

Micropaleontological and Isotopic Perspective of Surface Water Mass Variability in the NE Atlantic from MIS 6 to 5e (188 to 115 ka)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
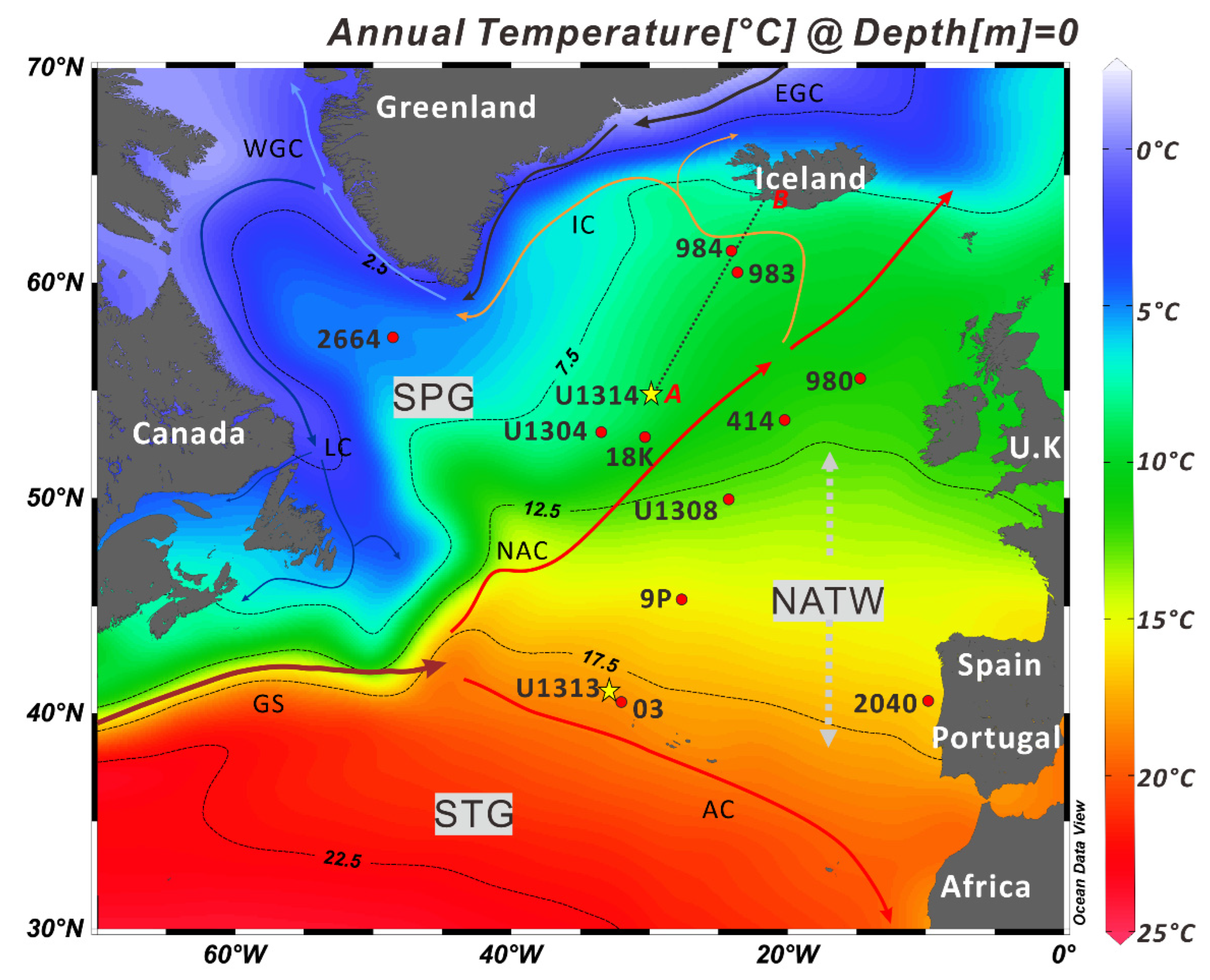
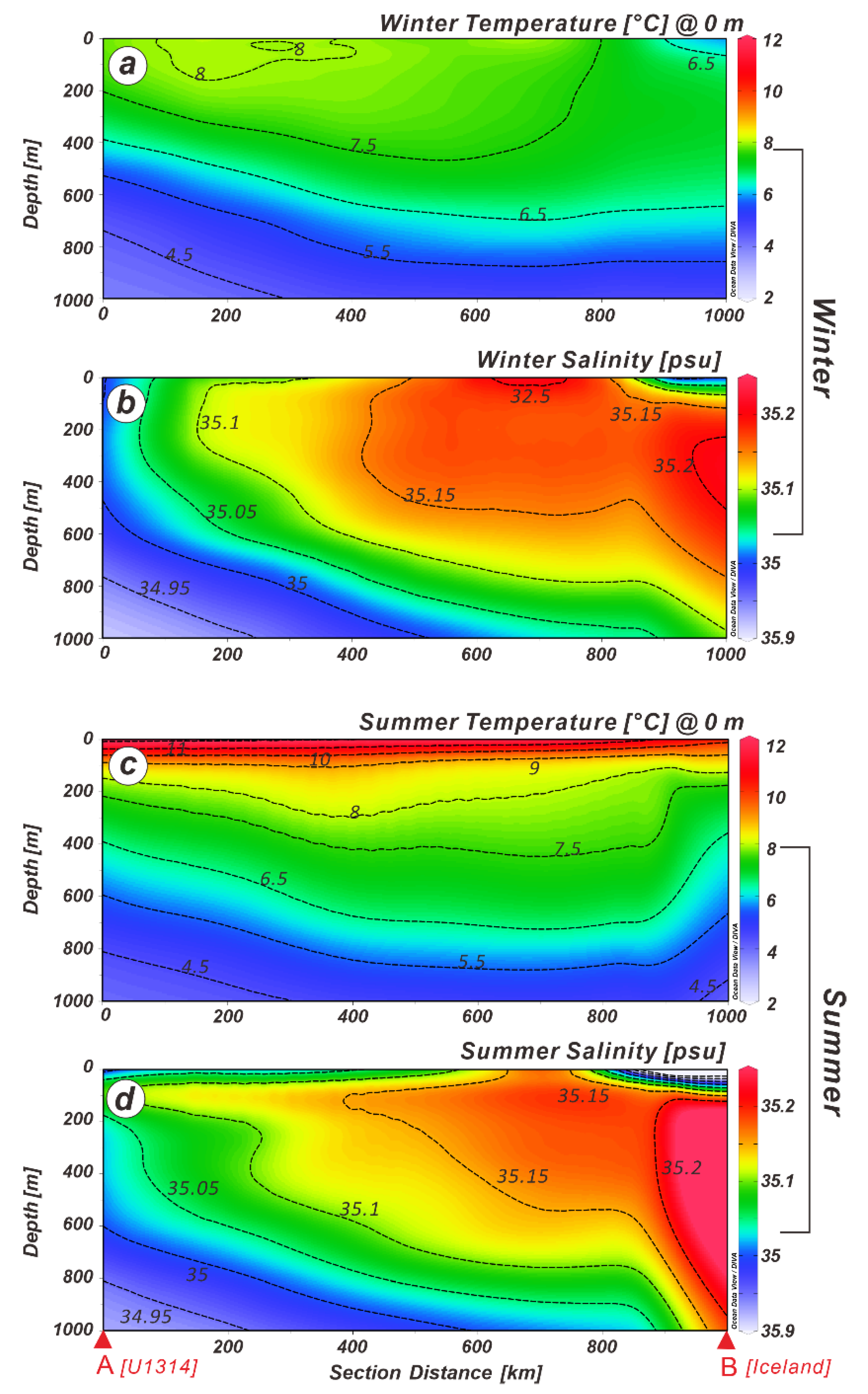
2. Geological and Oceanographic Setting
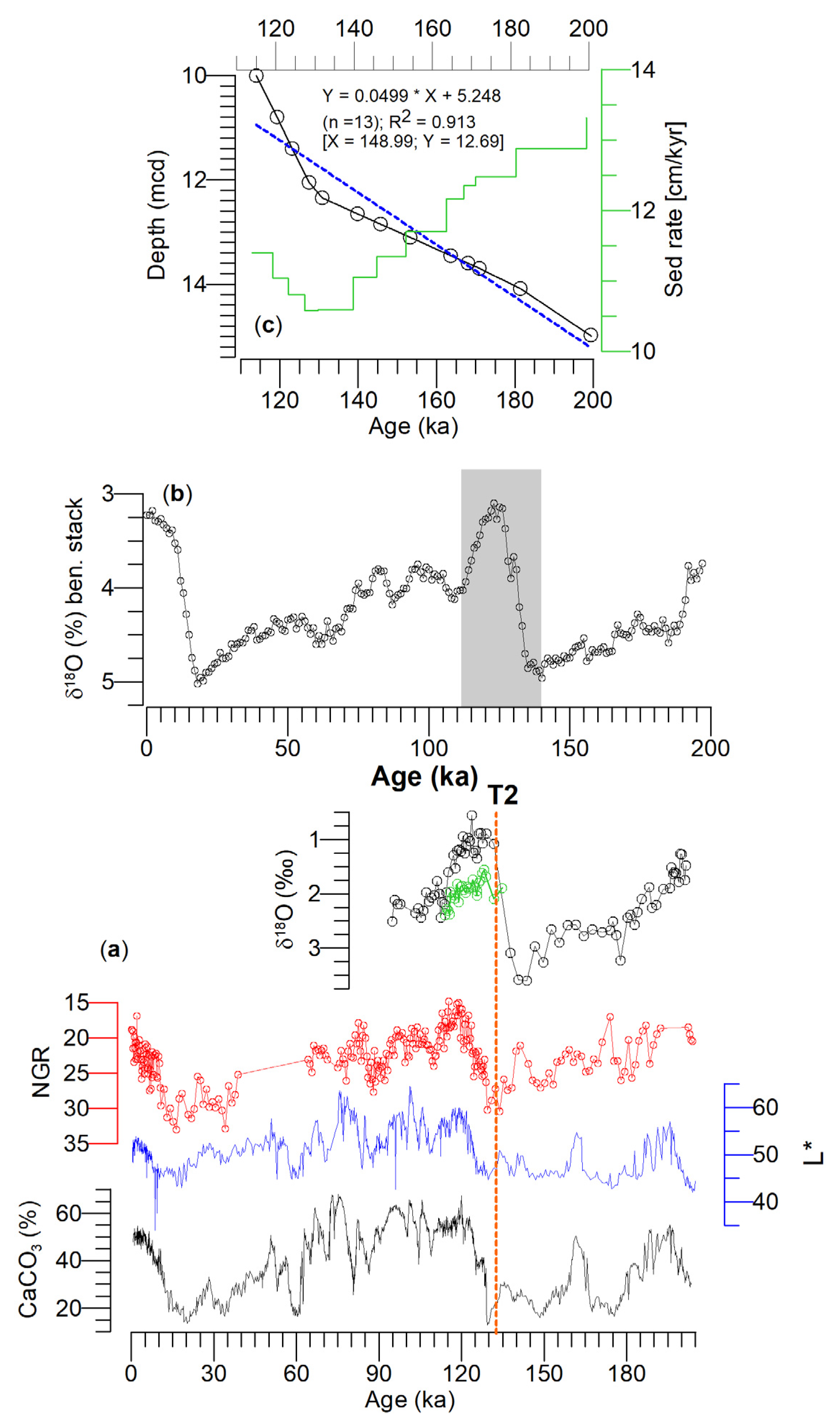
3. Materials and Methods
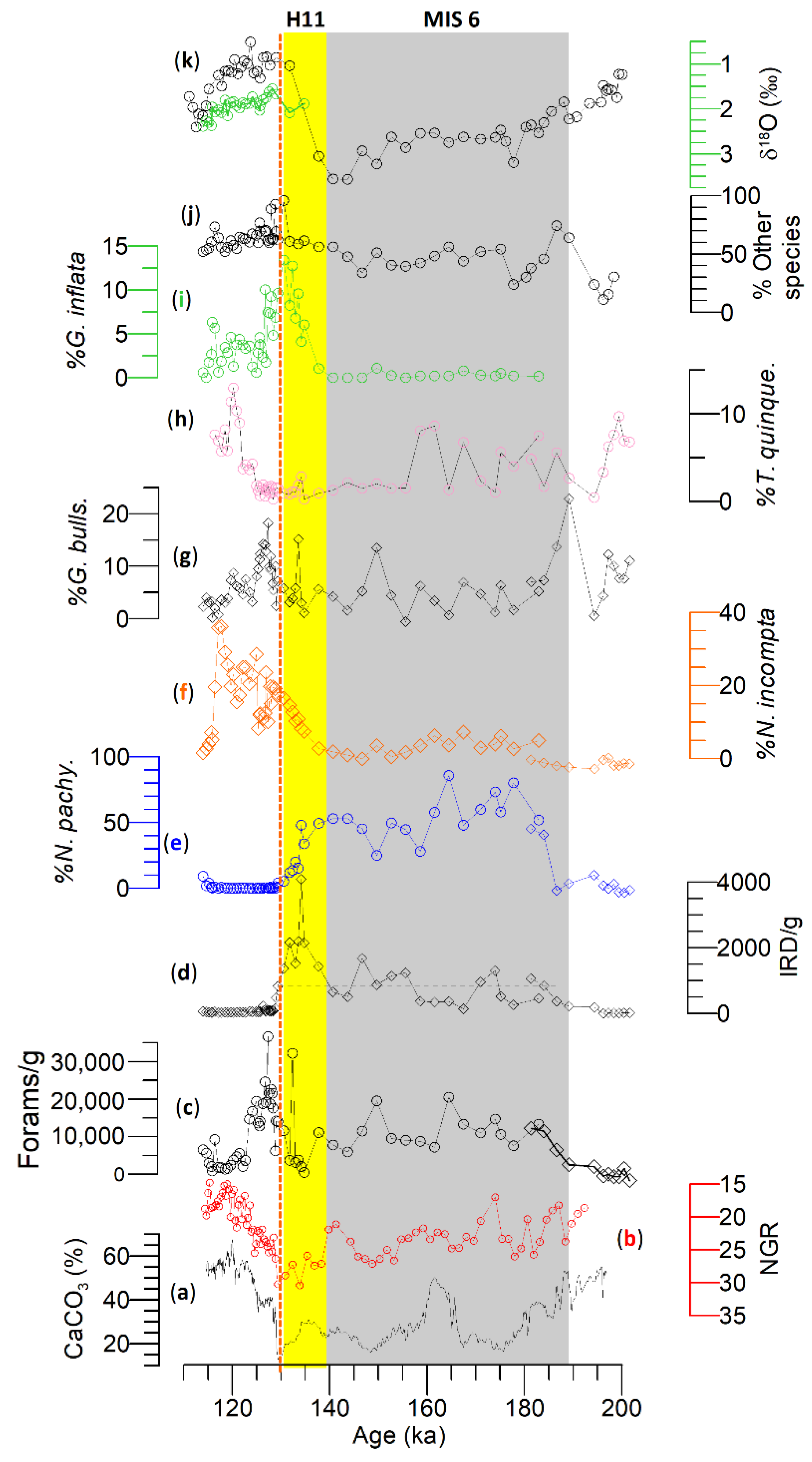
