

Does Microbial and Faunal Pattern Correspond to Dynamics in Hydrogeology and Hydrochemistry? Comparative Study of Two Isolated Groundwater Ecosystems in Münsterland, Germany


Abstract
:1. Introduction
2. Materials and Methods
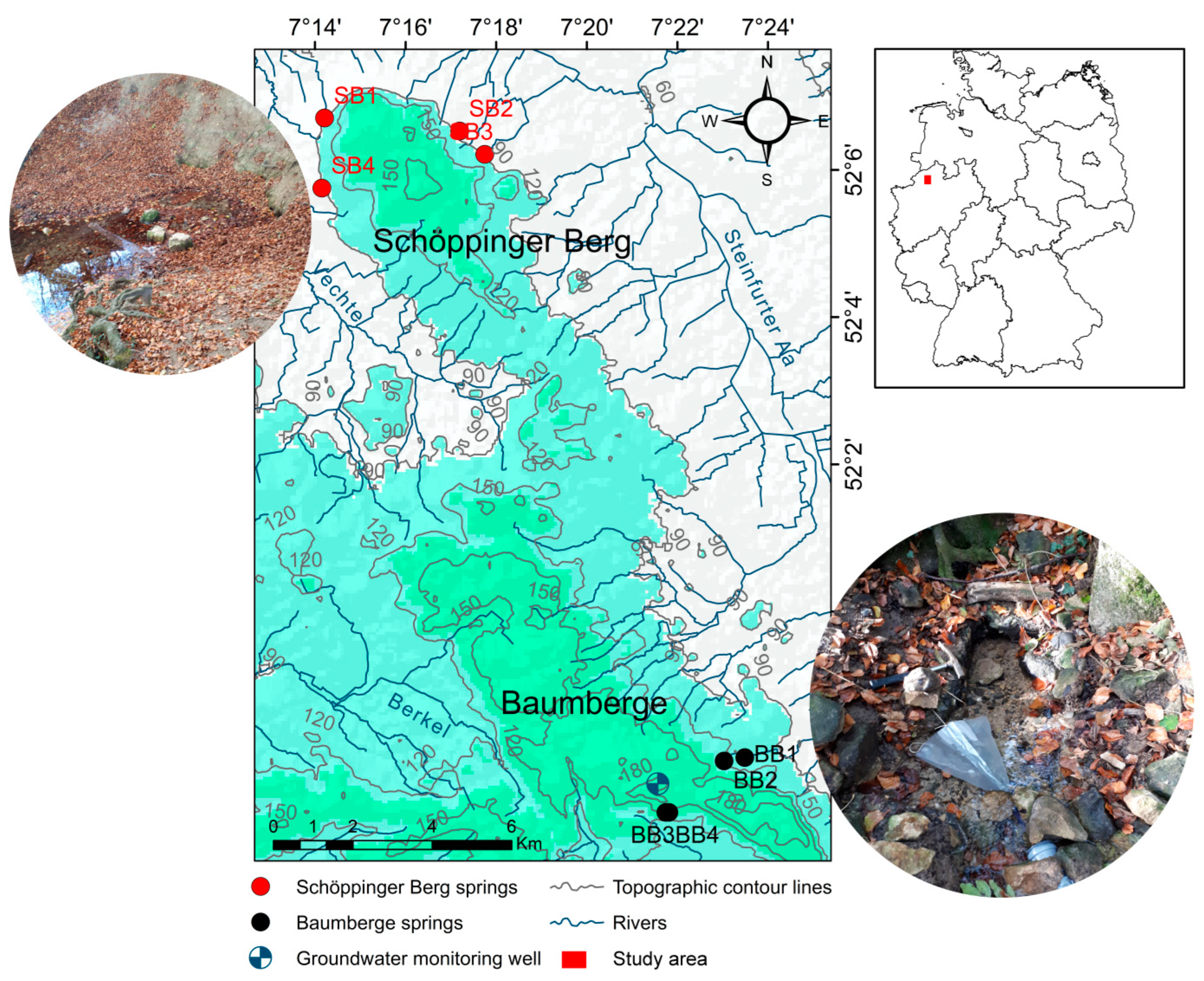
