
The Ability to Normalise Energy Metabolism in Advanced COVID-19 Disease Seems to Be One of the Key Factors Determining the Disease Progression—A Metabolomic NMR Study on Blood Plasma

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Sample Preparation
2.3. NMR Measurement
2.4. Data Processing
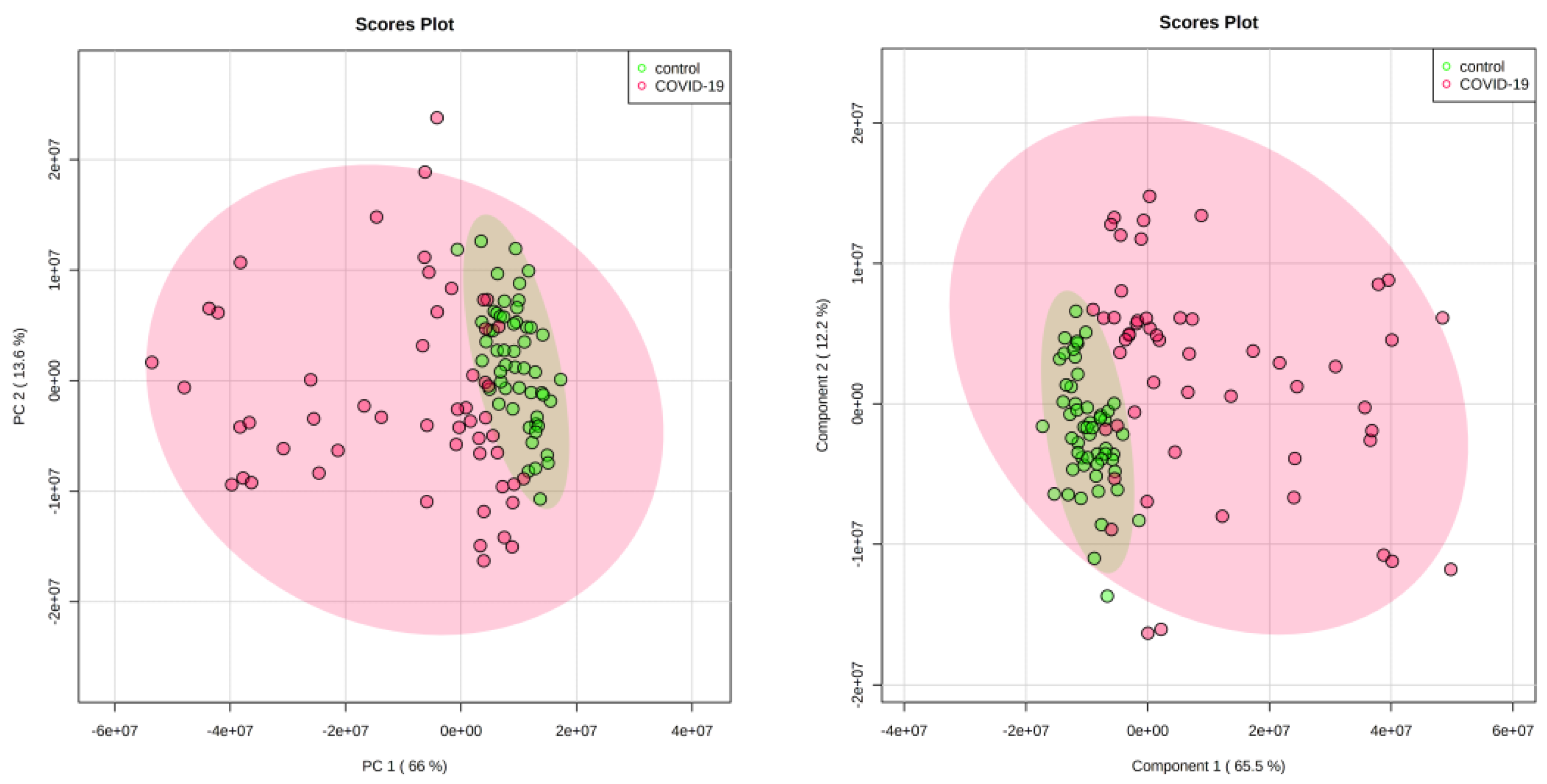
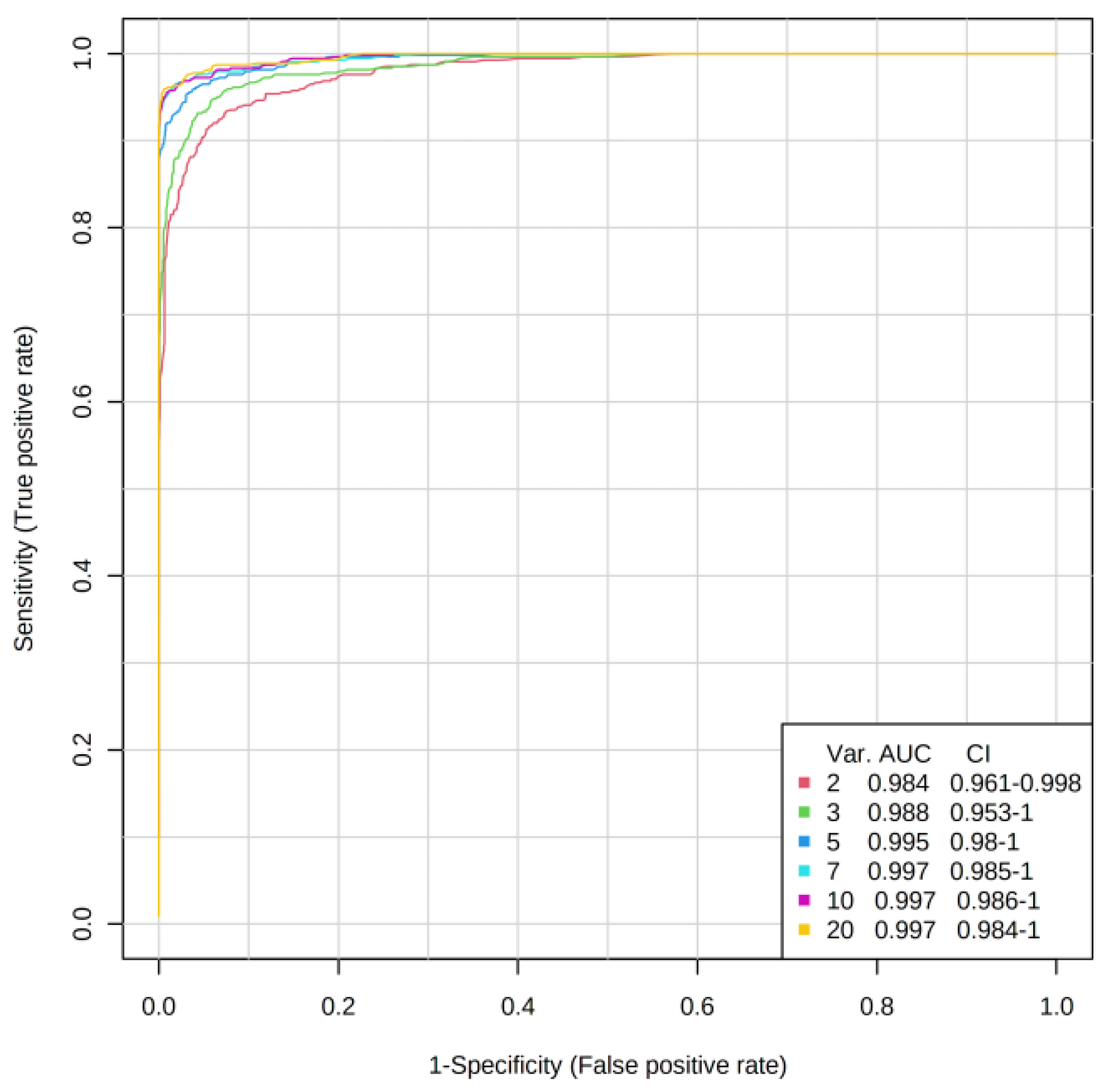
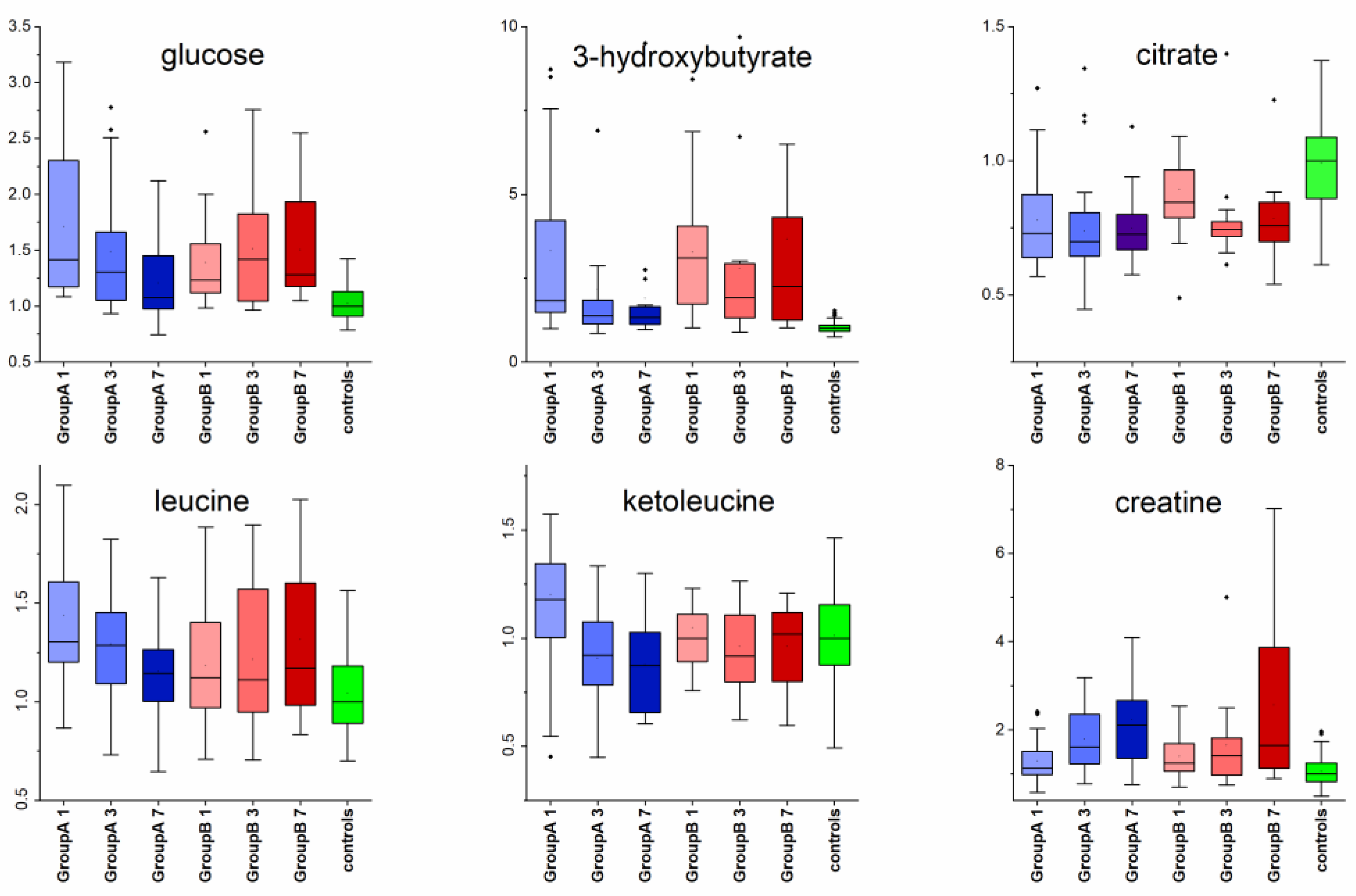
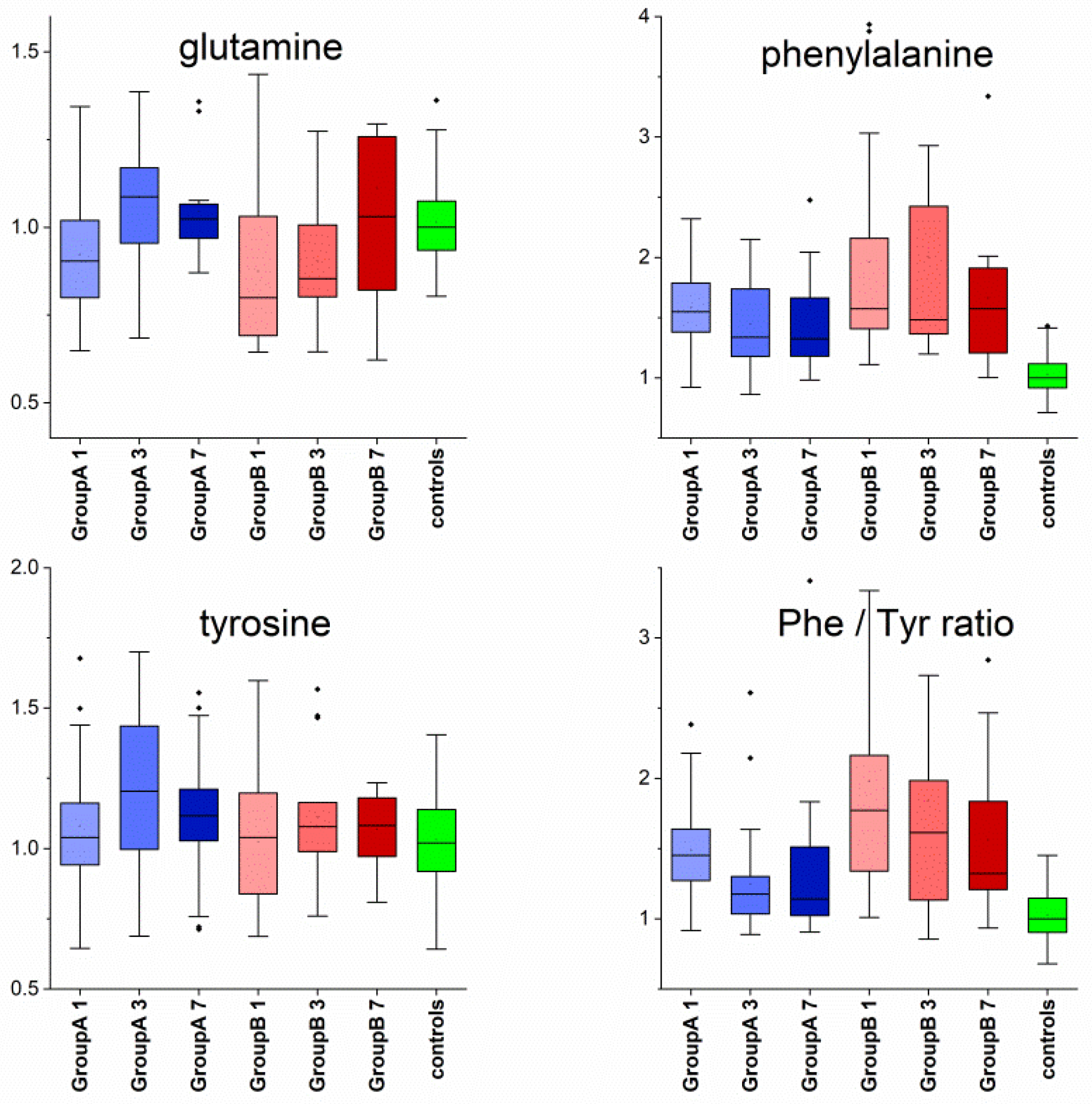
3. Results
4. Discussion
4.1. Discriminatory Analyses
4.2. Metabolomic Changes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wellen, K.E.; Hotamisligil, G.S. Inflammation, stress, and diabetes. J. Clin. Investig. 2005, 115, 1111–1119. [Google Scholar] [CrossRef] [Green Version]
- Jung, J.; Zeng, H.; Horng, T. Metabolism as a guiding force for immunity. Nat. Cell Biol. 2019, 21, 85–93. [Google Scholar] [CrossRef]
- McGaha, T.L.; Huang, L.; Lemos, H.; Metz, R.; Mautino, M.; Prendergast, G.C.; Mellor, A.L. Amino acid catabolism: A pivotal regulator of innate and adaptive immunity. Immunol. Rev. 2012, 249, 135–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jafar, N.; Edriss, H.; Nugent, K. The Effect of Short-Term Hyperglycemia on the Innate Immune System. Am. J. Med Sci. 2016, 351, 201–211. [Google Scholar] [CrossRef]
- Berbudi, A.; Rahmadika, N.; Tjahjadi, A.I.; Ruslami, R. Type 2 Diabetes and its Impact on the Immune System. Curr. Diabetes Rev. 2020, 16, 442–449. [Google Scholar] [CrossRef] [PubMed]
- Geerlings, S.E.; Hoepelman, A.I.M. Immune dysfunction in patients with diabetes mellitus (DM). FEMS Immunol. Med. Microbiol. 1999, 26, 259–265. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Shu, T.; Yang, X.; Song, J.-X.; Zhang, M.; Yao, C.; Liu, W.; Huang, M.; Yu, Y.; Yang, Q.; et al. Plasma metabolomic and lipidomic alterations associated with COVID-19. Natl. Sci. Rev. 2020, 7, 1157–1168. [Google Scholar] [CrossRef]
- Song, J.-W.; Lam, S.M.; Fan, X.; Cao, W.-J.; Wang, S.-Y.; Tian, H.; Chua, G.H.; Zhang, C.; Meng, F.-P.; Xu, Z.; et al. Omics-Driven Systems Interrogation of Metabolic Dysregulation in COVID-19 Pathogenesis. Cell Metab. 2020, 32, 188–202. [Google Scholar] [CrossRef]
