CrossRef]
- Garofalo, C.; Tristezza, M.; Grieco, F.; Spano, G.; Capozzi, V. From Grape Berries to Wine: Population Dynamics of Cultivable Yeasts Associated to “Nero Di Troia” Autochthonous Grape Cultivar. World J. Microbiol. Biotechnol. 2016, 32, 59. [Google Scholar] [CrossRef]
- Garofalo, C.; Berbegal, C.; Grieco, F.; Tufariello, M.; Spano, G.; Capozzi, V. Selection of Indigenous Yeast Strains for the Production of Sparkling Wines from Native Apulian Grape Varieties. Int. J. Food Microbiol. 2018, 285, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Capozzi, V.; Berbegal, C.; Tufariello, M.; Grieco, F.; Spano, G.; Grieco, F. Impact of co-inoculation of Saccharomyces cerevisiae, Hanseniaspora uvarum and Oenococcus oeni autochthonous strains in controlled multi starter grape must fermentations. LWT 2019, 109, 241–249. [Google Scholar] [CrossRef]
- Comitini, F.; Ferretti, R.; Clementi, F.; Mannazzu, I.; Ciani, M. Interactions between Saccharomyces cerevisiae and Malolactic Bacteria: Preliminary Characterization of a Yeast Proteinaceous Compound (s) Active against Oenococcus oeni. J. Appl. Microbiol. 2005, 99, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Ivey, M.; Massel, M.; Phister, T.G. Microbial Interactions in Food Fermentations. Annu. Rev. Food Sci. Technol. 2013, 4, 141–162. [Google Scholar] [CrossRef]
- Ponomarova, O.; Gabrielli, N.; Sévin, D.C.; Mülleder, M.; Zirngibl, K.; Bulyha, K.; Andrejev, S.; Kafkia, E.; Typas, A.; Sauer, U.; et al. Yeast Creates a Niche for Symbiotic Lactic Acid Bacteria through Nitrogen Overflow. Cell Syst. 2017, 5, 345–357.e6. [Google Scholar] [CrossRef] [Green Version]
- Mendoza, L.M.; de Nadra, M.C.M.; Farías, M.E. Kinetics and Metabolic Behavior of a Composite Culture of Kloeckera Apiculata and Saccharomyces cerevisiae Wine Related Strains. Biotechnol. Lett. 2007, 29, 1057–1063. [Google Scholar] [CrossRef]
- Köhl, J.; Kolnaar, R.; Ravensberg, W.J. Mode of Action of Microbial Biological Control Agents Against Plant Diseases: Relevance Beyond Efficacy. Front. Plant Sci. 2019, 10, 845. [Google Scholar] [CrossRef] [Green Version]
- Vejarano, R. Non-Saccharomyces in Winemaking: Source of Mannoproteins, Nitrogen, Enzymes, and Antimicrobial Compounds. Fermentation 2020, 6, 76. [Google Scholar] [CrossRef]
- Berbegal, C.; Fragasso, M.; Russo, P.; Bimbo, F.; Grieco, F.; Spano, G.; Capozzi, V. Climate Changes and Food Quality: The Potential of Microbial Activities as Mitigating Strategies in the Wine Sector. Fermentation 2019, 5, 85. [Google Scholar] [CrossRef] [Green Version]
- Jolly, N.P.; Varela, C.; Pretorius, I.S. Not Your Ordinary Yeast: Non-Saccharomyces Yeasts in Wine Production Uncovered. FEMS Yeast Res. 2014, 14, 215–237. [Google Scholar] [CrossRef] [Green Version]
- Türkel, S.; Ener, B. Isolation and Characterization of New Metschnikowia Pulcherrima Strains as Producers of the Antimicrobial Pigment Pulcherrimin. Z. Naturforsch. C J. Biosci. 2009, 64, 405–410. [Google Scholar] [CrossRef] [PubMed]
- Medina, K.; Boido, E.; Dellacassa, E.; Carrau, F. Growth of Non-Saccharomyces Yeasts Affects Nutrient Availability for Saccharomyces Cerevisiae during Wine Fermentation. Int. J. Food Microbiol. 2012, 157, 245–250. [Google Scholar] [CrossRef] [PubMed]
