
In Vitro Cytotoxic Evaluation and Apoptotic Effects of Datura innoxia Grown in Saudi Arabia and Phytochemical Analysis

 , , , , ,
, , , , ,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Collection and Authentication
2.2. Cell Viability (MTT Assay)
2.3. Cell Cycle Analysis
2.4. FITC Annexin V/PI Apoptosis Detection
2.5. Gene Quantification by qRT-PCR
2.6. Hoechst 33258 Staining
2.7. GC-MS Analysis of D. innoxia Constituents
2.8. Statistical Analysis
3. Results
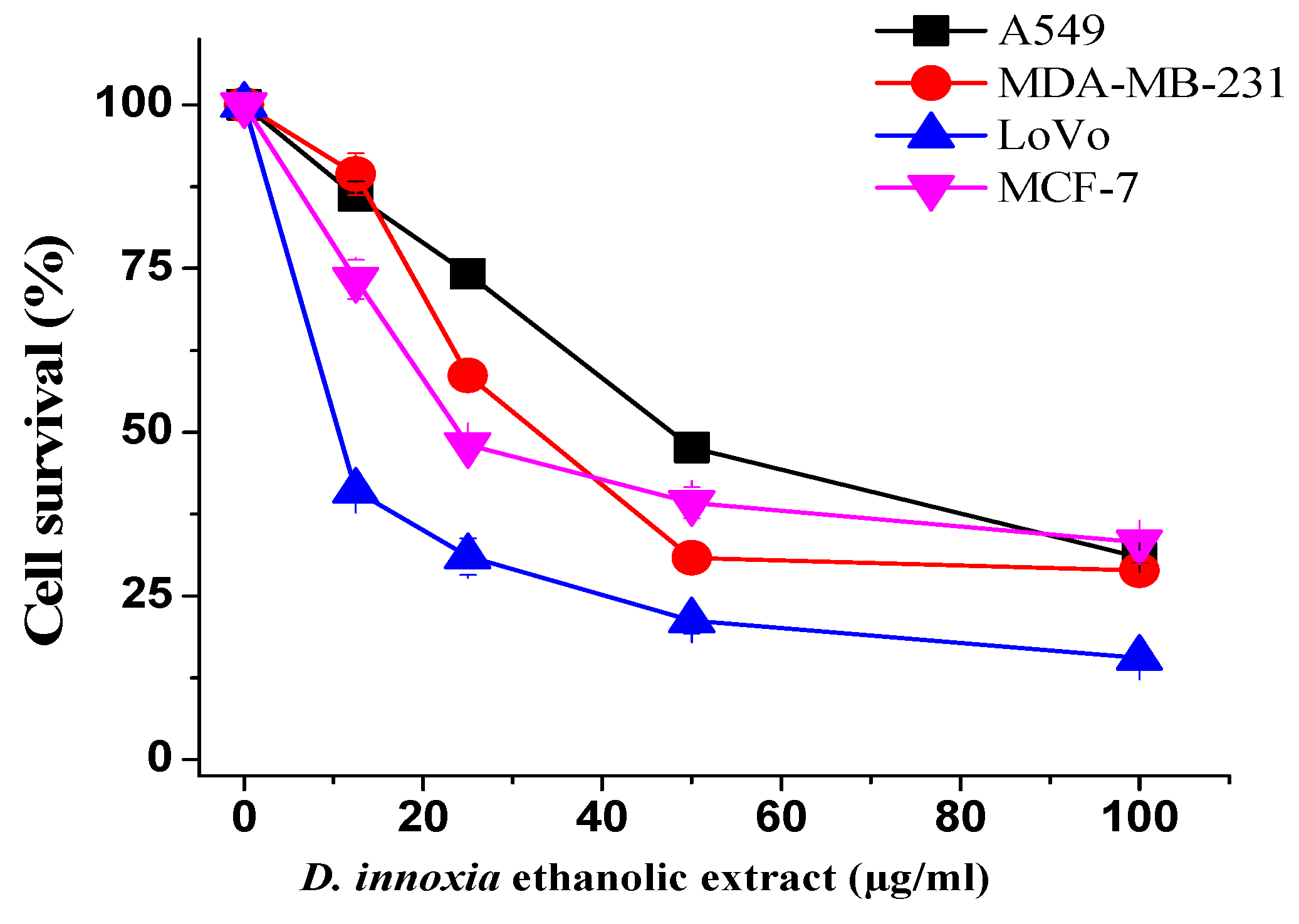
3.1. Ethanol Extract of D. innoxia Inhibit Cells Proliferation
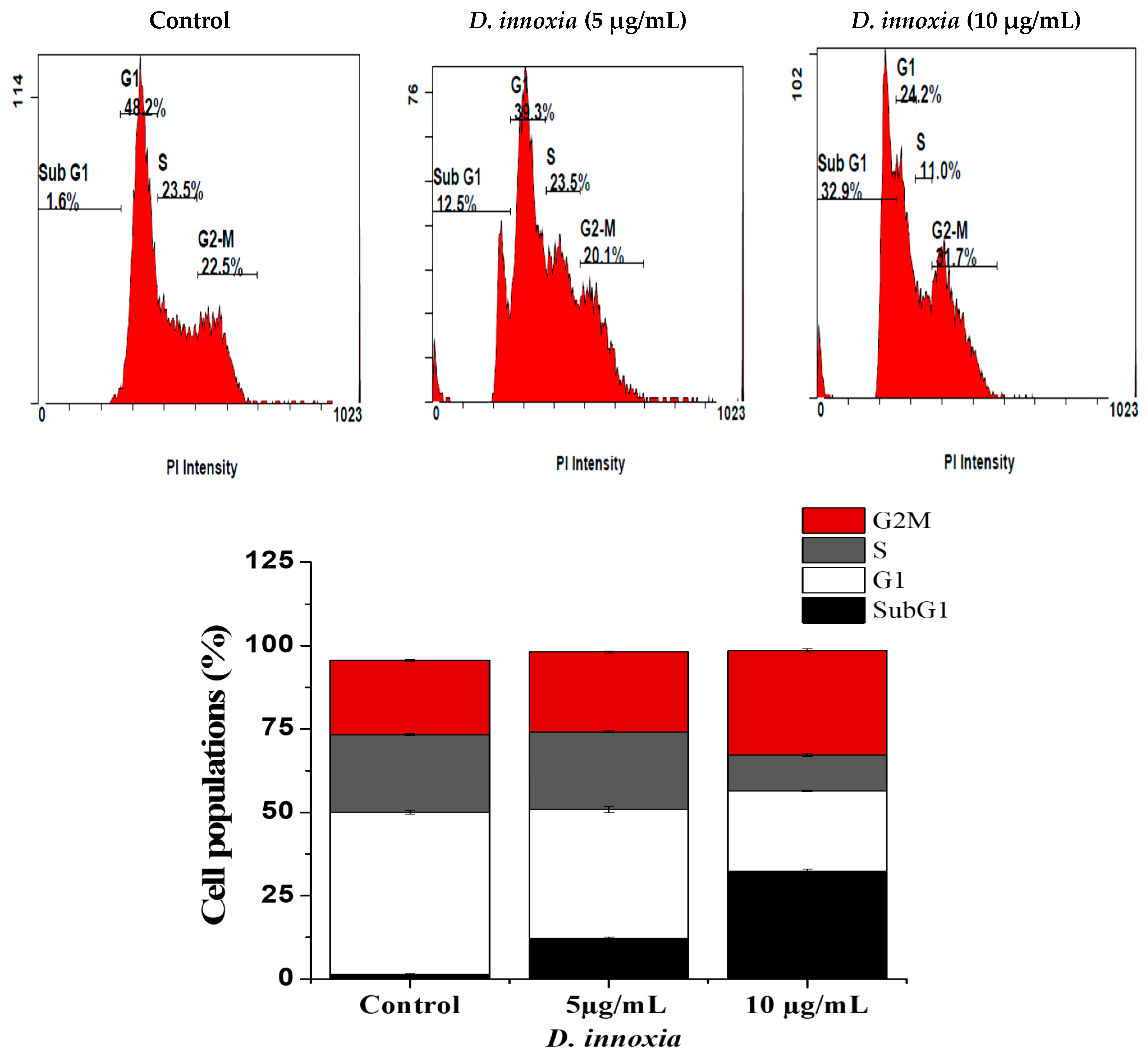
3.2. D. innoxia Causes a Sub-G1 Cells Accumulation
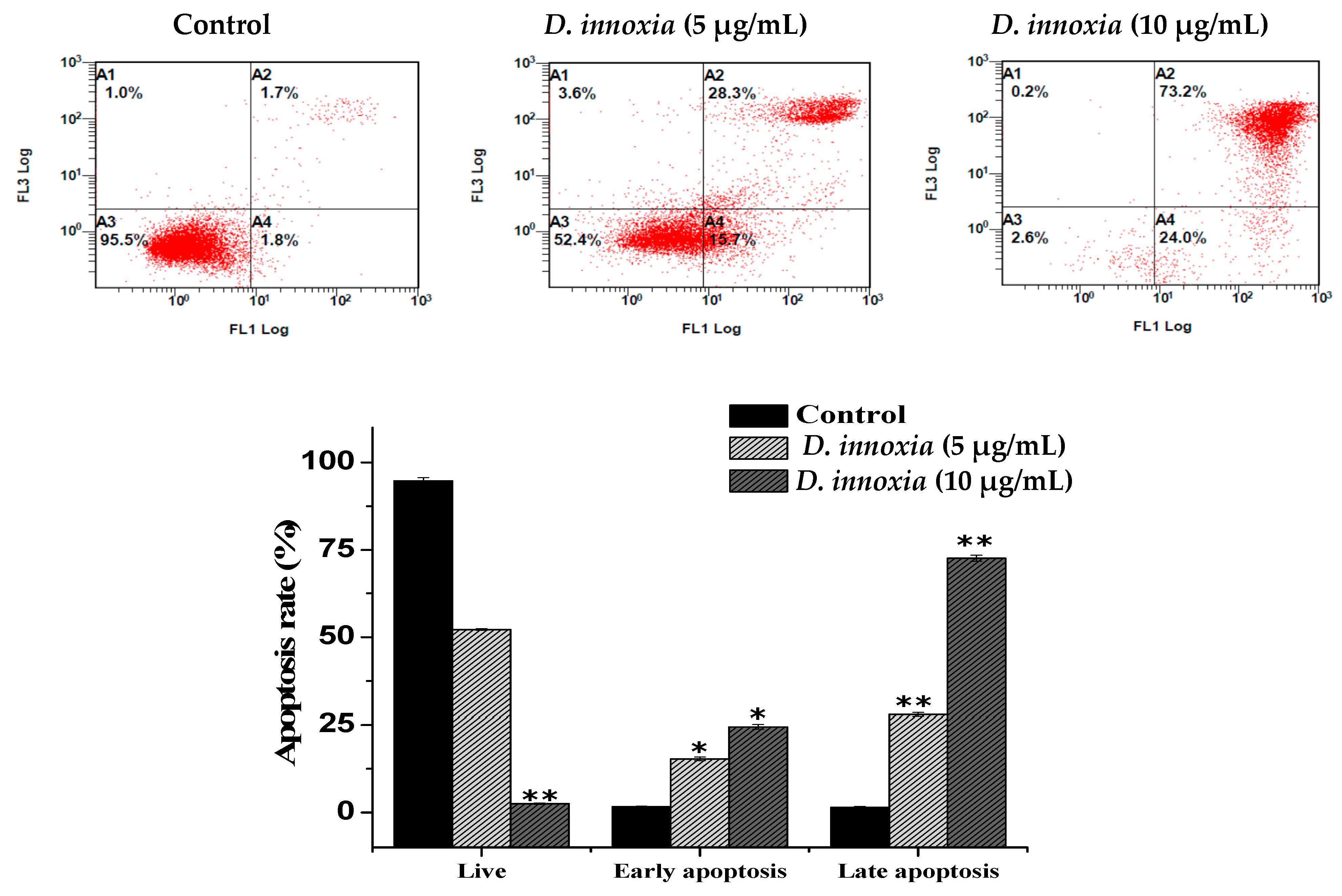
3.3. Apoptotic Cell Death Quantification of D. innoxia Treated Cells
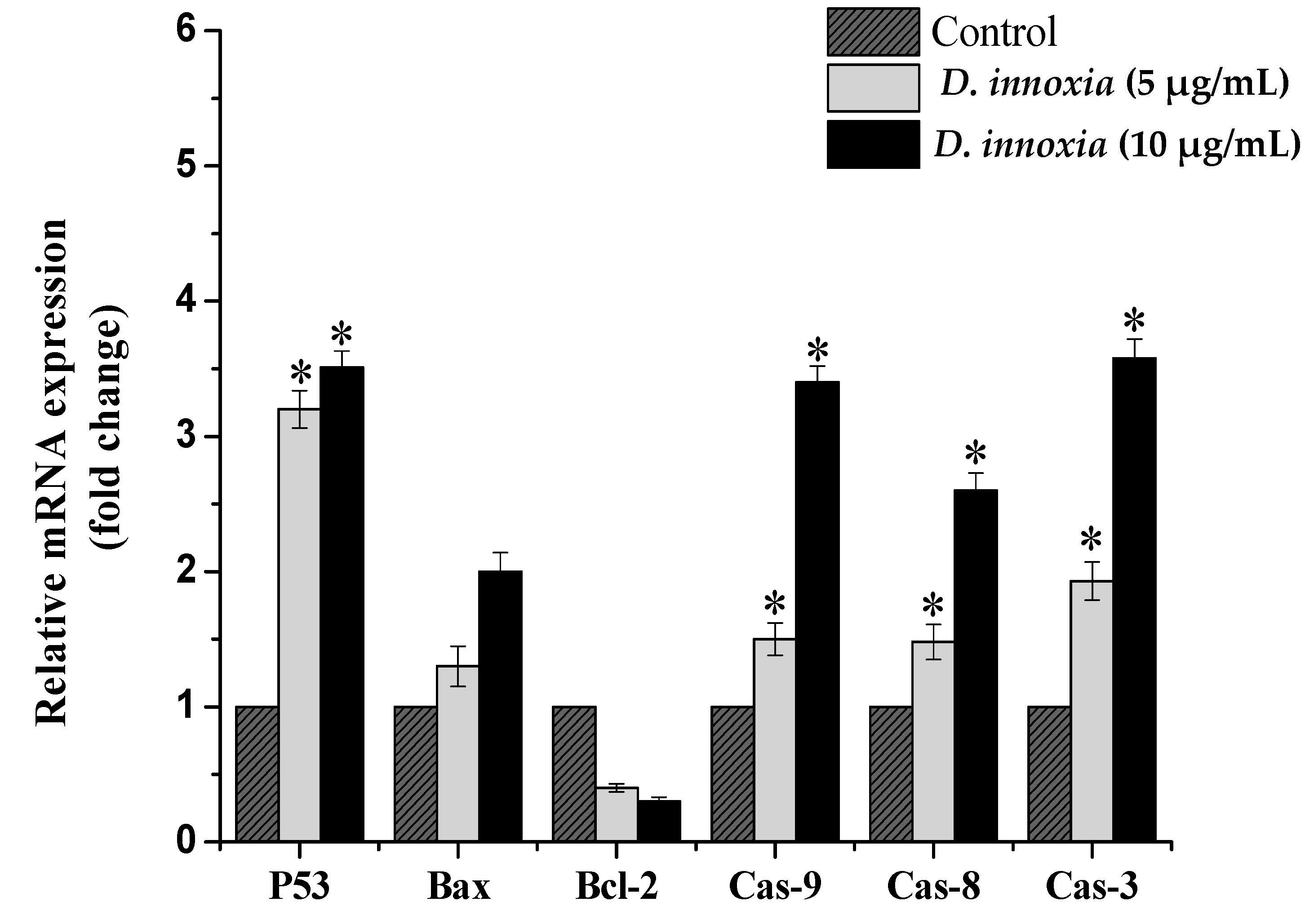
3.4. The Quantitative Real-Time PCR Analysis of Apoptosis-Related Genes
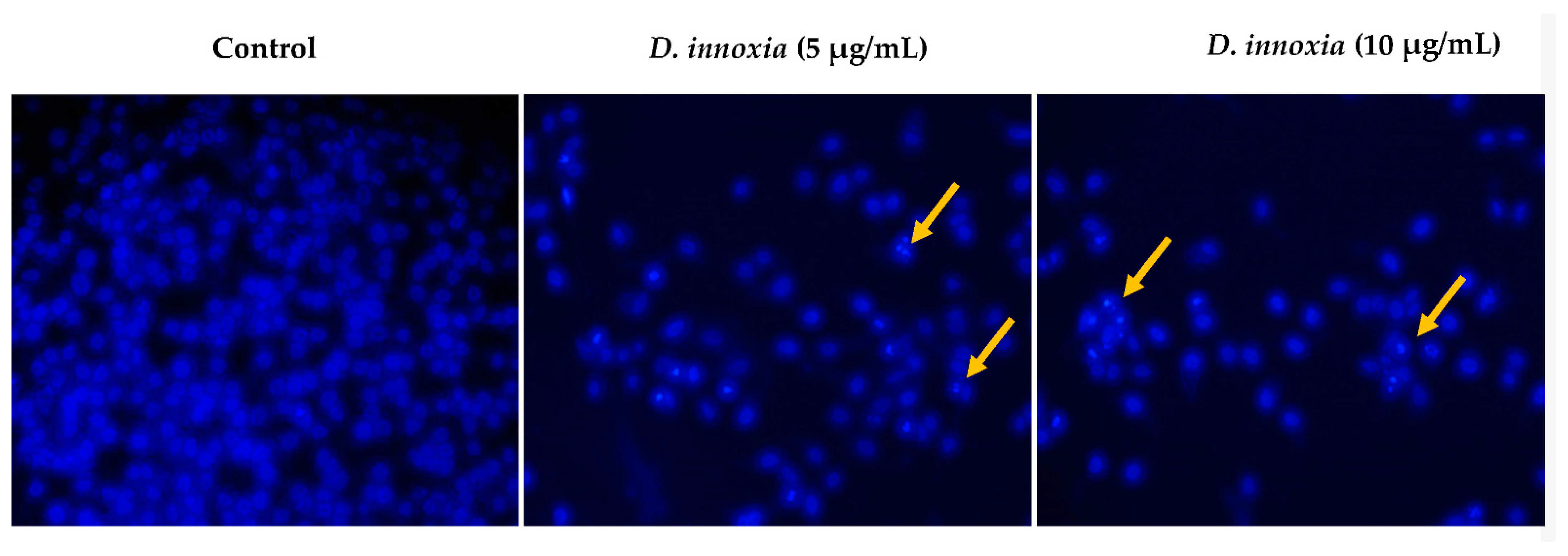
3.5. D. innoxia Causes a DNA Fragmentation
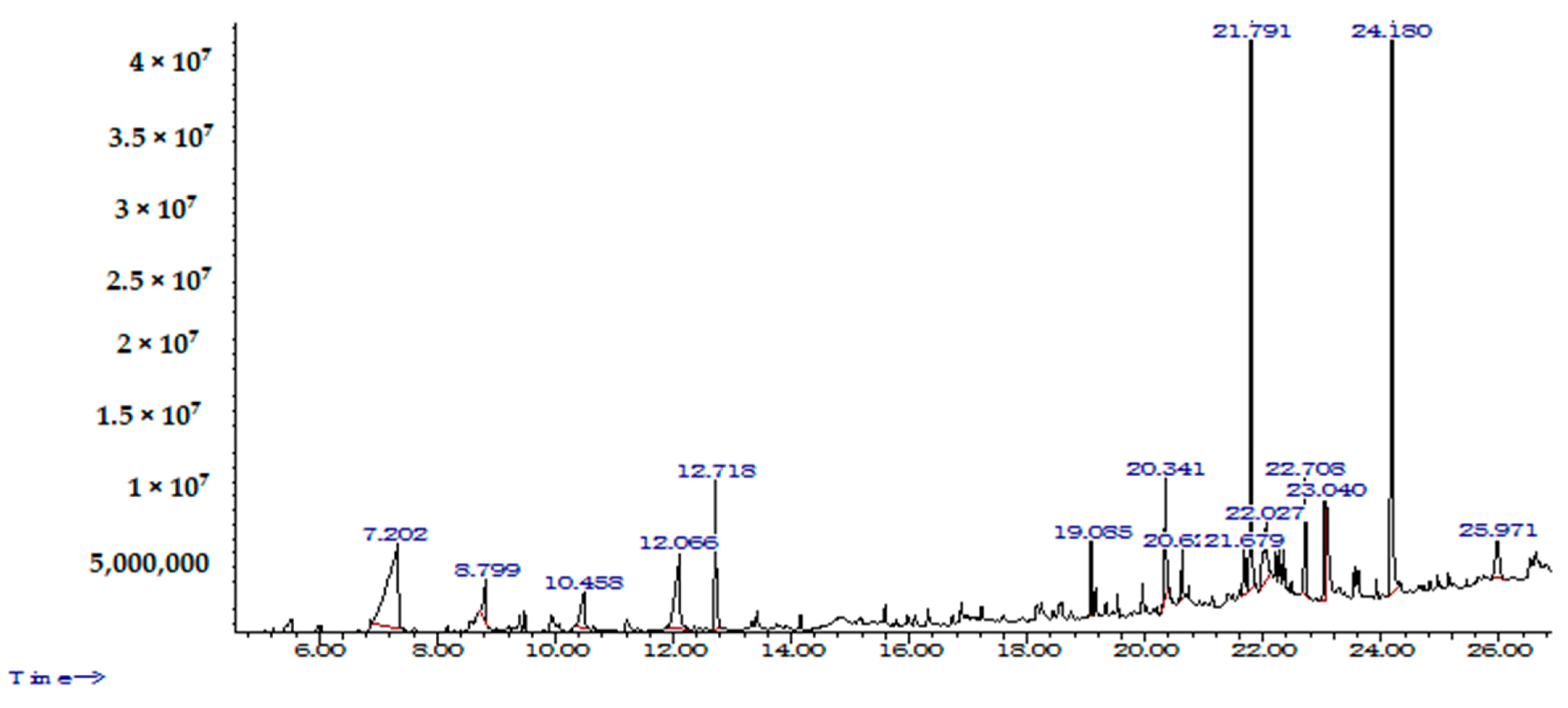
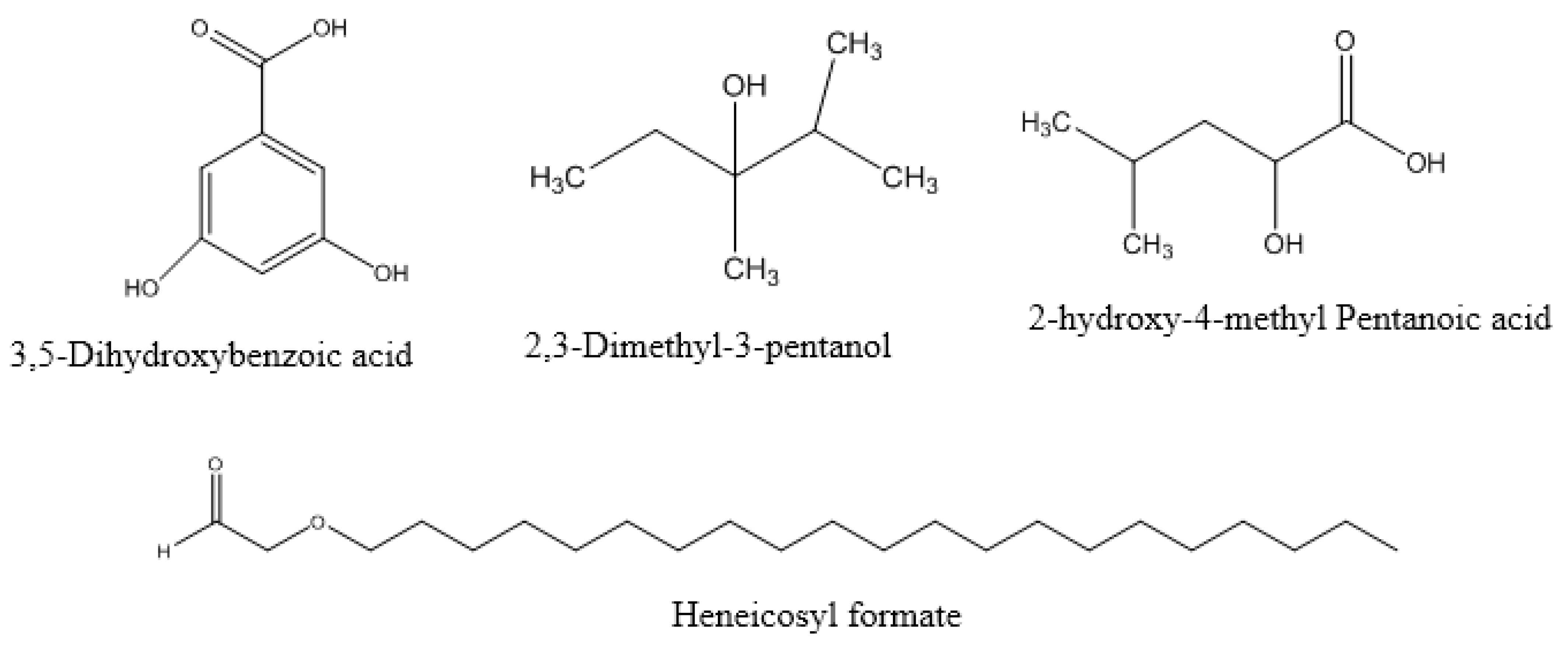
3.6. Identification of D. innoxia Components by GC-MS
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021. [Google Scholar] [CrossRef]
- Zhang, Q.-Y.; Wang, F.-X.; Jia, K.-K.; Kong, L.-D. Natural Product Interventions for Chemotherapy and Radiotherapy-Induced Side Effects. Front. Pharm. 2018, 9, 1253. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, J.; Abbasi, B.A.; Mahmood, T.; Kanwal, S.; Ali, B.; Shah, S.A.; Khalil, A.T. Plant-derived anticancer agents: A green anticancer approach. Asian Pac. J. Trop. Biomed. 2017, 7, 1129–1150. [Google Scholar] [CrossRef]
- Desai, A.G.; Qazi, G.N.; Ganju, R.K.; El-Tamer, M.; Singh, J.; Saxena, A.K.; Bedi, Y.S.; Taneja, S.C.; Bhat, H.K. Medicinal plants and cancer chemoprevention. Curr. Drug Metab. 2008, 9, 581–591. [Google Scholar] [CrossRef] [Green Version]