4. Results
- Site U1314
- Site U1313
5. Discussion
5.1. Subpolar to Subtropical Water Mass Movement during MIS 6
5.2. Subpolar-Subtropical Surface Water Masses Transformation during T2 and MIS 5e
6. Conclusions
- (i).
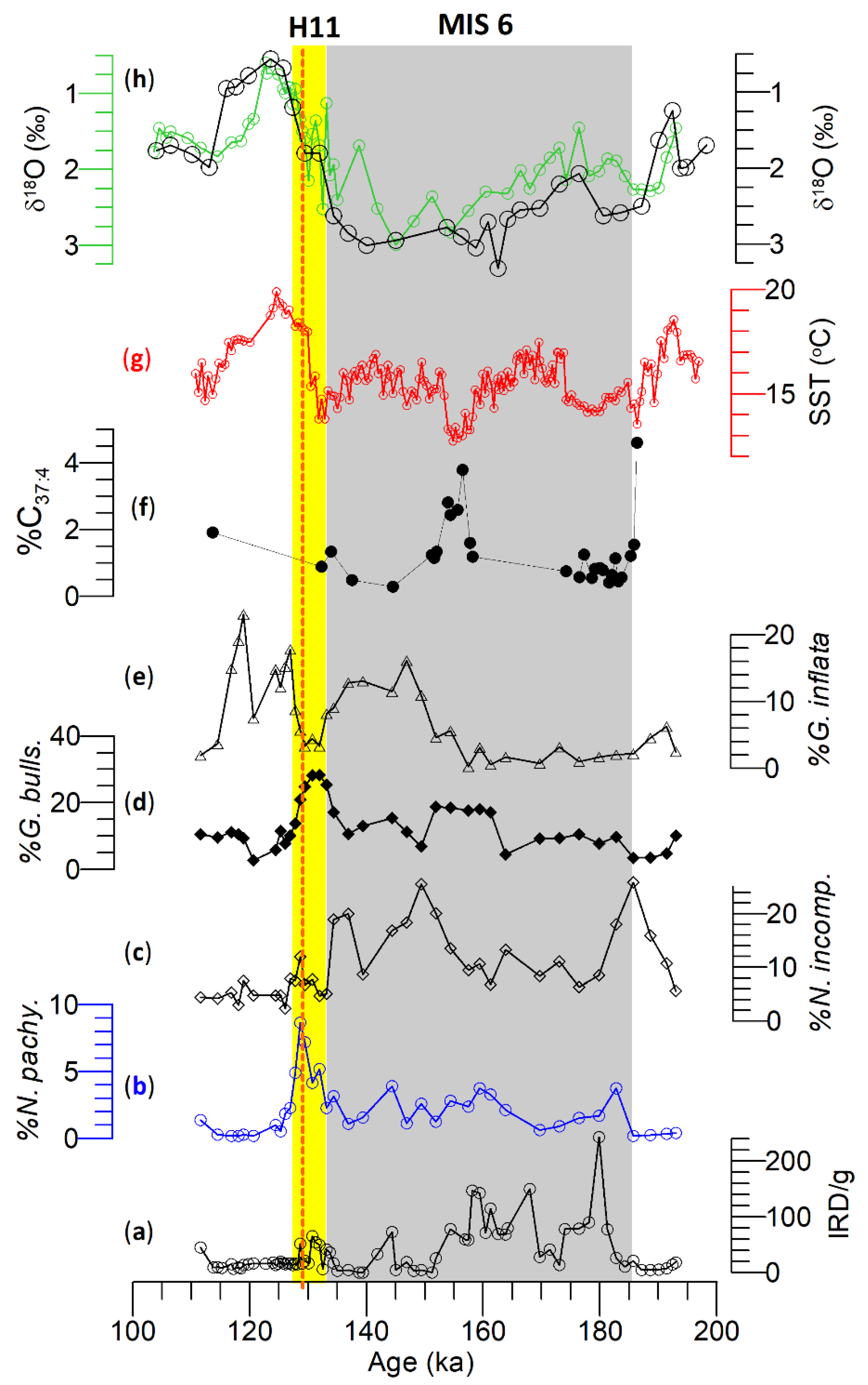
- During MIS 6, four low amplitude IRD peaks were identified where the IRD peaks at Site U1314 were higher than that of Site U1313. N. pachyderma is dominant at Site U1314, reflecting the prevailing presence of polar and subpolar water masses. The diversity of planktonic foraminifers at the STG Site U1313 is higher than that of subpolar Site U1314 in addition to subpolar and transitional species alongside significant changes in N. pachyderma. These changes in the foraminiferal assemblages reflect the evolution of the North Atlantic transitional water and subpolar water masses.