- Blasco, H.; Bessy, C.; Plantier, L.; Lefevre, A.; Piver, E.; Bernard, L.; Marlet, J.; Stefic, K.; Bretagne, I.B.-D.; Cannet, P.; et al. The specific metabolome profiling of patients infected by SARS-COV-2 supports the key role of tryptophan-nicotinamide pathway and cytosine metabolism. Sci. Rep. 2020, 10, 1–12. [Google Scholar] [CrossRef]
- Shen, B.; Yi, X.; Sun, Y.; Bi, X.; Du, J. Proteomic and metabolomic characterisation of COVID-19 patient sera. Cell 2020, 182, 59–72. [Google Scholar] [CrossRef] [PubMed]
- Pang, Z.; Zhou, G.; Chong, J.; Xia, J. Comprehensive Meta-Analysis of COVID-19 Global Metabolomics Datasets. Metabolites 2021, 11, 44. [Google Scholar] [CrossRef]
- Roberts, I.; Muelas, M.W.; Taylor, J.M.; Davison, A.S.; Xu, Y.; Grixti, J.M.; Gotts, N.; Sorokin, A.; Goodacre, R.; Kell, D.B. Untargeted metabolomics of COVID-19 patient serum reveals potential prognostic markers of both severity and outcome. medRxiv 2020. [Google Scholar] [CrossRef]
- Migaud, M.; Gandotra, S.; Chand, H.S.; Gillespie, M.N.; Thannickal, V.J.; Langley, R.J. Metabolomics to Predict Antiviral Drug Efficacy in COVID-19. Am. J. Respir. Cell Mol. Biol. 2020, 63, 396–398. [Google Scholar] [CrossRef]
- Meoni, G.; Ghini, V.; Maggi, L.; Vignoli, A.; Mazzoni, A.; Salvati, L.; Capone, M.; Vanni, A.; Tenori, L.; Fontanari, P.; et al. Metabolomic/lipidomic profiling of COVID-19 and individual response to tocilizumab. PLoS Pathog. 2021, 17, e1009243. [Google Scholar] [CrossRef]
- Gowda, G.A.N.; Gowda, Y.N.; Raftery, D. Expanding the Limits of Human Blood Metabolite Quantitation Using NMR Spectroscopy. Anal. Chem. 2015, 87, 706–715. [Google Scholar] [CrossRef] [Green Version]
- Wishart, D.S.; Feunang, Y.D.; Marcu, A.; Guo, A.C.; Liang, K.; Vázquez-Fresno, R.; Sajed, T.; Johnson, D.; Allison, P.; Karu, N.; et al. HMDB 4.0: The human metabolome database for 2018. Nucleic Acids Res. 2018, 46, D608–D617. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Wishart, D.S. Using MetaboAnalyst 3.0 for Comprehensive Metabolomics Data Analysis. Curr. Protoc. Bioinform. 2016, 55, 14-10. [Google Scholar] [CrossRef]
- Liu, M.; Tang, H.; Niholson, J.K.; Lindon, J.C. Use of 1H NMR-determined diffusion coefficients to characterise lipoprotein fractions in human blood plasma. Magn. Reson. Chem. 2002, 40, S83–S88. [Google Scholar] [CrossRef]