2.1. Study Area
2.2. Methodology
2.2.1. Field Work
Field Parameters and Aquatic Invertebrates Sampling
Hydro(geo)chemical and Microbiological Sampling
2.2.2. Laboratory Work
2.2.3. Data Processing
3. Results and Discussion
3.1. Physicochemical Parameters, Spring Discharge and Groundwater Table
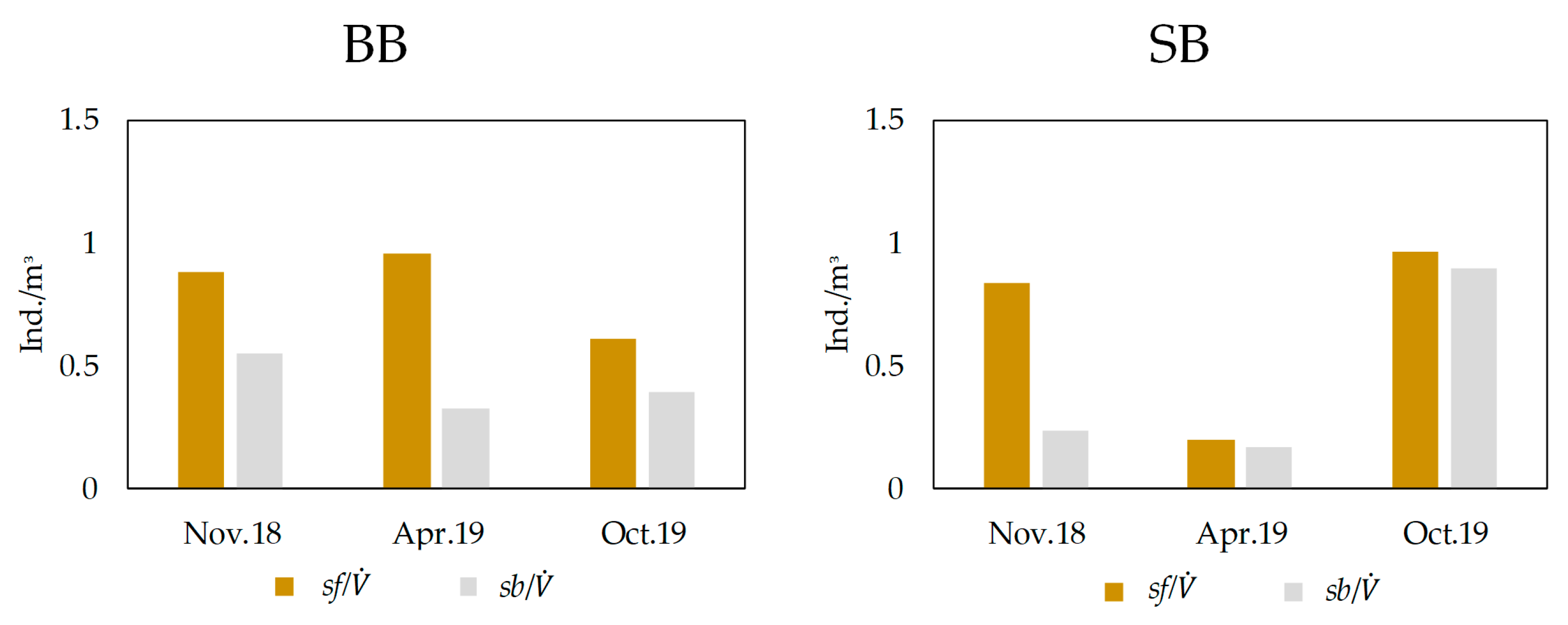
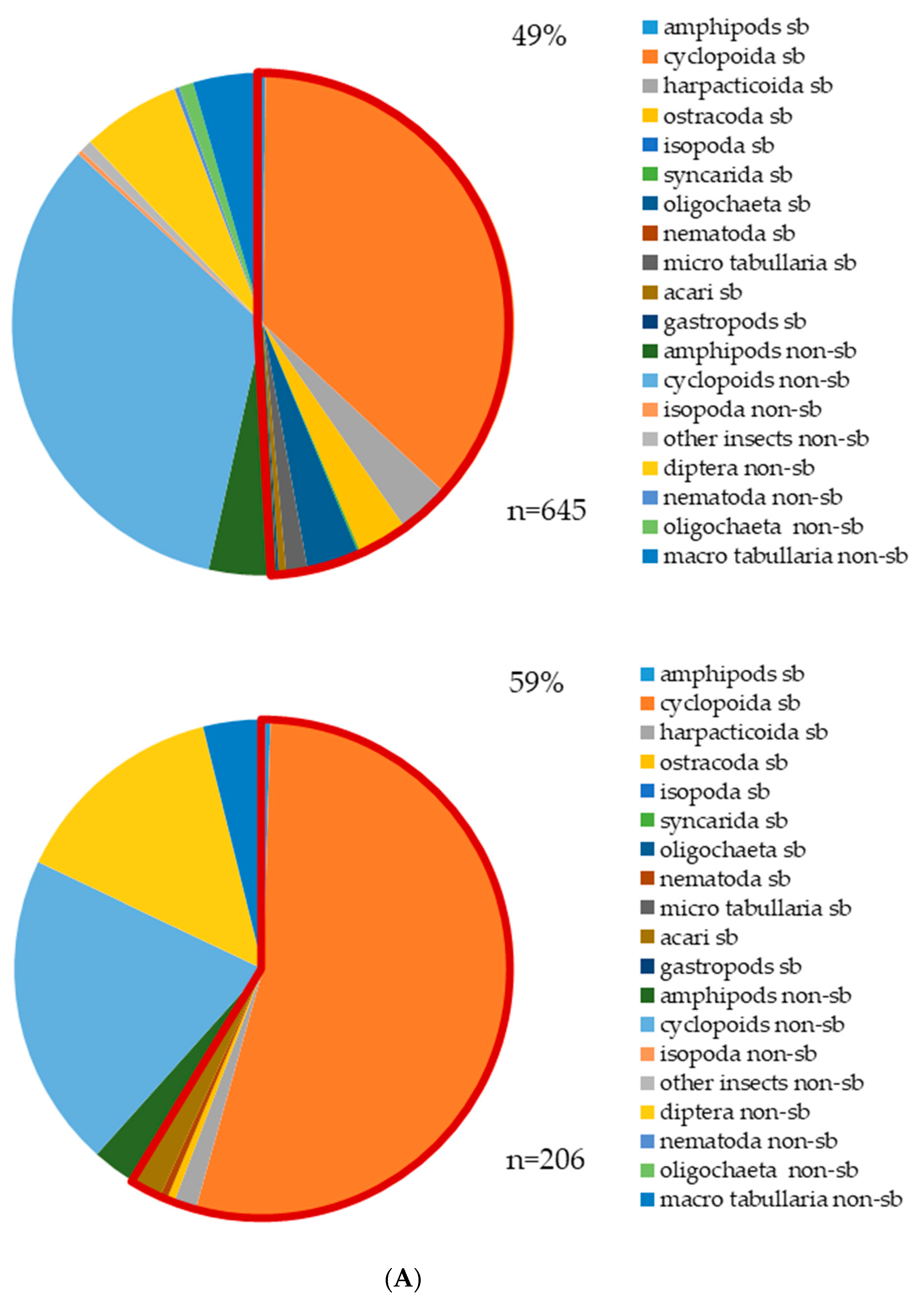
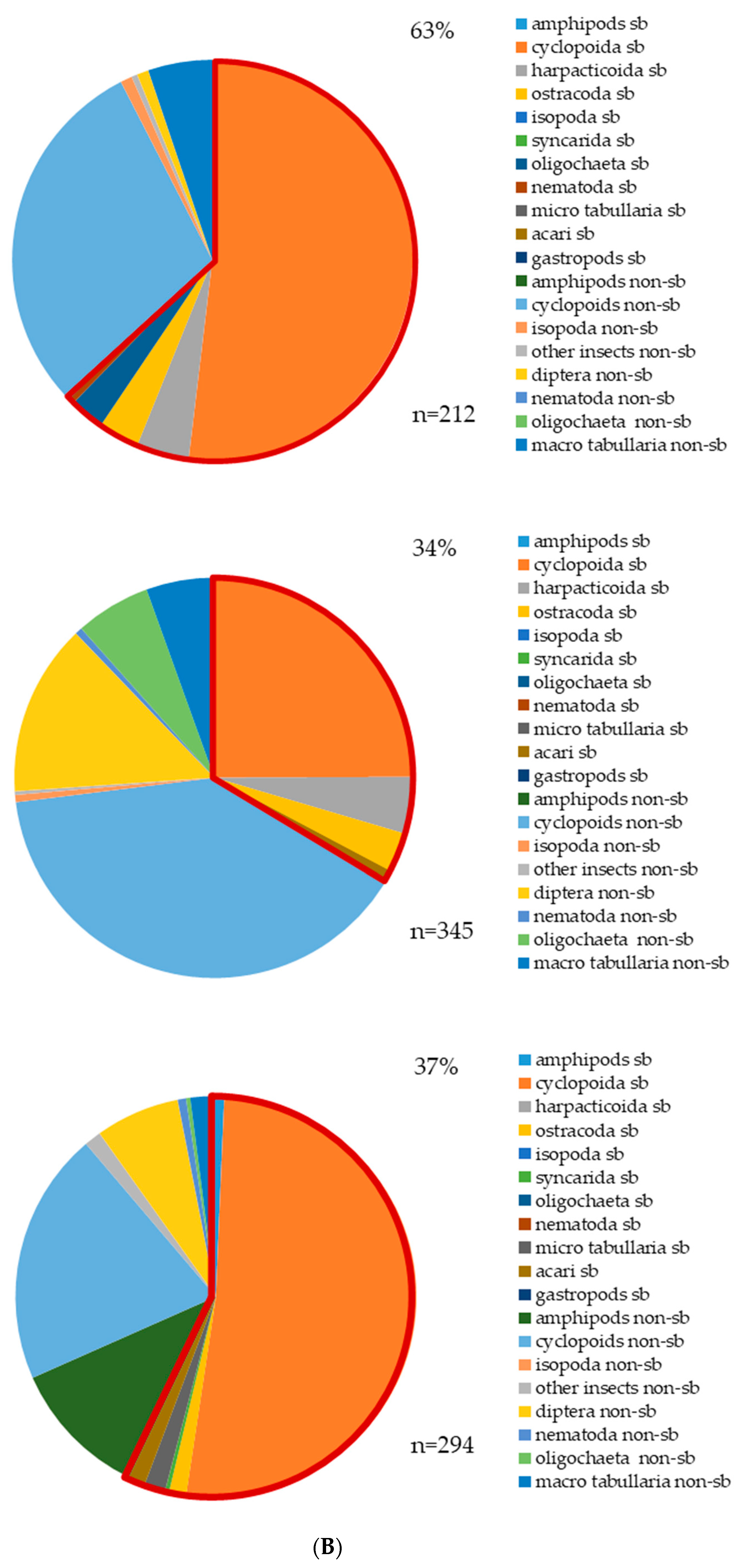
3.2. Aquatic Invertebrates (Stygofauna) and Stygobites
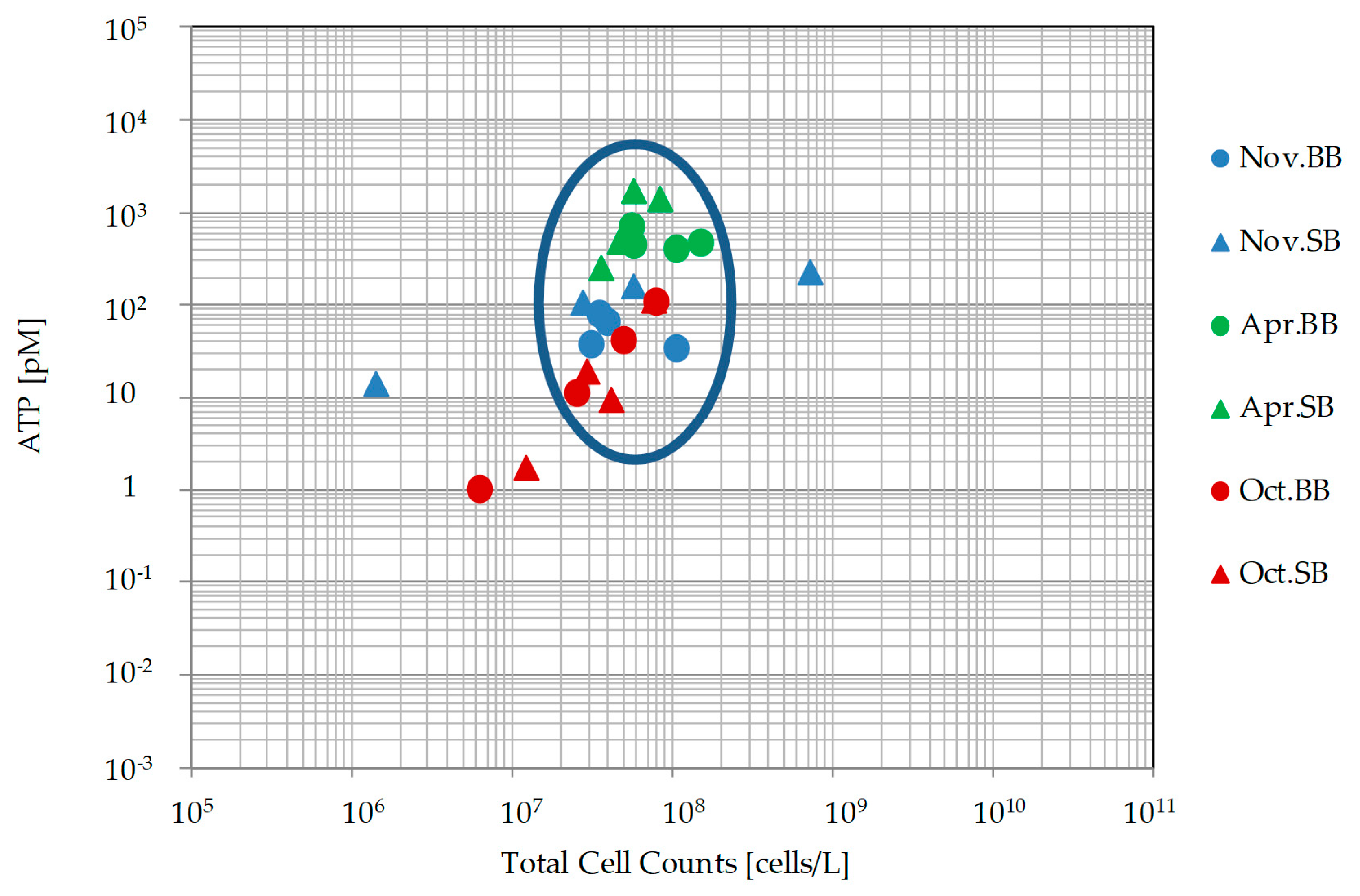
3.3. Microbiological Analyses (Biomass and Activity)