- (ii).
- During MIS 6, the δ18O of G. bulloides and G. inflata is nearly identical at Site U1313, most likely caused by the seasonal incursion of transitional water masses resulting in seasonal blooms of each species dwelling in their respective depth habitats. However, this proposition requires examination by isolating temperature from the δ18O in both species.
- (iii).
- Planktonic foraminifera δ18O values at sites U1313 and U1314 during T2 reflect regional heterogeneity of transition from penultimate deglacial to interglacial time. It appears that the transition speed at Site U1314 was fast compared to Site U1313. Further, the identical δ18O values of G. bulloides and G. inflata at Site U1313 reflect a well-mixed water column during T2.
- (iv).
- Negative △δ18O (δ18OGb–δ18OGinf.) values between sites U1313 and U1314 reflect the re-establishment of the water column (i.e., mixed layer and thermocline) during MIS 5e. The N. incompta increase at Site U1314, and the increase in G. inflata at Site U1313 reflects the influence of warm NAC during MIS 5e.
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Svendsen, J.I.; Alexanderson, H.; Astakhov, V.I.; Demidov, I.; Dowdeswell, J.A.; Funder, S.; Gataullin, V.; Henriksen, M.; Hjort, C.; Houmark-Nielsen, M.; et al. Late Quaternary Ice Sheet History of Northern Eurasia. Quat. Sci. Rev. 2004, 23, 1229–1271. [Google Scholar] [CrossRef]
- Jakobsson, M.; Nilsson, J.; O’Regan, M.; Backman, J.; Löwemark, L.; Dowdeswell, J.A.; Mayer, L.; Polyak, L.; Colleoni, F.; Anderson, L.G.; et al. An Arctic Ocean Ice Shelf during MIS 6 Constrained by New Geophysical and Geological Data. Quat. Sci. Rev. 2010, 29, 3505–3517. [Google Scholar] [CrossRef]
- McManus, J.F.; Oppo, D.W.; Cullen, J.L. A 0.5-Million-Year Record of Millennial-Scale Climate Variability in the North Atlantic. Science 1999, 283, 971–975. [Google Scholar] [CrossRef]
- Mokeddem, Z.; McManus, J.F. Persistent Climatic and Oceanographic Oscillations in the Subpolar North Atlantic during the MIS 6 Glaciation and MIS 5 Interglacial. Paleoceanography 2016, 31, 758–778. [Google Scholar] [CrossRef]
- Bond, G.; Broecker, W.; Johnsen, S.; McManus, J.; Labeyrie, L.; Jouzel, J.; Bonani, G. Correlations between Climate Records from North Atlantic Sediments and Greenland Ice. Nature 1993, 365, 143–147. [Google Scholar] [CrossRef]
- McManus, J.F.; Bond, G.C.; Broecker, W.S.; Johnsen, S.; Labeyrie, L.; Higgins, S. High-Resolution Climate Records from the North Atlantic during the Last Interglacial. Nature 1994, 371, 326–329. [Google Scholar] [CrossRef]
- Kandiano, E.S.; Bauch, H.A. Surface Ocean Temperatures in the North-East Atlantic during the Last 500 000 Years: Evidence from Foraminiferal Census Data: Surface Ocean Temperatures in the North-East Atlantic. Terra Nova 2003, 15, 265–271. [Google Scholar] [CrossRef]
- Ruddiman, W.F.; Raymo, M.E.; Martinson, D.G.; Clement, B.M.; Backman, J. Pleistocene Evolution: Northern Hemisphere Ice Sheets and North Atlantic Ocean. Paleoceanography 1989, 4, 353–412. [Google Scholar] [CrossRef]
- Chapman, M.R.; Shackleton, N.J.; Duplessy, J.-C. Sea Surface Temperature Variability during the Last Glacial–Interglacial Cycle: Assessing the Magnitude and Pattern of Climate Change in the North Atlantic. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2000, 157, 1–25. [Google Scholar] [CrossRef]
- Naafs, B.D.A.; Hefter, J.; Stein, R. Millennial-Scale Ice Rafting Events and Hudson Strait Heinrich(-like) Events during the Late Pliocene and Pleistocene: A Review. Quat. Sci. Rev. 2013, 80, 1–28. [Google Scholar] [CrossRef]
- Smith, M.; Glick, E.; Lodestro, S.; Rashid, H. Data Report: Oxygen Isotopes and Foraminifer Abundance Record for the Last Glacial–Interglacial Cycle and Marine Isotope Stage 6 at IODP Site U1313. In Expedition 303/306 Scientists; Channell, J.E.T., Kanamatsu, T., Sato, T., Stein, R., Alvarez Zarikian, C.A., Malone, M.J., Eds.; Integrated Ocean Drilling Program Management International, Inc.: College Station, TX, USA, 2013; Volume 303/306. [Google Scholar]
- Cheng, H.; Edwards, R.L.; Broecker, W.S.; Denton, G.H.; Kong, X.; Wang, Y.; Zhang, R.; Wang, X. Ice Age Terminations. Science 2009, 326, 248–252. [Google Scholar] [CrossRef] [PubMed]
- Ruddiman, W.F.; McIntyre, A. Oceanic Mechanisms for Amplification of the 23,000-Year Ice-Volume Cycle. Science 1981, 212, 617–627. [Google Scholar] [CrossRef] [PubMed]