- Gromski, P.S.; Muhamadali, H.; Ellis, D.I.; Xu, Y.; Correa, E.; Turner, M.L.; Goodacre, R. A tutorial review: Metabolomics and partial least squares-discriminant analysis—A marriage of convenience or a shotgun wedding. Anal. Chim. Acta 2015, 879, 10–23. [Google Scholar] [CrossRef]
- Xia, J.; Broadhurst, D.I.; Wilson, M.; Wishart, D.S. Translational biomarker discovery in clinical metabolomics: An introductory tutorial. Metabolomics 2012, 9, 280–299. [Google Scholar] [CrossRef] [Green Version]
- Tamez-Pérez, H.E.; Quintanilla-Flores, D.L.; Rodríguez-Gutiérrez, R.; González-González, J.G.; Tamez-Peña, A.L. Steroid hyperglycemia: Prevalence, early detection and therapeutic recommendations: A narrative review. World J. Diabetes 2015, 6, 1073–1078. [Google Scholar] [CrossRef]
- Pasternak, J.J.; McGregor, D.G.; Lanier, W.L. Effect of Single-Dose Dexamethasone on Blood Glucose Concentration in Patients Undergoing Craniotomy. J. Neurosurg. Anesthesiol. 2004, 16, 122–125. [Google Scholar] [CrossRef]
- Rayman, G.; Lumb, A.N.; Kennon, B.; Cottrell, C.; Nagi, D.; Page, E.; Voigt, D.; Courtney, H.C.; Atkins, H.; Higgins, K.; et al. Dexamethasone therapy in COVID-19 patients: Implications and guidance for the management of blood glucose in people with and without diabetes. Diabet. Med. 2021, 38, e14378. [Google Scholar] [CrossRef] [PubMed]
- Brufsky, A. Hyperglycemia, hydroxychloroquine, and the COVID-19 pandemic. J. Med Virol. 2020, 92, 770–775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butkowski, E.G.; Jelinek, H.F. Hyperglycaemia, oxidative stress and inflammatory markers. Redox Rep. 2016, 22, 257–264. [Google Scholar] [CrossRef] [Green Version]
- Erener, S. Diabetes, infection risk and COVID-19. Mol. Metab. 2020, 39, 101044. [Google Scholar] [CrossRef]
- Puchalska, P.; Crawford, P.A. Multi-dimensional Roles of Ketone Bodies in Fuel Metabolism, Signaling, and Therapeutics. Cell Metab. 2017, 25, 262–284. [Google Scholar] [CrossRef] [Green Version]