- Al-Eisawi, D.M.; Al-Ruzayza, S. The flora of holy Mecca district, Saudi Arabia. Int. J. Biodivers. Conserv. 2015, 7, 173–189. [Google Scholar]
- Al-Zahrani, A.A. Saudi anti-human cancer plants database (SACPD): A collection of plants with anti-human cancer activities. Oncol. Rev. 2018, 12, 349. [Google Scholar] [CrossRef] [Green Version]
- Evans, W.; Ghani, A.; Woolley, V.A. Distribution of littorine and other alkaloids in the roots of Datura species. Phytochemistry 1972, 11, 2527–2529. [Google Scholar] [CrossRef]
- Pandey, M.; Saraswati, S.; Agrawal, S. Antiproliferative effects of Datura innoxia extract in cervical HeLa cell line. J. Pharm. Res. 2011, 4, 1124–1126. [Google Scholar]
- Giral, F.; Hidalgo, C. Presence of alkaloids in Mexican plants. Int. J. Crude Drug Res. 1983, 21, 1–13. [Google Scholar] [CrossRef]
- El-Tawil, B. Chemical constituents of indigenous plants used in native medicine of Saudi Arabia. II. Arab Gulf J. Sci. Res. 1983. [Google Scholar]
- Aynehchi, Y.; Salehi Sormaghi, M.; Amin, G.; Khoshkhow, M.; Shabani, A. Survey of Iranian plants for saponins, alkaloids, flavonoids and tannins. III. Int. J. Crude Drug Res. 1985, 23, 33–41. [Google Scholar] [CrossRef]
- Shama, A.I.; Abd-Kreem, Y.; Fadowa, A.; Samar, R.; Sabahelkhier, M. In vitro antibacterial and antifungal activity and Datura innoxia extracts. Int. J. Environ. 2014, 3, 173–185. [Google Scholar] [CrossRef] [Green Version]
- Gajendran, B.; Durai, P.; Varier, K.M.; Liu, W.; Li, Y.; Rajendran, S.; Nagarathnam, R.; Chinnasamy, A. Green synthesis of silver nanoparticle from Datura inoxia flower extract and its cytotoxic activity. BioNanoScience 2019, 9, 564–572. [Google Scholar] [CrossRef]