- Sarnthein, M.; Winn, K.; Jung, S.J.A.; Duplessy, J.-C.; Labeyrie, L.; Erlenkeuser, H.; Ganssen, G. Changes in East Atlantic Deepwater Circulation over the Last 30,000 Years: Eight Time Slice Reconstructions. Paleoceanography 1994, 9, 209–267. [Google Scholar] [CrossRef]
- Rashid, H.; Boyle, E.A. Mixed-Layer Deepening During Heinrich Events: A Multi-Planktonic Foraminiferal δ 18 O Approach. Science 2007, 318, 439–441. [Google Scholar] [CrossRef]
- Cacho, I. Chapter 2—Quaternary Ice Ages in the Iberian Peninsula. In Iberia, Land of Glaciers; Oliva, M., Palacios, D., Fernández-Fernández, J.M., Eds.; Elsevier: Amsterdam, The Netherlands, 2022; pp. 13–35. ISBN 978-0-12-821941-6. [Google Scholar]
- Bauch, H.A.; Erlenkeuser, H.; Grootes, P.M.; Jouzel, J. Implications of Stratigraphic and Paleoclimatic Records of the Last Interglaciation from the Nordic Seas. Quat. Res. 1996, 46, 260–269. [Google Scholar] [CrossRef]
- Oppo, D.W.; Horowitz, M.; Lehman, S.J. Marine Core Evidence for Reduced Deep Water Production during Termination II Followed by a Relatively Stable Substage 5e (Eemian). Paleoceanography 1997, 12, 51–63. [Google Scholar] [CrossRef]
- Mokeddem, Z.; McManus, J.F.; Oppo, D.W. Oceanographic Dynamics and the End of the Last Interglacial in the Subpolar North Atlantic. Proc. Natl. Acad. Sci. USA 2014, 111, 11263–11268. [Google Scholar] [CrossRef]
- Govin, A.; Capron, E.; Tzedakis, P.C.; Verheyden, S.; Ghaleb, B.; Hillaire-Marcel, C.; St-Onge, G.; Stoner, J.S.; Bassinot, F.; Bazin, L.; et al. Sequence of Events from the Onset to the Demise of the Last Interglacial: Evaluating Strengths and Limitations of Chronologies Used in Climatic Archives. Quat. Sci. Rev. 2015, 129, 1–36. [Google Scholar] [CrossRef]
- Deaney, E.L.; Barker, S.; van de Flierdt, T. Timing and Nature of AMOC Recovery across Termination 2 and Magnitude of Deglacial CO2 Change. Nat. Commun. 2017, 8, 14595. [Google Scholar] [CrossRef]
- Irvali, N.; Ninnemann, U.S.; Galaasen, E.V.; Rosenthal, Y.; Kroon, D.; Oppo, D.W.; Kleiven, H.F.; Darling, K.F.; Kissel, C. Rapid switches in subpolar North Atlantic hydrography and climate during the Lat Interglacial (MIS 5e). Paleoceanography 2012, 27, PA2207. [Google Scholar] [CrossRef]
- CLIMAP Project Members. The Last Interglacial Ocean. Quat. Res. 1984, 21, 123–224. [Google Scholar] [CrossRef]
- Adkins, J.F.; Boyle, E.A.; Keigwin, L.; Cortijo, E. Variability of the North Atlantic Thermohaline Circulation during the Last Interglacial Period. Nature 1997, 390, 154–156. [Google Scholar] [CrossRef]
- Hodell, D.A.; Minth, E.K.; Curtis, J.H.; McCave, I.N.; Hall, I.R.; Channell, J.E.T.; Xuan, C. Surface and Deep-Water Hydrography on Gardar Drift (Iceland Basin) during the Last Interglacial Period. Earth Planet. Sci. Lett. 2009, 288, 10–19. [Google Scholar] [CrossRef]
- Kukla, G.J.; Bender, M.L.; de Beaulieu, J.-L.; Bond, G.; Broecker, W.S.; Cleveringa, P.; Gavin, J.E.; Herbert, T.D.; Imbrie, J.; Jouzel, J.; et al. Last Interglacial Climates. Quat. Res. 2002, 58, 2–13. [Google Scholar] [CrossRef]
- Shackleton, N.J.; Sánchez-Goñi, M.F.; Pailler, D.; Lancelot, Y. Marine Isotope Substage 5e and the Eemian Interglacial. Glob. Planet. Chang. 2003, 36, 151–155. [Google Scholar] [CrossRef]
- Lototskaya, A.; Ganssen, G.M. The Structure of Termination II (Penultimate Deglaciation and Eemian) in the North Atlantic. Quat. Sci. Rev. 1999, 18, 1641–1654. [Google Scholar] [CrossRef]
- Tzedakis, P.C.; Drysdale, R.N.; Margari, V.; Skinner, L.C.; Menviel, L.; Rhodes, R.H.; Taschetto, A.S.; Hodell, D.A.; Crowhurst, S.J.; Hellstrom, J.C.; et al. Enhanced Climate Instability in the North Atlantic and Southern Europe during the Last Interglacial. Nat. Commun. 2018, 9, 4235. [Google Scholar] [CrossRef]
- Clark, P.U.; He, F.; Golledge, N.R.; Mitrovica, J.X.; Dutton, A.; Hoffman, J.S.; Dendy, S. Oceanic Forcing of Penultimate Deglacial and Last Interglacial Sea-Level Rise. Nature 2020, 577, 660–664. [Google Scholar] [CrossRef]
- Stoll, H.M.; Cacho, I.; Gasson, E.; Sliwinski, J.; Kost, O.; Moreno, A.; Iglesias, M.; Torner, J.; Perez-Mejias, C.; Haghipour, N.; et al. Rapid Northern Hemisphere Ice Sheet Melting during the Penultimate Deglaciation. Nat. Commun. 2022, 13, 3819. [Google Scholar] [CrossRef]
- Brummer, G.-J.A.; Metcalfe, B.; Feldmeijer, W.; Prins, M.A.; Van ’T Hoff, J.; Ganssen, G.M. Modal Shift in North Atlantic Seasonality during the Last Deglaciation. Clim. Past 2020, 16, 265–282. [Google Scholar] [CrossRef]