- Fu, S.P.; Li, S.N.; Wang, J.F.; Li, Y.; Xie, S.S.; Xue, W.J.; Liu, H.M.; Huang, B.X.; Lv, Q.K.; Lei, L.C.; et al. BHBA suppresses LPS-induced inflammation in BV-2 cells by inhibiting NF-κB activation. Mediat. Inflamm. 2014, 2014, 983401. [Google Scholar] [CrossRef] [Green Version]
- Rahman, M.; Muhammad, S.; Khan, M.A.; Chen, H.; Ridder, D.A.; Muller-Fielitz, H.; Pokorna, B.; Vollbrandt, T.; Stolting, I.; Nadrowitz, R.; et al. The B-hydroxybutyrate receptor HCA2 activates a neuroprotective subset of macrophages. Nat. Commun. 2014, 5, 3944. [Google Scholar] [CrossRef] [PubMed]
- Youm, Y.; Nguyen, K.Y.; Grant, R.W.; Goldberg, E.L.; Bodogai, M.; Kim, D.; D’agostino, D.; Planavsky, N.; Lupfer, C.; Kanneganti, T.D.; et al. The ketone metabolite beta-hydroxybutyrate blocks NLRP3 inflammasome-mediated inflammatory disease. Nat. Med. 2015, 21, 263–269. [Google Scholar] [CrossRef] [Green Version]
- Shi, X.; Li, X.; Li, D.; Li, Y.; Song, Y.; Deng, Q.; Wang, J.; Zhang, Y.; Ding, H.; Yin, L.; et al. β-Hydroxybutyrate Activates the NF-κB Signaling Pathway to Promote the Expression of Pro-Inflammatory Factors in Calf Hepatocytes. Cell Physiol. Biochem. 2014, 33, 920–932. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wang, X.; Chen, J.; Zuo, X.; Zhang, H.; Deng, A. COVID -19 infection may cause ketosis and ketoacidosis. Diabetes Obes. Metab. 2020, 22, 1935–1941. [Google Scholar] [CrossRef] [PubMed]
- Salminen, A.; Kauppinen, A.; Hiltunen, M.; Kaarniranta, K. Krebs cycle intermediates regulate DNA and histone methylation: Epigenetic impact on the aging process. Ageing Res. Rev. 2014, 16, 45–65. [Google Scholar] [CrossRef] [PubMed]
- Dierckx, T.; van Elslande, J.; Salmela, H.; Decru, B.; Wauters, E.; Gunst, J.; Van, H.Y.; Wauters, J.; Stessel, B.; Vermeersch, P.; et al. The metabolic fingerprint of COVID-19 severity. medRxiv 2020. [Google Scholar] [CrossRef]
- Zhenyukh, O.; Civantos, E.; Ruiz-Ortega, M.; Sánchez, M.S.; Vázquez, C.; Peiró, C.; Egido, J.; Mas, S. High concentration of branched-chain amino acids promotes oxidative stress, inflammation and migration of human peripheral blood mononuclear cells via mTORC1 activation. Free. Radic. Biol. Med. 2017, 104, 165–177. [Google Scholar] [CrossRef]