- Cook, N.C.; Samman, S. Flavonoids—chemistry, metabolism, cardioprotective effects, and dietary sources. J. Nutr. Biochem. 1996, 7, 66–76. [Google Scholar] [CrossRef]
- Manach, C.; Williamson, G.; Morand, C.; Scalbert, A.; Rémésy, C. Bioavailability and bioefficacy of polyphenols in humans. I. Review of 97 bioavailability studies. Am. J. Clin. Nutr. 2005, 81, 230S–242S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hostettmann, K.; Marston, A. Saponins; Cambridge University Press: Cambridge, UK, 2005. [Google Scholar]
- Cheshomi, H.; Aldaghi, L.S.; Rezaei Seresht, H. Cytotoxicity of the methanol extract of datura innoxia petals on MCF-7 and HEK-293 cell lines. J. Biomed. 2016, 1, e6623. [Google Scholar] [CrossRef] [Green Version]
- Chamani, E.; Ebrahimi, R.; Khorsandi, K.; Meshkini, A.; Zarban, A.; Sharifzadeh, G. In vitro cytotoxicity of polyphenols from Datura innoxia aqueous leaf-extract on human leukemia K562 cells: DNA and nuclear proteins as targets. Drug Chem. Toxicol. 2020, 43, 138–148. [Google Scholar] [CrossRef]
- Nasr, F.A.; Noman, O.M.; Alqahtani, A.S.; Qamar, W.; Ahamad, S.R.; Al-Mishari, A.A.; Alyhya, N.; Farooq, M. Phytochemical constituents and anticancer activities of Tarchonanthus camphoratus essential oils grown in Saudi Arabia. Saudi Pharm. J. 2020, 28, 1474–1480. [Google Scholar] [CrossRef] [PubMed]
- Rio, D.C.; Ares, M., Jr.; Hannon, G.J.; Nilsen, T.W. Purification of RNA using TRIzol (TRI reagent). Cold Spring Harb. Protoc. 2010, 2010. [Google Scholar] [CrossRef]
- Abutaha, N.; Nasr, F.A.; Al-zharani, M.; Alqahtani, A.S.; Noman, O.M.; Mubarak, M.; Abdelhabib, S.; Wadaan, M.A. Effects of hexane root extract of ferula hermonis boiss. On human breast and colon cancer cells: An in vitro and in vivo study. Biomed. Res. Int. 2019, 2019, 3079895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McLafferty, F.W.; Stauffer, D.B. The Wiley/NBS Registry of Mass Spectral Data; Wiley: New York, NY, USA, 1989; Volume 1. [Google Scholar]