- Channell, J.E.T.; Kanamatsu, T.; Sato, T.; Stein, R.; Alvarez Zarikian, C.A.; Malone, M.J. (Eds.) Expedition 306 Scientists Expedition 306 Summary; Integrated Ocean Drilling Program: College Station, TX, USA, 2006; Volume 303/306. [Google Scholar]
- Wold, C.N. Cenozoic sediment accumulation on Drifts in the northern North Atlantic. Paleoceanography 1994, 9, 917–941. [Google Scholar] [CrossRef]
- Grützner, J.; Higgins, S.M. Threshold Behavior of Millennial Scale Variability in Deep Water Hydrography Inferred from a 1.1 Ma Long Record of Sediment Provenance at the Southern Gardar Drift. Paleoceanography 2010, 25, PA4204. [Google Scholar] [CrossRef]
- Hansen, B.; Østerhus, S. North Atlantic–Nordic Seas Exchanges. Prog. Oceanogr. 2000, 45, 109–208. [Google Scholar] [CrossRef]
- Lozier, M.S.; Li, F.; Bacon, S.; Bahr, F.; Bower, A.S.; Cunningham, S.A.; de Jong, M.F.; de Steur, L.; deYoung, B.; Fischer, J. A Sea Change in Our View of Overturning in the Subpolar North Atlantic. Science 2019, 363, 516–521. [Google Scholar] [CrossRef]
- Locarnini, R.A.; Mishonov, A.V.; Baranova, O.K.; Boyer, T.P.; Zweng, M.M.; Garcia, H.E.; Reagan, J.R.; Seidov, D.; Weathers, K.W.; Paver, C.R.; et al. World Ocean Atlas 2018 Volume 1: Temperature; NOAA Atlas NESDIS 81; U.S. Department of Commerce: Silver Spring, MD, USA, 2019; 52p.
- Petit, T.; Mercier, H.; Thierry, V. New Insight Into the Formation and Evolution of the East Reykjanes Ridge Current and Irminger Current. J. Geophys. Res. Oceans 2019, 124, 9171–9189. [Google Scholar] [CrossRef]
- Schlitzer, R. Ocean Data View; Alfred Wegner Institute: Bremerhaven, Germany, 2018. [Google Scholar]
- Alonso-Garcia, M.; Sierro, F.J.; Kucera, M.; Flores, J.A.; Cacho, I.; Andersen, N. Ocean Circulation, Ice Sheet Growth and Interhemispheric Coupling of Millennial Climate Variability during the Mid-Pleistocene (ca 800–400 Ka). Quat. Sci. Rev. 2011, 30, 3234–3247. [Google Scholar] [CrossRef]
- Rashid, H.; Lu, Q.Q.; Zeng, M.; Wang, Y.; Zhang, Z.W. Sea-Surface Characteristics of the Newfoundland Basin of the Northwest Atlantic Ocean during the Last 145,000 Years: A Study Based on the Sedimentological and Paleontological Proxies. Appl. Sci. 2021, 11, 3343. [Google Scholar] [CrossRef]
- Bond, G.C.; Lotti, R. Iceberg Discharges into the North Atlantic on Millennial Time Scales during the Last Glaciation. Science 1995, 267, 1005–1010. [Google Scholar] [CrossRef]
- Rashid, H.; Saint-Ange, F.; Barber, D.C.; Smith, M.E.; Devalia, N. Fine Scale Sediment Structure and Geochemical Signature between Eastern and Western North Atlantic during Heinrich Events 1 and 2. Quat. Sci. Rev. 2012, 46, 136–150. [Google Scholar] [CrossRef]
- Alonso-Garcia, M.; Sierro, F.J.; Flores, J.A. Arctic Front Shifts in the Subpolar North Atlantic during the Mid-Pleistocene (800–400 Ka) and Their Implications for Ocean Circulation. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2011, 311, 268–280. [Google Scholar] [CrossRef]
- Lisiecki, L.E.; Raymo, M.E. A Pliocene-Pleistocene Stack of 57 Globally Distributed Benthic δ 18 O Records. Paleoceanography 2005, 20, PA1003. [Google Scholar] [CrossRef]
- Alvarez Zarikian, C.A.; Stepanova, A.Y.; Grutzner, J. Glacial–Interglacial Variability in Deep Sea Ostracod Assemblage Composition at IODP Site U1314 in the Subpolar North Atlantic. Mar. Geol. 2009, 258, 69–87. [Google Scholar] [CrossRef]
- Obrochta, S.P.; Crowley, T.J.; Channell, J.E.T.; Hodell, D.A.; Baker, P.A.; Seki, A.; Yokoyama, Y. Climate Variability and Ice-Sheet Dynamics during the Last Three Glaciations. Earth Planet. Sci. Lett. 2014, 406, 198–212. [Google Scholar] [CrossRef]
- Barker, S.; Chen, J.; Gong, X.; Jonkers, L.; Knorr, G.; Thornalley, D. Icebergs Not the Trigger for North Atlantic Cold Events. Nature 2015, 520, 333–336. [Google Scholar] [CrossRef]
- De Abreu, L.; Shackleton, N.J.; Schönfeld, J.; Hall, M.; Chapman, M. Millennial-Scale Oceanic Climate Variability off the Western Iberian Margin during the Last Two Glacial Periods. Mar. Geol. 2003, 196, 1–20. [Google Scholar] [CrossRef]
- Toucanne, S.; Zaragosi, S.; Bourillet, J.F.; Gibbard, P.L.; Eynaud, F.; Giraudeau, J.; Turon, J.L.; Cremer, M.; Cortijo, E.; Martinez, P.; et al. A 1.2Ma Record of Glaciation and Fluvial Discharge from the West European Atlantic Margin. Quat. Sci. Rev. 2009, 28, 2974–2981. [Google Scholar] [CrossRef]
- Heinrich, H. Origin and Consequences of Cyclic Ice Rafting in the Northeast Atlantic Ocean During the Past 130,000 Years. Quat. Res. 1988, 29, 142–152. [Google Scholar] [CrossRef]