- HoleČek, M. Branched-chain amino acids in health and disease: Metabolism, alterations in blood plasma, and as supplements. Nutr. Metab. 2018, 15, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larsson, S.C.; Markus, H.S. Branched-chain amino acids and Alzheimer’s disease: A Mendelian randomisation analysis. Sci. Rep. 2017, 7, 13604. [Google Scholar] [CrossRef] [Green Version]
- Holeček, M. Why Are Branched-Chain Amino Acids Increased in Starvation and Diabetes? Nutrients 2020, 12, 3087. [Google Scholar] [CrossRef]
- Cruzat, V.; Rogero, M.M.; Keane, K.N.; Curi, R.; Newsholme, P. Glutamine: Metabolism and Immune Function, Supplementation and Clinical Translation. Nutrients 2018, 10, 1564. [Google Scholar] [CrossRef] [Green Version]
- Cengiz, M.; Uysal, B.B.; Ikitimur, H.; Ozcan, E.; Islamoğlu, M.S.; Aktepe, E.; Yavuzer, H.; Yavuzer, S. Effect of oral l-Glutamine supplementation on Covid-19 treatment. Clin. Nutr. Exp. 2020, 33, 24–31. [Google Scholar] [CrossRef]
- De Oliveira, D.C.; Lima, F.D.S.; Sartori, T.; Santos, A.C.A.; Rogero, M.M.; Fock, R.A. Glutamine metabolism and its effects on immune response: Molecular mechanism and gene expression. Nutrients 2016, 41, 14. [Google Scholar] [CrossRef] [Green Version]
- Shah, A.M.; Wang, Z.; Ma, J. Glutamine Metabolism and Its Role in Immunity, a Comprehensive Review. Animals 2020, 10, 326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdelaal, M.A.; Abdelrahman, D.; Cengiz, M.; Yavuzer, H.; Yavuzer, S.; Bien, I.; Bhuva, P.; Pham, J.V.; Siu, R.; Tang, M.; et al. Actions of L-Glutamine vs. COVID-19 Suggest Additional Benefit in Sickle Cell Disease. Blood 2020, 136, 11–12. [Google Scholar] [CrossRef]
- Murr, C.; Grammer, T.B.; Meinitzer, A.; Kleber, M.E.; März, W.; Fuchs, D. Immune Activation and Inflammation in Patients with Cardiovascular Disease Are Associated with Higher Phenylalanine to Tyrosine Ratios: The Ludwigshafen Risk and Cardiovascular Health Study. J. Amino Acids 2014, 2014, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Geisler, S.; Gostner, J.M.; Becker, K.; Ueberall, F.; Fuchs, D. Immune activation and inflammation increase the plasma phenylalanine-to-tyrosine ratio. Pteridines 2013, 24, 27–31. [Google Scholar] [CrossRef]