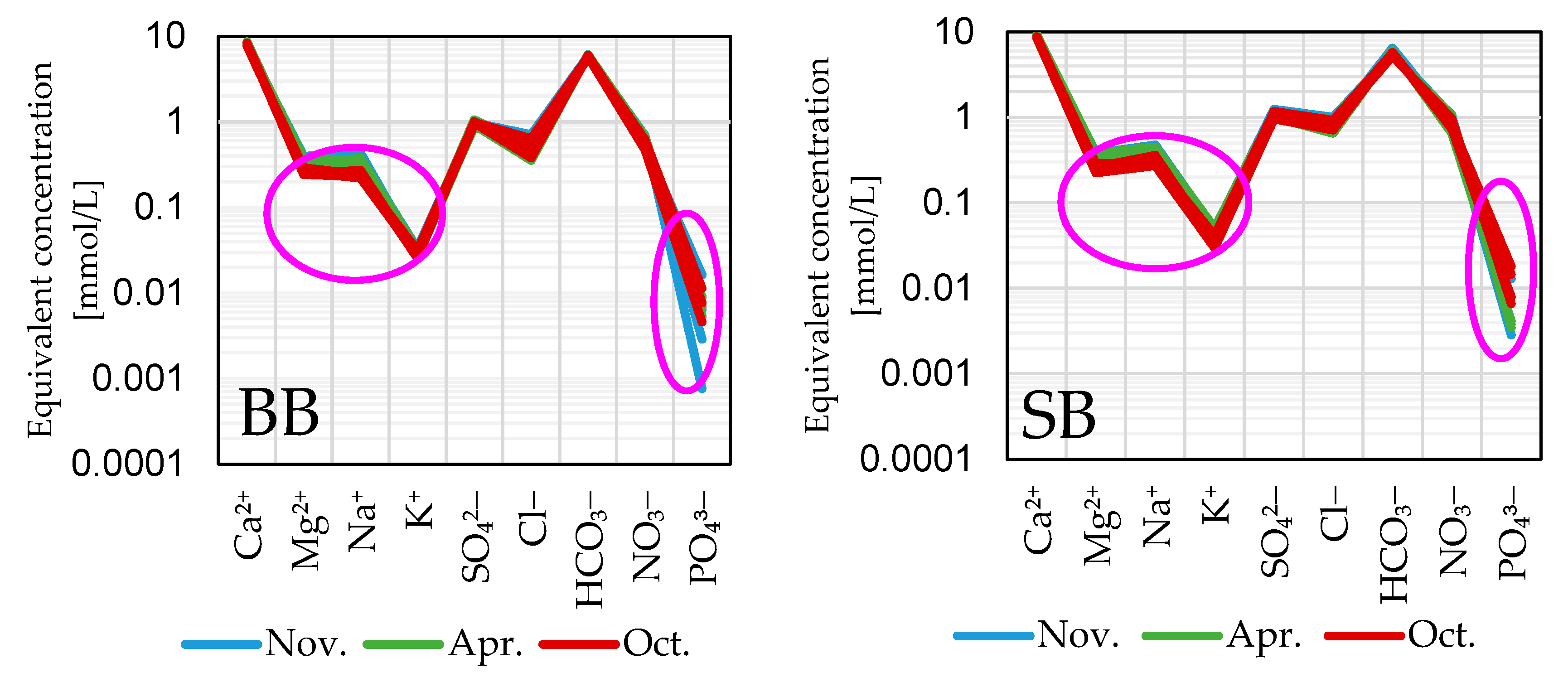
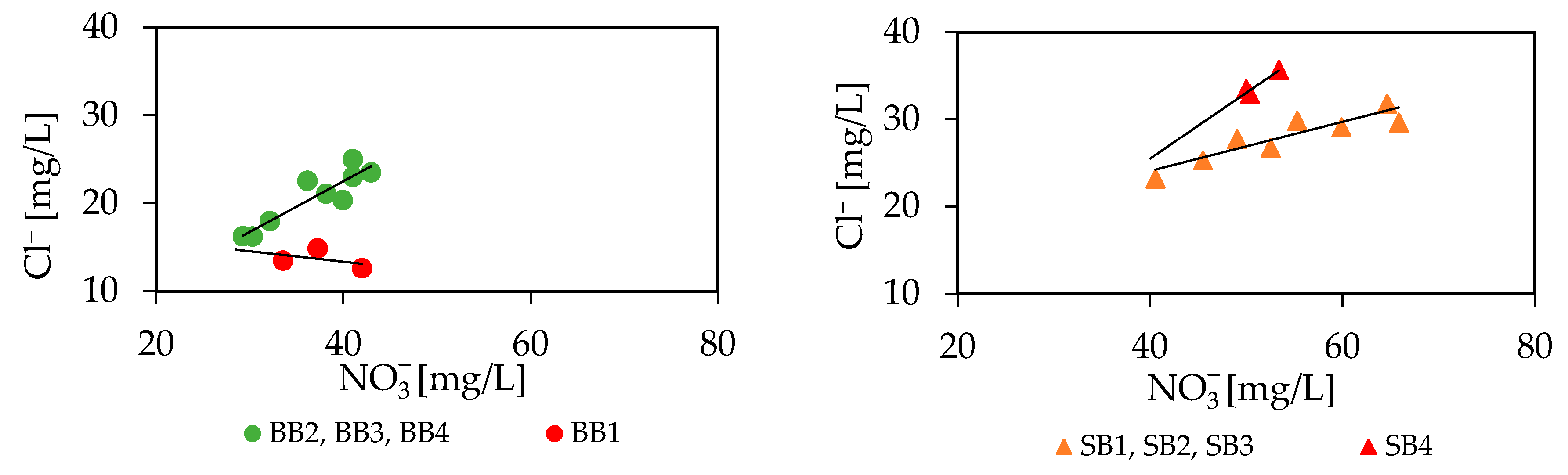
3.4. Groundwater Hydro(geo)chemistry
3.5. Stable Isotope Signatures in Spring Water
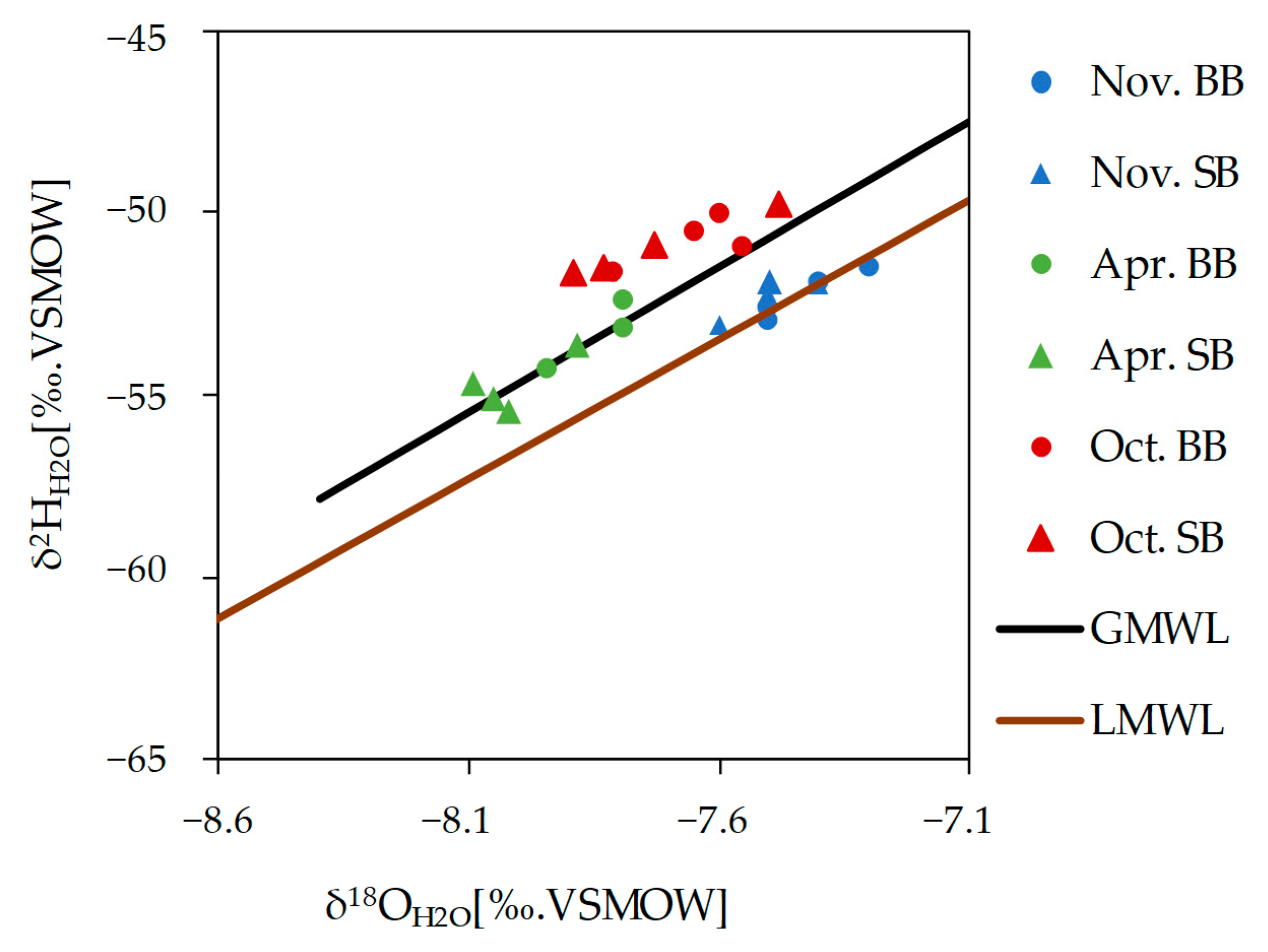
3.5.1. Stable Isotopes of Water (δ2H and δ18O)