- Grousset, F.E.; Pujol, C.; Labeyrie, L.; Auffret, G.; Boelaert, A. Were the North Atlantic Heinrich Events Triggered by the Behavior of the European Ice Sheets? Geology 2000, 28, 123. [Google Scholar] [CrossRef]
- Channell, J.E.T.; Hodell, D.A.; Romero, O.; Hillaire-Marcel, C.; de Vernal, A.; Stoner, J.S.; Mazaud, A.; Röhl, U. A 750-Kyr Detrital-Layer Stratigraphy for the North Atlantic (IODP Sites U1302–U1303, Orphan Knoll, Labrador Sea). Earth Planet. Sci. Lett. 2012, 317–318, 218–230. [Google Scholar] [CrossRef]
- Piper, D.J.W.; Tripsanas, E.; Mosher, D.C.; MacKillop, K. Paleoseismicity of the Continental Margin of Eastern Canada: Rare Regional Failures and Associated Turbidites in Orphan Basin. Geosphere 2019, 15, 85–107. [Google Scholar] [CrossRef]
- Bard, E.; Rostek, F.; Turon, J.-L.; Gendreau, S. Hydrological Impact of Heinrich Events in the Subtropical Northeast Atlantic. Science 2000, 289, 1321–1324. [Google Scholar] [CrossRef] [PubMed]
- Hodell, D.A.; Channell, J.E.T.; Curtis, J.H.; Romero, O.E.; Röhl, U. Onset of “Hudson Strait” Heinrich Events in the Eastern North Atlantic at the End of the Middle Pleistocene Transition (∼640 Ka)? Paleoceanogr. Paleoclimatol. 2008, 23, PA4218. [Google Scholar] [CrossRef]
- Margari, V.; Skinner, L.C.; Tzedakis, P.C.; Ganopolski, A.; Vautravers, M.; Shackleton, N.J. The Nature of Millennial-Scale Climate Variability during the Past Two Glacial Periods. Nat. Geosci. 2010, 3, 127–131. [Google Scholar] [CrossRef]
- Johannessen, T.; Jansen, E.; Flatøy, A.; Ravelo, A.C. The Relationship between Surface Water Masses, Oceanographic Fronts and Paleoclimatic Proxies in Surface Sediments of the Greenland, Iceland, Norwegian Seas; Zahn, R., Pedersen, T.F., Kaminski, M.A., Labeyrie, L., Eds.; Springer: Berlin/Heidelberg, Germany, 1994; pp. 61–85. [Google Scholar]
- Stangeew, E. Distribution and Isotopic Composition of Living Planktonic Foraminifera N. pachyderma (Sinistral) and T. quinqueloba in the High Latitude North Atlantic. Ph.D. Thesis, University of Kiel, Kiel, Germany, 2001. [Google Scholar]
- Barrenechea Angeles, I.; Lejzerowicz, F.; Cordier, T.; Scheplitz, J.; Kucera, M.; Ariztegui, D.; Pawlowski, J.; Morard, R. Planktonic Foraminifera EDNA Signature Deposited on the Seafloor Remains Preserved after Burial in Marine Sediments. Sci. Rep. 2020, 10, 20351. [Google Scholar] [CrossRef]
- Sahoo, N.; Saalim, S.M.; Matul, A.; Mohan, R.; Tikhonova, A.; Kozina, N. Planktic Foraminiferal Assemblages in Surface Sediments From the Subpolar North Atlantic Ocean. Front. Mar. Sci. 2022, 8, 781675. [Google Scholar] [CrossRef]
- Kandiano, E.S.; Bauch, H.A.; Müller, A. Sea Surface Temperature Variability in the North Atlantic during the Last Two Glacial–Interglacial Cycles: Comparison of Faunal, Oxygen Isotopic, and Mg/Ca-Derived Records. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2004, 204, 145–164. [Google Scholar] [CrossRef]
- Chapman, M.R. Seasonal Production Patterns of Planktonic Foraminifera in the NE Atlantic Ocean: Implications for Paleotemperature and Hydrographic Reconstructions: CURRENTS. Paleoceanography 2010, 25, PA1101. [Google Scholar] [CrossRef]
- Farmer, E.J.; Chapman, M.R.; Andrews, J.E. Holocene Temperature Evolution of the Subpolar North Atlantic Recorded in the Mg/Ca Ratios of Surface and Thermocline Dwelling Planktonic Foraminifers. Glob. Planet. Chang. 2011, 79, 234–243. [Google Scholar] [CrossRef]
- Billups, K.; Vizcaíno, M.; Chiarello, J.; Kaiser, E.A. Reconstructing Western Boundary Current Stability in the North Atlantic Ocean for the Past 700 Kyr From Globorotalia truncatulinoides Coiling Ratios. Paleoceanogr. Paleoclimatol. 2020, 35, e2020PA003958. [Google Scholar] [CrossRef]
- Bé, A.; Tolderlund, O.S. Distribution and Ecology of Living Planktonic Foraminifera in Surface Waters of the Atlantic and Indian Oceans. In The Micropaleontology of Oceans; Cambridge University Press: New York, NY, USA, 1971; pp. 105–149. [Google Scholar]
- Duplessy, J.-C.; Labeyrie, L.; Juilletleclerc, A.; Maitre, F.; Duprat, J.; Sarnthein, M. Surface Salinity Reconstruction of the North Atlantic Ocean during the Last Glacial Maximum. Oceanol. Acta 1991, 14, 311–324. [Google Scholar]
- Elderfield, H.; Ganssen, G. Past Temperature and δ18O of Surface Ocean Waters Inferred from Foraminiferal Mg/Ca Ratios. Nature 2000, 405, 442–445. [Google Scholar] [CrossRef] [PubMed]