- Barberis, E.; Timo, S.; Amede, E.; Vanella, V.V.; Puricelli, C.; Cappellano, G.; Raineri, D.; Cittone, M.G.; Rizzi, E.; Pedrinelli, A.R.; et al. Large-Scale plasma analysis revealed new mechanisms and molecules associated with the host response to SARS-CoV-2. Int. J. Mol. Sci. 2020, 21, 8623. [Google Scholar] [CrossRef]
- Huang, C.; Huang, L.; Wang, Y.; Li, X.; Ren, L.; Gu, X.; Kang, L.; Guo, L.; Liu, M.; Zhou, X.; et al. 6-month consequences of COVID-19 in patients discharged from hospital: A cohort study. Lancet 2021, 397, 220–232. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter: Median (IQR) | Group A | Group B |
|---|---|---|
| size | 34 | 19 |
| age, years | 65 (21) | 71 (16) |
| gender | 15 female | 8 female |
| number of samples Day 1 | 34 | 19 |
| number of samples Day 3 | 31 | 16 |
| number of samples Day 7 | 26 | 10 |
| oxygen | 34 | 19 |
| HFNO | - | 7 |
| NIV | 1 * | 2 |
| smoker | 2 | 2 |
| non smoker | 23 | 10 |
| ex-smoker | 5 | 5 |
| smoking not known | 4 | 2 |
| chronic obstructive pulmonary disease | 2 | 4 |
| obesity | 11 | 11 |
| hypertension | 22 | 13 |
| asthma | 2 | - |
| kidney disease | 4 | 4 |
| ischemic heart disease | 9 | 9 |
| diabetes | 14 | 7 |
| cancer | 1 | 2 |
| cancer history | 1 | 3 |
| thyroid disease | 3 | 1 |
| liver cirrhosis | - | 1 |
| rheumatoid arthritis | 1 | 3 |
| stroke history | 1 | 1 |
| acute stroke | 1 | - |
| sarcodiosis | 1 | - |
| Metabolite | NMR Peak Assignment, Confirmed by Jres and Cosy |
|---|---|
| lactate | 1.33 (d; J = 7.0), 4.12 (q; J = 7.0) |
| glutamine | 2.12 (m), 2.15 (m), 2.44 (m), 2.48 (m), 3.77 (dd) |
| isoleucine | 0.94 (t; J = 7.5), 1.01 (d; J = 7.0), 3.68 (d; J = 4.2) |
| leucine | 0.96 (d; J = 6.2), 0.97 (d; J = 6.1), 1.68 (m), 1.72 (m), 1.75(m) |
| phenylalanine | 3.13 (m), 3.28 (m), 7.34 (d; J = 7.5), 7.38 (t; J = 7.4), 7.44 (t) |
| tyrosine | 3.05 (dd), 3.20 (dd), 3.93 (dd), 6.91 (d; J = 8.5), 7.20 (d; J = 8.5) |
| valine | 0.99 (d; J = 7.1), 1.04 (d; J = 7.1), 2.27 (m), 3.61 (d; J = 4.4) |
| pyruvate | 2.38 (s) |
| citrate | 2.54 (d), 2.67 (d) |
| acetate | 1.92 (s) |
| alanine | 1.48 (d; J = 7.30), 3.78 (q) |
| glucose | 3.23 (m), 3.40 (m), 3.46 (m), 3.52 (dd), 3.78 (m), 3.82 (m), 3.89 (dd), 4.64 (d), 5.23 (d) |
| 3-hydroxybutyrate | 1.20 (d; J = 6.23 Hz), 2.31 (m), 2.41 (m), 4.16 (m) |