- Adams, R.P. Identification of Essential oil Components by Gas Chromatography/Mass Spectrometry; Allured Publishing Corporation: Carol Stream, IL, USA, 2007; Volume 456. [Google Scholar]
- Jaafar, F.R.; Ajeena, S.J.; Mehdy, S.S. Anti-inflammatory impacts and analgesiac activity of aqueous extract Datura innoxia leaves against induced pain and inflammation in mice. J. Entomol. Zool. Stud. 2018, 6, 1894–1899. [Google Scholar]
- Arulvasu, C.; Babu, G.; Manikandan, R.; Srinivasan, P.; Sellamuthu, S.; Prabhu, D.; Dinesh, D. Anti-cancer effect of Datura innoxia P. Mill. Leaf extract in vitro through induction of apoptosis in human Colon Adenocarcinoma and larynx cancer cell lines. J. Pharm. Res. 2010, 3, 1485–1488. [Google Scholar]
- Fatima, H.; Khan, K.; Zia, M.; Ur-Rehman, T.; Mirza, B.; Haq, I.-u. Extraction optimization of medicinally important metabolites from Datura innoxia Mill.: An in vitro biological and phytochemical investigation. BMC Complement. Altern. Med. 2015, 15, 376. [Google Scholar] [CrossRef] [Green Version]
- Uğur, D.; Güneş, H.; Güneş, F.; Mammadov, R. Cytotoxic Activities of Certain Medicinal Plants on Different Cancer Cell Lines. Turk. J. Pharm. Sci. 2017, 14, 222–230. [Google Scholar] [CrossRef]
- Boik, J. Natural Compounds in Cancer Therapy; Oregon Medical Press: Princeton, MN, USA, 2001. [Google Scholar]
- Riccardi, C.; Nicoletti, I. Analysis of apoptosis by propidium iodide staining and flow cytometry. Nat. Protoc. 2006, 1, 1458–1461. [Google Scholar] [CrossRef] [PubMed]