- Mendoza, L.M.; de Nadra, M.C.M.; Farías, M.E. Antagonistic Interaction between Yeasts and Lactic Acid Bacteria of Oenological Relevance: Partial Characterization of Inhibitory Compounds Produced by Yeasts. Food Res. Int. 2010, 43, 1990–1998. [Google Scholar] [CrossRef] [Green Version]
- Melvydas, V.; Svediene, J.; Skridlaite, G.; Vaiciuniene, J.; Garjonyte, R. In Vitro Inhibition of Saccharomyces cerevisiae Growth by Metschnikowia Spp. Triggered by Fast Removal of Iron via Two Ways. Braz. J. Microbiol. 2020, 51, 1953–1964. [Google Scholar] [CrossRef]
- Nardi, T.; Panero, L.; Petrozziello, M.; Guaita, M.; Tsolakis, C.; Cassino, C.; Vagnoli, P.; Bosso, A. Managing Wine Quality Using Torulaspora Delbrueckii and Oenococcus Oeni Starters in Mixed Fermentations of a Red Barbera Wine. Eur. Food Res. Technol. 2019, 245, 293–307. [Google Scholar] [CrossRef]
- Du Plessis, H.W.; du Toit, M.; Hoff, J.W.; Hart, R.S.; Ndimba, B.K.; Jolly, N.P. Characterisation of Non-Saccharomyces Yeasts Using Different Methodologies and Evaluation of Their Compatibility with Malolactic Fermentation. South Afr. J. Enol. Vitic. 2017, 38, 46–63. [Google Scholar] [CrossRef]
- Russo, P.; Englezos, V.; Capozzi, V.; Pollon, M.; Río Segade, S.; Rantsiou, K.; Spano, G.; Cocolin, L. Effect of Mixed Fermentations with Starmerella Bacillaris and Saccharomyces cerevisiae on Management of Malolactic Fermentation. Food Res. Int. 2020, 134, 109246. [Google Scholar] [CrossRef]
- Nardi, T. Microbial Resources as a Tool for Enhancing Sustainability in Winemaking. Microorganisms 2020, 8, 507. [Google Scholar] [CrossRef] [Green Version]
- Bartle, L.; Sumby, K.; Sundstrom, J.; Jiranek, V. The Microbial Challenge of Winemaking: Yeast-Bacteria Compatibility. FEMS Yeast Res. 2019, 19, foz040. [Google Scholar] [CrossRef]
- Capozzi, V.; Garofalo, C.; Chiriatti, M.A.; Grieco, F.; Spano, G. Microbial Terroir and Food Innovation: The Case of Yeast Biodiversity in Wine. Microbiol. Res. 2015, 181, 75–83. [Google Scholar] [CrossRef]
- Belda, I.; Ruiz, J.; Alonso, A.; Marquina, D.; Santos, A. The Biology of Pichia Membranifaciens Killer Toxins. Toxins 2017, 9, 112. [Google Scholar] [CrossRef] [PubMed]
- De Ullivarri, M.F.; Mendoza, L.M.; Raya, R.R. Killer Yeasts as Biocontrol Agents of Spoilage Yeasts and Bacteria Isolated from Wine. BIO Web Conf. 2014, 3, 02001. [Google Scholar] [CrossRef]
- Russo, P.; Fragasso, M.; Berbegal, C.; Grieco, F.; Spano, G.; Capozzi, V. Microorganisms Able to Produce Biogenic Amines and Factors Affecting Their Activity. In Biogenic Amines in Food; Royal Society of Chemistry Publishing: Cambridge, UK, 2019; pp. 18–40. [Google Scholar]
- Barbieri, F.; Montanari, C.; Gardini, F.; Tabanelli, G. Biogenic Amine Production by Lactic Acid Bacteria: A Review. Foods 2019, 8, 17. [Google Scholar] [CrossRef] [Green Version]
- Krieger-Weber, S.; Heras, J.M.; Suarez, C. Lactobacillus plantarum, a New Biological Tool to Control Malolactic Fermentation: A Review and an Outlook. Beverages 2020, 6, 23. [Google Scholar] [CrossRef] [Green Version]