| creatine | 3.04 (s), 3.94 (s) |
| lysine | 1.33 (d), 3.58 (d; J = 4.9), 4.25 (m) |
| 2-oxoisocapronate (2-ketoleucine) | 0.94 (d; J = 6.6), 2.11 (m), 2.61 (d; J = 7.0) |
| α-ketoisovalerate (2-ketovaline) | 1.11 (d; J = 7.1), 3.01(dq) |
| 3-methyl-2-oxo-valerate (2-ketoisoleucine) | 0.90 (t; J = 7.5), 1.10 (d; J = 6.7) |
| lipoprotein fraction | 0.82–0.93 (m), 1.20–1.37 (m) |
| # creatinine | 3.05 (s), 4.07 (s) |
| # histidine | 7.07 (s), 7.80 (s) |
| # proline | 1.46 (m), 1.50 (m), 1.73 (m), 1.89 (m), 1.93 (m), 3.03 (t; J = 7.6) |
| # threonine | 1.34 (d), 3.56 (d;J = 4.9),4.26 (m) |
| # tryptophan | 7.21 (t), 7.30 (t), 7.33 (s), 7.56 (d), 7.74 (d; J = 8.0) |
| System | OOB Error (5 Variables) | AUC | Number of Variables | Metabolites in Importance Order |
|---|---|---|---|---|
| All Patients Day 1/Controls | 3/108 | 0.984 0.995 | 2 5 | 3-hydroxybutyrate, phenylalanine, Phe/Tyr ratio, acetate, glucose |
| Group A Day 1/Controls | 1/89 | 0.977 0.996 | 2 5 | 3-hydroxybutyrate, phenylalanine, glucose, Phe/Tyr ratio, acetate, |
| Group B Day 1/Controls | 1/74 | 0.972 0.991 | 2 5 | phenylalanine, 3-hydroxybutyrate, Phe/Tyr ratio, acetate, glutamine or glucose |
| Group A/Group B Day1 | - | 0.568 0.674 | 2 5 | AUC value too low |
| Group A/Group B Day 3 | - | 0.754 0.783 | 2 5 | alanine, lysine, glutamine, Phe/Tyr ratio, phenylalanine |
| Group A/Group B Day 7 | - | 0.487 0.503 | 2 5 | AUC value too low |
| Metabolite | Nonparametric ANOVA (Kruskal–Wallis) | Mann–Whitney U-test, Only Significant Changes (p < 0.05) are Listed, Arrows Indicate the Direction of Change | ||||
|---|---|---|---|---|---|---|
| chi. Squared | p-Value | FDR p-Value Adjusted | Group A Against Controls | Group B Against Controls | Group A Against GroupB | |
| glucose | 80 | 3.9 × 10−15 | 1.4 × 10−14 | Day1↑, Day3↑ | Day1↑, Day3↑, Day7↑ | |
| 3-OH-butyrate | 130 | 1.1 × 10−24 | 1.2 × 10−23 | Day1↑, Day3↑, Day7↑ | Day1↑, Day3↑, Day7↑ | Day7, A < B |
| citrate | 77 | 1.2 × 10−14 | 3.2 × 10−14 | Day1↓, Day3↓, Day7↓ | Day1↓, Day3↓, Day7↓ | |
| leucine | 39 | 7.6 × 10−7 | 1.5 × 10−6 | Day1↑, Day3↑, Day7↑ | Day1↑, Day3↑, Day7↑ | |
| isoleucine | 31 | 2.8 × 10−5 | 4.9 × 10−5 | Day1↑, Day3↑, Day7↑ | Day1↑, Day3↑, Day7↑ | |