- Demchenko, A.P. Beyond annexin V: Fluorescence response of cellular membranes to apoptosis. Cytotechnology 2013, 65, 157–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pistritto, G.; Trisciuoglio, D.; Ceci, C.; Garufi, A.; D’Orazi, G. Apoptosis as anticancer mechanism: Function and dysfunction of its modulators and targeted therapeutic strategies. Aging 2016, 8, 603–619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiraz, Y.; Adan, A.; Kartal Yandim, M.; Baran, Y. Major apoptotic mechanisms and genes involved in apoptosis. Tumour Biol. 2016, 37, 8471–8486. [Google Scholar] [CrossRef] [Green Version]
- Chamani, E.; Rezaei, Z.; Dastjerdi, K.; Javanshir, S.; Khorsandi, K.; Mohammadi, G.A. Evaluation of some genes and proteins involved in apoptosis on human chronic myeloid leukemia cells (K562 cells) by datura innoxia leaves aqueous extract. J. Biomol. Struct. Dyn. 2020, 38, 4838–4849. [Google Scholar] [CrossRef] [PubMed]
- Diker, D.; Markovitz, D.; Rothman, M.; Sendovski, U. Coma as a presenting sign of Datura stramonium seed tea poisoning. Eur. J. Intern. Med. 2007, 18, 336–338. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Nawale, L.; Sarkar, D.; Suresh, C. Two chitotriose-specific lectins show anti-angiogenesis, induces caspase-9-mediated apoptosis and early arrest of pancreatic tumor cell cycle. PLoS ONE 2016, 11, e0146110. [Google Scholar] [CrossRef] [PubMed]
- Iordache, A.; Culea, M.; Gherman, C.; Cozar, O. Characterization of some plant extracts by GC–MS. Nucl. Instrum. Methods Phys. Res. Sect. B Beam Interact. Mater. At. 2009, 267, 338–342. [Google Scholar] [CrossRef]
- Sankaranarayanan, R.; Valiveti, C.K.; Dachineni, R.; Kumar, D.R.; Lick, T.; Bhat, G.J. Aspirin metabolites 2,3-DHBA and 2,5-DHBA inhibit cancer cell growth: Implications in colorectal cancer prevention. Mol. Med. Rep. 2020, 21, 20–34. [Google Scholar] [CrossRef] [Green Version]