- Alexandre, H.; Costello, P.J.; Remize, F.; Guzzo, J.; Guilloux-Benatier, M. Saccharomyces cerevisiae–Oenococcus oeni Interactions in Wine: Current Knowledge and Perspectives. Int. J. Food Microbiol. 2004, 93, 141–154. [Google Scholar] [CrossRef]
- Aponte, M.; Romano, R.; Villano, C.; Blaiotta, G. Dominance of S. cerevisiae Commercial Starter Strains during Greco Di Tufo and Aglianico Wine Fermentations and Evaluation of Oenological Performances of Some Indigenous/Residential Strains. Foods 2020, 9, 1549. [Google Scholar] [CrossRef] [PubMed]
- Perrone, B.; Giacosa, S.; Rolle, L.; Cocolin, L.; Rantsiou, K. Investigation of the Dominance Behavior of Saccharomyces cerevisiae Strains during Wine Fermentation. Int. J. Food Microbiol. 2013, 165, 156–162. [Google Scholar] [CrossRef]
- Barrajón, N.; Arévalo-Villena, M.; Úbeda, J.; Briones, A. Enological Properties in Wild and Commercial Saccharomyces cerevisiae Yeasts: Relationship with Competition during Alcoholic Fermentation. World J. Microbiol. Biotechnol. 2011, 27, 2703–2710. [Google Scholar] [CrossRef]
- Papadimitriou, K.; Alegría, Á.; Bron, P.A.; de Angelis, M.; Gobbetti, M.; Kleerebezem, M.; Lemos, J.A.; Linares, D.M.; Ross, P.; Stanton, C.; et al. Stress Physiology of Lactic Acid Bacteria. Microbiol. Mol. Biol. Rev. 2016, 80, 837–890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leroy, F.; De Vuyst, L. Growth of the Bacteriocin-Producing Lactobacillus sakei Strain CTC 494 in MRS Broth Is Strongly Reduced Due to Nutrient Exhaustion: A Nutrient Depletion Model for the Growth of Lactic Acid Bacteria. Appl. Environ. Microbiol. 2001, 67, 4407–4413. [Google Scholar] [CrossRef] [Green Version]
- Arena, M.P.; Capozzi, V.; Spano, G.; Fiocco, D. The Potential of Lactic Acid Bacteria to Colonize Biotic and Abiotic Surfaces and the Investigation of Their Interactions and Mechanisms. Appl. Microbiol. Biotechnol. 2017, 101, 2641–2657. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, A.V.; de Nadra, M.C.M. Production of Hydrogen Peroxide by Lactobacillus hilgardii and Its Effect on Leuconostoc oenos Growth. Curr. Microbiol. 1995, 30, 23–25. [Google Scholar] [CrossRef]
- Aredes Fernández, P.A.; Farías, M.E.; de Nadra, M.C.M. Interaction between Oenococcus oeni and Lactobacillus hilgardii Isolated from Wine. Modification of Available Nitrogen and Biogenic Amine Production. Biotechnol. Lett. 2010, 32, 1095–1102. [Google Scholar] [CrossRef] [PubMed]
- Castellano, P.; Pérez Ibarreche, M.; Blanco Massani, M.; Fontana, C.; Vignolo, G.M. Strategies for Pathogen Biocontrol Using Lactic Acid Bacteria and Their Metabolites: A Focus on Meat Ecosystems and Industrial Environments. Microorganisms 2017, 5, 38. [Google Scholar] [CrossRef] [Green Version]
- Kuchen, B.; Vazquez, F.; Maturano, Y.P.; Scaglia, G.J.E.; Pera, L.M.; Vallejo, M.D. Toward Application of Biocontrol to Inhibit Wine Spoilage Yeasts: The Use of Statistical Designs for Screening and Optimisation. OENO One 2021, 55, 75–96. [Google Scholar] [CrossRef]