- Schiebel, R.; Bijma, J.; Hemleben, C. Population Dynamics of the Planktic Foraminifer Globigerina Bulloides from the Eastern North Atlantic. Deep. Sea Res. Part I Oceanogr. Res. Pap. 1997, 44, 1701–1713. [Google Scholar] [CrossRef]
- Fairbanks, R.G.; Wiebe, P.H.; Bé, A.W.H. Vertical Distribution and Isotopic Composition of Living Planktonic Foraminifera in the Western North Atlantic. Science 1980, 207, 61–63. [Google Scholar] [CrossRef] [PubMed]
- Rebotim, A.; Voelker, A.H.L.; Jonkers, L.; Waniek, J.J.; Schulz, M.; Kucera, M. Calcification Depth of Deep-Dwelling Planktonic Foraminifera from the Eastern North Atlantic Constrained by Stable Oxygen Isotope Ratios of Shells from Stratified Plankton Tows. J. Micropalaeontol. 2019, 38, 113–131. [Google Scholar] [CrossRef]
- Mulitza, S.; Dürkoop, A.; Hale, W.; Wefer, G.; Stefan Niebler, H. Planktonic Foraminifera as Recorders of Past Surface-Water Stratification. Geolology 1997, 25, 335. [Google Scholar] [CrossRef]
- Ganssen, G.M.; Kroon, D. The Isotopic Signature of Planktonic Foraminifera from NE Atlantic Surface Sediments: Implications for the Reconstruction of Past Oceanic Conditions. J. Geol. Soc. 2000, 157, 693–699. [Google Scholar] [CrossRef]
- Kucera, M.; Weinelt, M.; Kiefer, T.; Pflaumann, U.; Hayes, A.; Weinelt, M.; Chen, M.-T.; Mix, A.C.; Barrows, T.T.; Cortijo, E.; et al. Reconstruction of Sea-Surface Temperatures from Assemblages of Planktonic Foraminifera: Multi-Technique Approach Based on Geographically Constrained Calibration Data Sets and Its Application to Glacial Atlantic and Pacific Oceans. Quat. Sci. Rev. 2005, 24, 951–998. [Google Scholar] [CrossRef]
- Ottens, J.J. Planktic Foraminifera as North Atlantic Water Mass Indicators. Oceanol. Acta 1991, 14, 123–140. [Google Scholar]
- Metcalfe, B.; Feldmeijer, W.; Ganssen, G.M. Oxygen Isotope Variability of Planktonic Foraminifera Provide Clues to Past Upper Ocean Seasonal Variability. Paleoceanogr. Paleoclimatol. 2019, 34, 374–393. [Google Scholar] [CrossRef]
- Chapman, M.R.; Shackleton, N.J. Global Ice-Volume Fluctuations, North Atlantic Ice-Rafting Events, and Deep-Ocean Circulation Changes between 130 and 70 Ka. Geology 1999, 27, 795. [Google Scholar] [CrossRef]
- Kretschmer, M.; Coumou, D.; Agel, L.; Barlow, M.; Tziperman, E.; Cohen, J. More-Persistent Weak Stratospheric Polar Vortex States Linked to Cold Extremes. Bull. Am. Meteorol. Soc. 2018, 99, 49–60. [Google Scholar] [CrossRef]
- Jonkers, L.; Kučera, M. Sensitivity to Species Selection Indicates the Effect of Nuisance Variables on Marine Microfossil Transfer Functions. Clim. Past 2019, 15, 881–891. [Google Scholar] [CrossRef]
- Rodrigues, T.; Alonso-García, M.; Hodell, D.A.; Rufino, M.; Naughton, F.; Grimalt, J.O.; Voelker, A.H.L.; Abrantes, F. A 1-Ma Record of Sea Surface Temperature and Extreme Cooling Events in the North Atlantic: A Perspective from the Iberian Margin. Quat. Sci. Rev. 2017, 172, 118–130. [Google Scholar] [CrossRef]
- Žarić, S.; Donner, B.; Fischer, G.; Mulitza, S.; Wefer, G. Sensitivity of Planktic Foraminifera to Sea Surface Temperature and Export Production as Derived from Sediment Trap Data. Mar. Micropaleontol. 2005, 55, 75–105. [Google Scholar] [CrossRef]
- Greco, M.; Jonkers, L.; Kretschmer, K.; Bijma, J.; Kucera, M. Depth Habitat of the Planktonic Foraminifera Neogloboquadrina Pachyderma in the northern high latitudes explained by sea-ice and chlorophyll concentrations. Biogeosciences 2019, 16, 3425–3437. [Google Scholar] [CrossRef]
- Rufino, M.M.; Salgueiro, E.; Voelker, A.A.H.L.; Polito, P.S.; Cermeño, P.A.; Abrantes, F. Ocean Kinetic Energy and Photosynthetic Biomass Are Important Drivers of Planktonic Foraminifera Diversity in the Atlantic Ocean. Front. Mar. Sci. 2022, 9, 887346. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rashid, H.; Chen, J.; Menke, M.; Zeng, M. Micropaleontological and Isotopic Perspective of Surface Water Mass Variability in the NE Atlantic from MIS 6 to 5e (188 to 115 ka). Geosciences 2023, 13, 149. https://doi.org/10.3390/geosciences13050149
Rashid H, Chen J, Menke M, Zeng M. Micropaleontological and Isotopic Perspective of Surface Water Mass Variability in the NE Atlantic from MIS 6 to 5e (188 to 115 ka). Geosciences. 2023; 13(5):149. https://doi.org/10.3390/geosciences13050149
Chicago/Turabian StyleRashid, Harunur, Jiaxin Chen, Mary Menke, and Min Zeng. 2023. "Micropaleontological and Isotopic Perspective of Surface Water Mass Variability in the NE Atlantic from MIS 6 to 5e (188 to 115 ka)" Geosciences 13, no. 5: 149. https://doi.org/10.3390/geosciences13050149