- Jigna, P.; Rathish, N.; Sumitra, C. Preliminary screening of some folklore medicinal plants from western India for potential antimicrobial activity. Indian J. Pharmacol. 2005, 37, 408–409. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Forward Primer Sequences | Reverse Primer Sequences | |
|---|---|---|---|
| 1 | P53 | 5′-TGGCTCTGACTGTACCACCATCC-3′ | 5′-CAGCTCTCGGAACATCTCGAAGC-3′ |
| 2 | Bax | 5′-GGA TGC GTC CAC CAA GAA G-3′ | 5′-CCT CTG CAG CTC CAT GTT AC-3′ |
| 3 | Bcl-2 | 5′-GTG GAT GAC TGA GTA CCT GAA C-3′ | 5′-GCC AGG AGA ATT CAA ACA GAG G-3′ |
| 4 | Caspase 9 | 5′-CAG GCC CCA TAT GAT CGA GG-3′ | 5′-TCG ACA ACT TTG CTG CTT GC-3′ |
| 5 | Caspase 8 | 5′-CTG GTC TGA AGG CTG GTT GT-3′ | 5′-CAG GCT CAG GAA CTT GAG GG-3′ |
| 6 | Caspase 3 | 5′-CTG GTT TTC GGT GGG TGT G-3′ | 5′-ACG GCA GGC CTG AAT AAT GAA |
| 7 | GAPDH | 5′-GGT ATC GTG GAA GGA CTC ATG AC-3′ | 5′-ATG CCA GTG AGC TTC CCG TTC AGC |
| Sample | Cell Lines and IC50 (µg/mL) | |||
|---|---|---|---|---|
| A549 | MDA-MB-231 | LoVo | MCF-7 | |
| D. innoxia | 47.76 ± 1.78 | 32.61 ± 1.29 | 10.1 ± 0.55 | 23.97 ± 0.66 |
| Doxorubicin | 1.2 ± 0.06 | 1.3 ± 0.08 | 1.1 ± 0.8 | 1.5 ± 0. 6 |
| Compound Name | Chemical Formula | Molecular Weight (g/mol) | RT (min) | Area% |
|---|---|---|---|---|
| Bicyclo [3.2.0]heptane | C7H12 | 96.17 | 5.41 | 0.26 |
| 1-Propene-1-thiol | C3H6S | 74.15 | 5.525 | 0.73 |
| 3-Methyl-isoxazol-5(4H)-one | C4H5NO2 | 99.09 | 6.027 | 0.32 |
| (Z)-3-Methyl-2-hexene | C7H14 | 98.19 | 6.67 | 0.31 |
| Alpha.Aminooxy-propionic acid | C3H7NO3 | 105.09 | 6.861 | 0.35 |
| 2,3-Dimethyl-3-pentanol | C7H16O | 116.2 | 7.338 | 12.89 |
| 3,4,6-Tri-O-methyl-d-glucose | C9H18O6 | 222.24 | 8.693 | 0.51 |
| 3,3-Dimethyl-2-pentanol | C7H16O | 116.2 | 8.814 | 2.09 |
| 2,6-diethyl-Benzenamine | C10H15N | 149.23 | 9.393 | 0.32 |
| 3-(1-Cyclopentenyl) furan | C9H10O | 134.17 | 9.475 | 0.44 |