| valine | 13 | 0.040 | 0.047 | Day1↑, Day3↑, Day7↑ | Day1↑, Day3↑, Day7↑ | |
| ketoleucine | 25 | 3.7 × 10−4 | 5.5 × 10−4 | Day1↑, Day3↓, Day7↓ | Day1, A > B | |
| ketoisoleucine | 19 | 0.0042 | 0.0059 | Day1↑, Day3↓, Day7↓ | ||
| ketovaline | 17 | 0.0094 | 0.012 | Day1↑, Day3↓, Day7↓ | Day1, A > B | |
| creatine | 64 | 8.5 × 10−12 | 2.0 × 10−11 | Day3↑, Day7↑ | Day3↑, Day7↑ | |
| alanine | 49 | 8.5 × 10−9 | 1.8 × 10−8 | Day1↓ | Day1↓, Day3↓, Day7↓ | Day3, Day7, A > B |
| glutamine | 29 | 6.1 × 10−5 | 9.8 × 10−5 | Day1↓ | Day1↓,Day3↓ | |
| phenylalanine | 120 | 1.2 × 10−23 | 8.5 × 10−23 | Day1↑, Day3↑, Day7↑ | Day1↑, Day3↑, Day7↑ | |
| Phe/Tyr ratio | 85 | 3.3 × 10−16 | 1.4 × 10−15 | Day1↑, Day3↑, Day7↑ | Day1↑, Day3↑, Day7↑ | Day1, A < B |
| lipoproteins | 150 | 7.8 × 10−31 | 1.6 × 10−29 | Day1↓, Day3↓, Day7↓ | Day1↓, Day3↓, Day7↓ | |
| acetate | 100. | 2.1 × 10−19 | 1.1 × 10−18 | Day1↓, Day3↓, Day7↓ | Day1↓, Day3↓, Day7↓ | |
| lysine | 79 | 6.6 × 10−15 | 2.0 × 10−14 | Day1↑, Day3↑, Day7↑ | Day7↑ | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baranovicova, E.; Bobcakova, A.; Vysehradsky, R.; Dankova, Z.; Halasova, E.; Nosal, V.; Lehotsky, J. The Ability to Normalise Energy Metabolism in Advanced COVID-19 Disease Seems to Be One of the Key Factors Determining the Disease Progression—A Metabolomic NMR Study on Blood Plasma. Appl. Sci. 2021, 11, 4231. https://doi.org/10.3390/app11094231
Baranovicova E, Bobcakova A, Vysehradsky R, Dankova Z, Halasova E, Nosal V, Lehotsky J. The Ability to Normalise Energy Metabolism in Advanced COVID-19 Disease Seems to Be One of the Key Factors Determining the Disease Progression—A Metabolomic NMR Study on Blood Plasma. Applied Sciences. 2021; 11(9):4231. https://doi.org/10.3390/app11094231
Chicago/Turabian StyleBaranovicova, Eva, Anna Bobcakova, Robert Vysehradsky, Zuzana Dankova, Erika Halasova, Vladimir Nosal, and Jan Lehotsky. 2021. "The Ability to Normalise Energy Metabolism in Advanced COVID-19 Disease Seems to Be One of the Key Factors Determining the Disease Progression—A Metabolomic NMR Study on Blood Plasma" Applied Sciences 11, no. 9: 4231. https://doi.org/10.3390/app11094231