- Petitgonnet, C.; Klein, G.L.; Roullier-Gall, C.; Schmitt-Kopplin, P.; Quintanilla-Casas, B.; Vichi, S.; Julien-David, D.; Alexandre, H. Influence of Cell-Cell Contact between L. thermotolerans and S. cerevisiae on Yeast Interactions and the Exo-Metabolome. Food Microbiol. 2019, 83, 122–133. [Google Scholar] [CrossRef]
- Roullier-Gall, C.; David, V.; Hemmler, D.; Schmitt-Kopplin, P.; Alexandre, H. Exploring Yeast Interactions through Metabolic Profiling. Sci. Rep. 2020, 10, 6073. [Google Scholar] [CrossRef] [Green Version]
- Ciani, M.; Comitini, F. Yeast Interactions in Multi-Starter Wine Fermentation. Curr. Opin. Food Sci. 2015, 1, 1–6. [Google Scholar] [CrossRef]
- Marzano, M.; Fosso, B.; Manzari, C.; Grieco, F.; Intranuovo, M.; Cozzi, G.; Mulè, G.; Scioscia, G.; Valiente, G.; Tullo, A.; et al. Complexity and dynamics of the winemaking bacterial communities in berries, musts, and wines from Apulian grape cultivars through time and space. PLoS ONE 2016, 11, e0157383. [Google Scholar] [CrossRef]
- Morata, A.; Arroyo, T.; Bañuelos, M.A.; Blanco, P.; Briones, A.; Cantoral, J.M.; Castrillo, D.; Cordero-Bueso, G.; Del Fresno, J.M.; Escott, C. Wine Yeast Selection in the Iberian Peninsula: Saccharomyces and Non-Saccharomyces as Drivers of Innovation in Spanish and Portuguese Wine Industries. Crit. Rev. Food Sci. Nutr. 2022, 1–29. [Google Scholar] [CrossRef]


{kind=link}
| Species | Strain Code | Matrix/Source |
|---|---|---|
| Saccharomyces cerevisiae | superlievito alcoligens | CSC |
| S. cerevisiae | elegance | CSC |
| S. cerevisiae ex bayanus | EC1118 | CSC |
| S. cerevisiae | E4 | OSF |
| S. cerevisiae | I6 | OSF |
| S. cerevisiae | SUPRARED HG | CSC |
| S. cerevisiae | T2 | CSC |
| Hanseniaspora uvarum | 1444 | CECT |
| H. uvarum | B05B29 | OSF |
| Hanseniaspora guilliermondii | M105A31 | OSF |
| Torulaspora delbrueckii | 11199 | CECT |
| T. delbrueckii | B05B12 | OSF |
| T. delbrueckii | 291 | CSC |
| Pichia fermentans | M105A3 | OSF |
| P. fermentans | B05A36 | OSF |
| P. fermentans | B05A29 | OSF |
| Issatchenkia terricola | B05B8 | OSF |
| Starmerella bacillaris | B05B6 | OSF |
| Metschnikowia pulcherrima | B0512B3 | OSF |
| M. pulcherrima | B0512B24 | OSF |
| M. pulcherrima | B0512B25 | OSF |
| M. pulcherrima | B0512B26 | OSF |
| M. pulcherrima | B0512B15 | OSF |
| M. pulcherrima | B05B2P | OSF |
| M. pulcherrima | B05A36 | OSF |
| M. pulcherrima | M105A51 | OSF |
| M. pulcherrima | B0522 | OSF |
| M. pulcherrima | 346 | CSC |
| Brettanomyces bruxellensis | 2 | OSF |
| B. bruxellensis | 4 | OSF |
| B. bruxellensis | 5 | OSF |
| B. bruxellensis | 6 | OSF |
| B. bruxellensis | 7 | OSF |
| Lactiplantibacillus plantarum | Lp90 | OSF |
| L. plantarum | 44 | OSF |
| L. plantarum | V22 | CSC |
| L. plantarum | 38 CDS | OSF |
| L. plantarum | T1 | OSF |
| Levilactobacillus brevis | 9809 | IOEB |
| Lentilactobacillus hilgardii | 4786 | CECT |
| Pediococcus parvulus | 126 | OSF |
| Pediococcus spp. | 32 | OSF |
| Oenococcus oeni | OT3 | OSF |
| O. oeni | 6 | OSF |
| O. oeni | OT4 | OSF |
| Inhibiting Species | Inhibited Species | Inhibitory Activity |
|---|---|---|
| M. pulcherrima 346 | S. cerevisiae I6 | ± C |
| L. plantarum V22 | ± C | |
| L. brevis IOEB | ± C | |