| N,N-dimethyl-Propanamide | C5H11NO | 101.15 | 9.933 | 1.03 |
| 4-methyl-1,3-Dioxane | C5H10O2 | 102.13 | 10.493 | 2.2 |
| 3-methyl-Benzoyl chloride | C8H7ClO | 154.59 | 11.212 | 0.65 |
| 2-hydroxy-4-methyl Pentanoic acid | C6H12O3 | 132.16 | 12.109 | 5.19 |
| Phenoxyacetamide | C8H9NO2 | 151.16 | 12.714 | 3.65 |
| 1-Heptynylbenzene | C13H16 | 172.27 | 13.42 | 0.5 |
| Palmitoleic acid | C16H30O2 | 254.41 | 15.971 | 0.3 |
| 3-(2-furanyl)-2-Propenal | C7H6O2 | 122.12 | 16.098 | 0.36 |
| Megastigmatrienone | C13H18O | 190.28 | 16.321 | 0.46 |
| 2-nitrophenyl azide | C6H4N4O2 | 164.12 | 16.881 | 0.5 |
| Spiro [2.3] hexan-4-one, 5,5-diethyl | C10H16O | 152.23 | 17.224 | 0.33 |
| 3-Nonyn-1-ol | C9H16O | 140.22 | 17.6 | 0.29 |
| Methyl 3-methoxy-4-nitrobenzoate | C9H9NO5 | 211.17 | 18.166 | 0.61 |
| Cyclopropaneoctanal | C11H20O | 168.28 | 18.242 | 0.6 |
| (6R)-6alpha-[(Z)-1,3-Butadienyl]-1,4-cycloheptadiene | C11H14 | 146.23 | 18.541 | 0.39 |
| 2,3,3,4,5-pentaethyl 1,2,5-Oxadiborolane | C12H26O2O | 207.96 | 19.082 | 1.26 |
| Cyclotridecane | C13H26 | 182.35 | 19.158 | 0.6 |
| Dolichodial | C10H14O2 | 166.21 | 19.527 | 0.44 |
| p-Menth-8(10)-en-9-ol | C10H18O | 154.25 | 19.96 | 0.61 |
| 1,4-Icosanediol | C20H42O2 | 314.5 | 20.628 | 0.95 |
| 13-Tetradece-11-yn-1-ol | C14H24O | 208.34 | 20.736 | 0.26 |
| 9,17-Octadecadienal | C18H32O | 264.4 | 21.137 | 0.29 |
| 1,19-Eicosadiene | C20H38 | 278.5 | 21.678 | 1.35 |
| Heneicosyl formate | C22H44O2 | 340.6 | 21.792 | 14.14 |
| 8-Tetradecen-1-ol | C14H28O | 212.37 | 22.053 | 4.21 |
| 3-Methoxybenzylamine | C8H11NO | 137.18 | 22.282 | 0.7 |
| Linoleic acid | C18H32O2 | 280.4 | 22.352 | 0.86 |
| Bicyclo [6.1.0] non-1-ene | C9H14 | 122.21 | 22.473 | 0.33 |
| 4-Isopropenylcyclohexanone | C9H14O | 138.21 | 22.708 | 2.89 |