| L. plantarum 38 CDS | ± C | |
| Pediococcus spp. | ± C | |
| O. oeni OT3 | ± C | |
| T. delbrueckii 291 | L. plantarum Lp90 | ++ C |
| L. plantarum 44 | ++ C | |
| L. brevis IOEB | ++ C | |
| P. parvulus 126 | ++ C | |
| L. hilgardii CECT 4786 | ++ C | |
| L. plantarum 38 CDS | ++ C | |
| Pediococcus spp. | ± C | |
| P. fermentans M105A30 | Pediococcus spp. | ± C |
| P. fermentans B05A36 | Pediococcus spp. | ± C |
| H. guilliermondii M105A31 | P. fermentans M105A30 | ± J |
| H. uvarum B05B29 | P. fermentans M105A30 | ± J |
| P. fermentans B05A29 | T. delbrueckii 291 | ± J |
| T. delbrueckii CECT 11199 | ± J | |
| B. bruxellensis 2 | ± J | |
| B. bruxellensis 6 | ± J | |
| M. pulcherrima B0512B3 | P. fermentans M105A30 | ± J |
| M. pulcherrima B0512B24 | P. fermentans M105A30 | ± J |
| M. pulcherrima B0512B25 | P. fermentans M105A30 | ± J |
| M. pulcherrima B0512B26 | P. fermentans M105A30 | ± J |
| M. pulcherrima B0512B15 | P. fermentans M105A30 | ± J |
| M. pulcherrima B05B2P | P. fermentans M105A30 | ± J |
| M. pulcherrima B05A36 | P. fermentans M105A30 | ± J |
| M. pulcherrima B0522 | P. fermentans M105A30 | ± J |
| B. bruxellensis 2 | Pediococcus spp. | ++ J |
| O. oeni 6 | ++ J | |
| B. bruxellensis 4 | Pediococcus spp. | ± J |
| O. oeni 6 | + J | |
| B. bruxellensis 5 | Pediococcus spp. | ± J |
| O. oeni 6 | + J | |
| B. bruxellensis 6 | Pediococcus spp. | ± J |
| O. oeni 6 | + J | |
| B. bruxellensis 7 | Pediococcus spp. | ± J |
| O. oeni 6 | + J |
| Inhibiting Species | Inhibited Species | Inhibitory Activity |
|---|---|---|
| S. cerevisiae T2 | S. cerevisiae superlievito alcoligens | ± W |
| S. cerevisiae I6 | ± C,W | |
| S. cerevisiae superlievito alcoligens | L. plantarum Lp90 | ++ C |
| L. plantarum 44 | ++ C | |
| L. plantarum V22 | ++ C | |
| L. brevis IOEB | ++ C | |
| P. parvulus 126 | ++ C | |
| L. hilgardii CECT 4786 | ++ C | |
| L. plantarum 38 CDS | ++ C |
| Inhibiting Species | Inhibited Species | Inhibitory Activity |
|---|---|---|
| L. plantarum Lp90 | L. plantarum 44 | ± C |
| L. plantarum V22 | ± C | |
| L. brevis IOEB | ± C | |
| P. parvulus 126 | ± C | |
| L. hilgardii CECT 4786 | ± C | |
| L. plantarum 38 CDS | + C | |
| Pediococcus spp. | ± C | |
| L. plantarum 44 | L. plantarum 44 | ± C |
| L. plantarum V22 | ± C | |
| L. brevis IOEB | ± C | |
| P. parvulus 126 | ± C | |
| L. hilgardii CECT 4786 | ± C | |
| L. plantarum 38 CDS | + C | |
| Pediococcus spp. | ± C | |
| L. plantarum V22 | L. plantarum Lp90 | + C |
| L. plantarum 44 | ± C | |
| L. plantarum V22 | ± C | |
| L. brevis IOEB | ± C | |
| P. parvulus 126 | ± C | |
| L. hilgardii CECT 4786 | ± C | |
| L. plantarum 38 CDS | + C | |
| Pediococcus spp. | ± C | |
| L. brevis IOEB | L. plantarum Lp90 | + C |
| L. plantarum 44 | ± C | |
| L. plantarum V22 | ± C | |
| L. brevis IOEB | ± C | |
| P. parvulus 126 | ± C | |
| L. hilgardii CECT 4786 | ± C | |
| L. plantarum 38 CDS | + C | |
| Pediococcus spp. | ± C | |
| P. parvulus 126 | L. plantarum Lp90 | + C |
| L. plantarum 44 | ± C | |
| L. plantarum V22 | ± C | |
| L. brevis IOEB | ± C | |
| P. parvulus 126 | ± C | |
| L. hilgardii CECT 4786 | ± C | |