| Cyclohexane, 1,1′-methylenebis | C13H24 | 180.33 | 23.045 | 2.55 |
| Z, Z-3,13-Octadecadien-1-ol acetate | C20H36O2 | 308.5 | 23.548 | 0.6 |
| 9-Methyl-Z, Z-10,12-hexadecadien-1-ol acetate | C19H34O2 | 294.5 | 23.618 | 0.54 |
| 2,10-dimethyl-9-Undecenal | C13H24O | 196.33 | 23.917 | 0.35 |
| 3,5-Dihydroxybenzoic acid | C7H6O4 | 154.12 | 24.184 | 16.53 |
| 7-Pentadecyne | C15H28 | 208.38 | 24.954 | 0.25 |
| 1,3,12-Nonadecatriene | C19H34 | 262.5 | 25.139 | 0.49 |
| 1,5,9-Cyclododecatriene | C12H18 | 162.27 | 26.532 | 0.79 |
| Monoelaidin | C21H40O4 | 356.5 | 27.41 | 1.23 |
| 2-methyl-5-(1-methyl ethenyl)-Cyclohexanol | C10H18O | 154.25 | 27.76 | 1.21 |
| Z-(13,14-Epoxy) tetradec-11-en-1-ol acetate | C16H28O3 | 268.39 | 27.868 | 1.24 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al-Zharani, M.; Nasr, F.A.; Alqahtani, A.S.; Cordero, M.A.W.; Alotaibi, A.A.; Bepari, A.; Alarifi, S.; Daoud, A.; Barnawi, I.O.; Daradka, H.M. In Vitro Cytotoxic Evaluation and Apoptotic Effects of Datura innoxia Grown in Saudi Arabia and Phytochemical Analysis. Appl. Sci. 2021, 11, 2864. https://doi.org/10.3390/app11062864
Al-Zharani M, Nasr FA, Alqahtani AS, Cordero MAW, Alotaibi AA, Bepari A, Alarifi S, Daoud A, Barnawi IO, Daradka HM. In Vitro Cytotoxic Evaluation and Apoptotic Effects of Datura innoxia Grown in Saudi Arabia and Phytochemical Analysis. Applied Sciences. 2021; 11(6):2864. https://doi.org/10.3390/app11062864
Chicago/Turabian StyleAl-Zharani, Mohammed, Fahd A. Nasr, Ali S. Alqahtani, Mary Anne W. Cordero, Amal A. Alotaibi, Asmatanzeem Bepari, Saud Alarifi, Ali Daoud, Ibrahim O. Barnawi, and Haytham M. Daradka. 2021. "In Vitro Cytotoxic Evaluation and Apoptotic Effects of Datura innoxia Grown in Saudi Arabia and Phytochemical Analysis" Applied Sciences 11, no. 6: 2864. https://doi.org/10.3390/app11062864