| L. plantarum 38 CDS | + C | |
| Pediococcus spp. | ± C | |
| L. hilgardii CECT 4786 | L. plantarum 44 | ± C |
| L. plantarum V22 | ± C | |
| L. brevis IOEB | ± C | |
| P. parvulus 126 | ± C | |
| L. hilgardii CECT 4786 | ± C | |
| L. plantarum 38 CDS | + C | |
| Pediococcus spp. | ± C | |
| L. plantarum 38 CDS | L. plantarum 44 | ± C |
| L. plantarum V22 | ± C | |
| L. brevis IOEB | ± C | |
| P. parvulus 126 | ± C | |
| L. hilgardii CECT 4786 | ± C | |
| L. plantarum 38 CDS | + C | |
| Pediococcus spp. | ± C | |
| L. plantarum T1 | L. plantarum Lp90 | ++ C |
| L. plantarum 44 | ++ C | |
| L. brevis IOEB | ++ C | |
| P. parvulus 126 | ++ C | |
| L. plantarum 38 CDS | ++ C | |
| Pediococcus spp. | + C | |
| Pediococcus spp. | O. oeni OT3 | + C |
| L. plantarum Lp90 | ++ C | |
| L. plantarum 44 | + C | |
| L. plantarum V22 | ++ C | |
| L. brevis IOEB | ± C | |
| P. parvulus 126 | + C | |
| L. hilgardii CECT 4786 | ± C | |
| L. plantarum 38 CDS | ++ C | |
| M. pulcherrima B0512B3 | ± C | |
| M. pulcherrima B0512B25 | ± C | |
| P. fermentans M105A30 | ++ C | |
| P. fermentans B05A36 | ++ C | |
| T. delbrueckii 291 | ++ C | |
| O. oeni OT3 | O. oeni OT3 | + C |
| L. plantarum Lp90 | ++ C | |
| L. plantarum 44 | + C | |
| L. plantarum V22 | ++ C | |
| L. brevis IOEB | ± C | |
| P. parvulus 126 | ± C | |
| L. hilgardii CECT 4786 | ± C | |
| L. plantarum 38 CDS | ++ C | |
| M. pulcherrima B0512B3 | ± C | |
| M. pulcherrima B0512B25 | ± C | |
| M. pulcherrima B0512B26 | ± C | |
| M. pulcherrima B0522 | ± C | |
| P. fermentans M105A30 | + C | |
| P. fermentans B05A36 | ++ C | |
| T. delbrueckii 291 | + C | |
| O. oeni 6 | O. oeni OT3 | ± C |
| O. oeni OT4 | O. oeni OT3 | ± C |
| M. pulcherrima B0512B3 | ± C | |
| P. fermentans M105A30 | ++ C | |
| P. fermentans B05A36 | ± C | |
| T. delbrueckii 291 | ++ C | |
| B. bruxellensis 2 | + C | |
| B. bruxellensis 4 | ± C | |
| B. bruxellensis 6 | ± C | |
| B. bruxellensis 7 | ± C |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Gioia, M.; Russo, P.; De Simone, N.; Grieco, F.; Spano, G.; Capozzi, V.; Fragasso, M. Interactions among Relevant Non-Saccharomyces, Saccharomyces, and Lactic Acid Bacteria Species of the Wine Microbial Consortium: Towards Advances in Antagonistic Phenomena and Biocontrol Potential. Appl. Sci. 2022, 12, 12760. https://doi.org/10.3390/app122412760
De Gioia M, Russo P, De Simone N, Grieco F, Spano G, Capozzi V, Fragasso M. Interactions among Relevant Non-Saccharomyces, Saccharomyces, and Lactic Acid Bacteria Species of the Wine Microbial Consortium: Towards Advances in Antagonistic Phenomena and Biocontrol Potential. Applied Sciences. 2022; 12(24):12760. https://doi.org/10.3390/app122412760
Chicago/Turabian StyleDe Gioia, Marianna, Pasquale Russo, Nicola De Simone, Francesco Grieco, Giuseppe Spano, Vittorio Capozzi, and Mariagiovanna Fragasso. 2022. "Interactions among Relevant Non-Saccharomyces, Saccharomyces, and Lactic Acid Bacteria Species of the Wine Microbial Consortium: Towards Advances in Antagonistic Phenomena and Biocontrol Potential" Applied Sciences 12, no. 24: 12760. https://doi.org/10.3390/app122412760