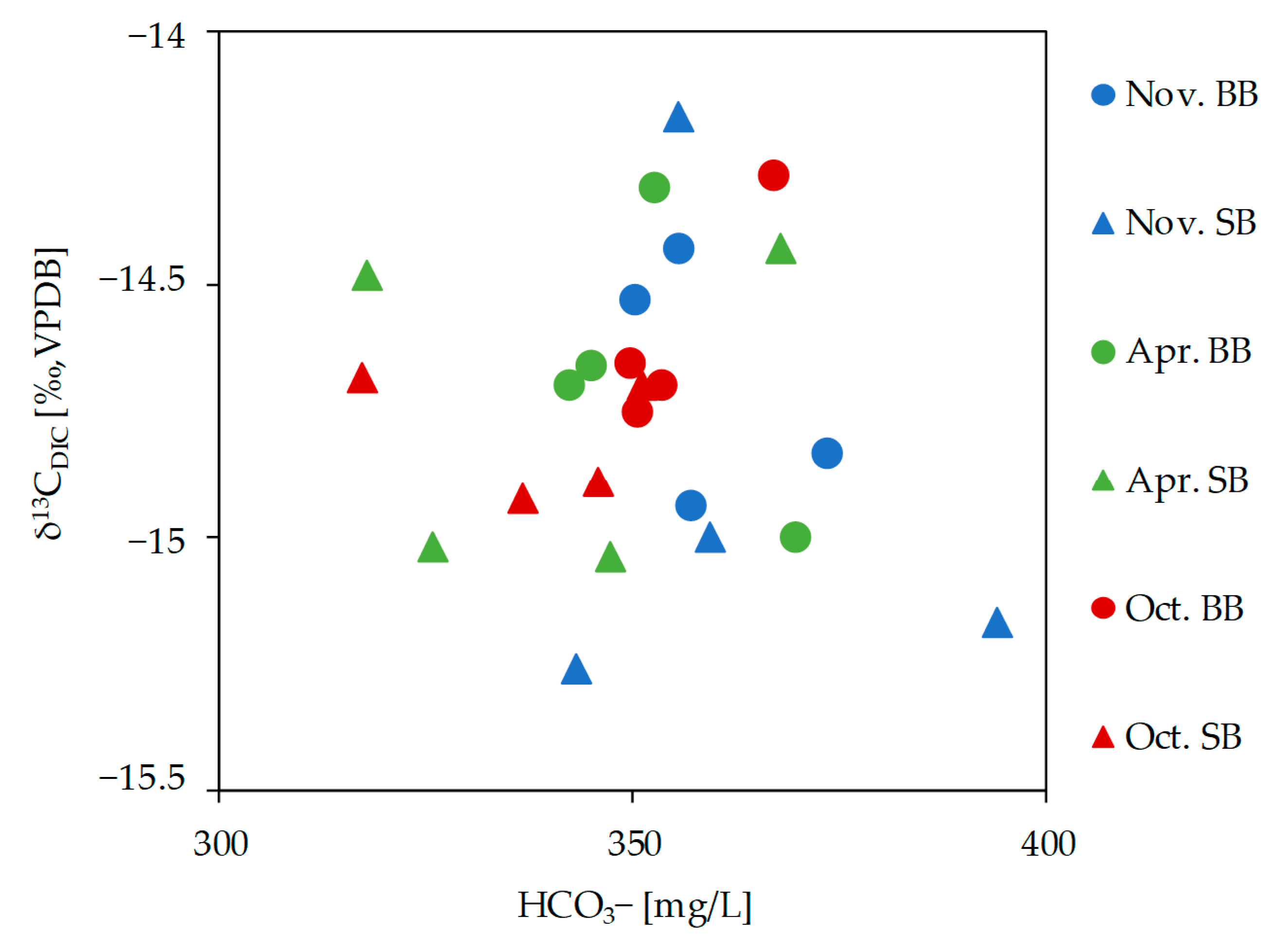
3.5.2. Stable Isotope of Dissolved Inorganic Carbon (δ13CDIC)
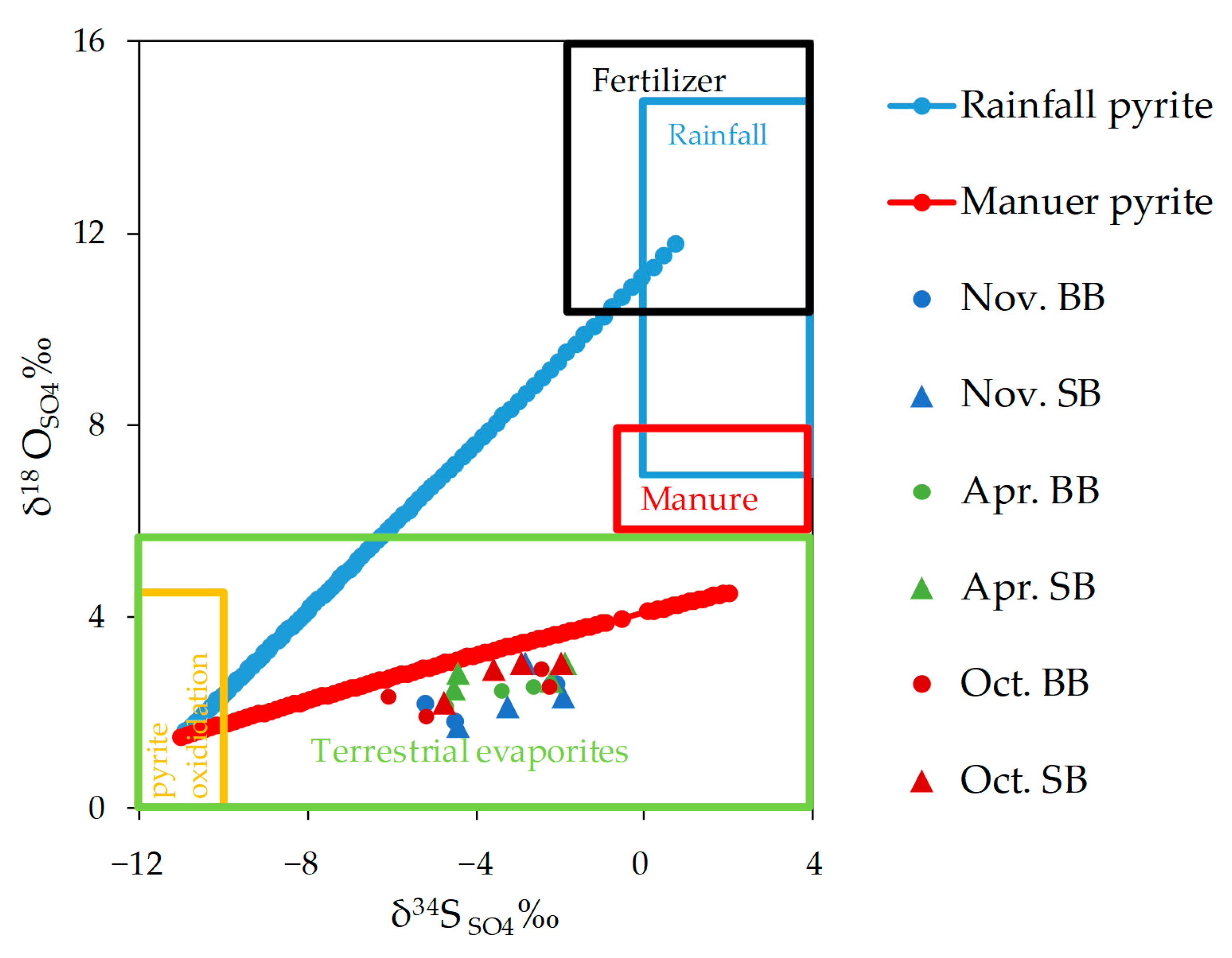
3.5.3. Stable Isotopes of Sulfate (δ34SSO4 and δ18OSO4)
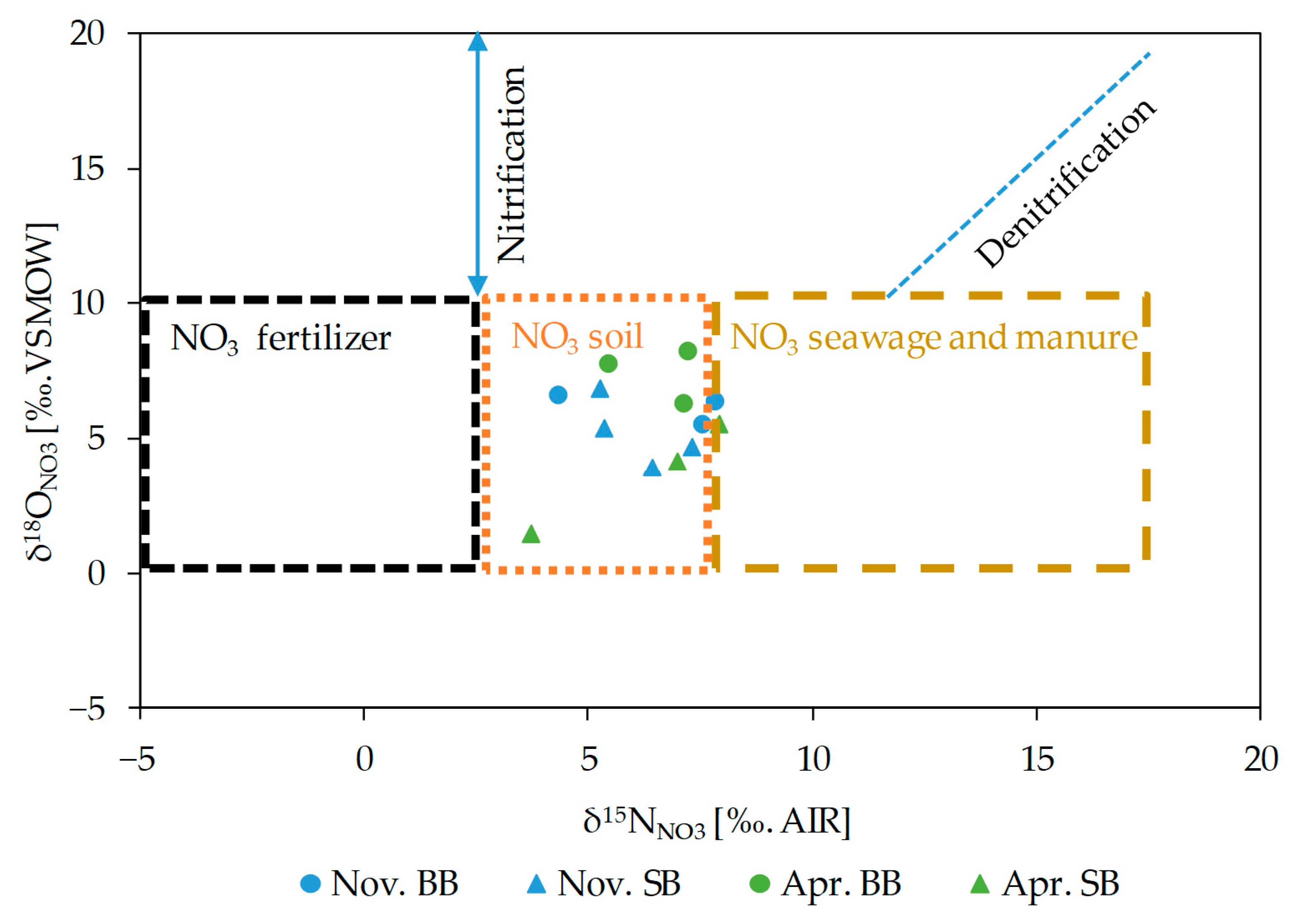
3.5.4. Stable Isotope of Nitrate (δ15NNO3 and δ18ONO3)
3.6. Statistical Analyses
3.6.1. Spearman Correlation
3.6.2. Principal Component Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Prasad, B.; Narayana, T. Subsurface Water Quality of Different Sampling Stations with Some Selected Parameters at Machilipatnam Town. Nat. Environ. Pollut. Technol. 2004, 3, 47–50. [Google Scholar]
- Gamar, A.; Zair, Z.; El Kabriti, M.; El Hilali, F. Study of the impact of the wild dump leachates of the region of El Hajeb (Morocco) on the physicochemical quality of the adjacent water table. Karbala Int. J. Mod. Sci. 2018, 4, 382–392. [Google Scholar] [CrossRef]
- Hose, G.C.; Symington, K.; Lategan, M.J.; Siegele, R. The Toxicity and Uptake of As, Cr and Zn in a Stygobitic Syncarid (Syncarida: Bathynellidae). Water 2019, 11, 2508. [Google Scholar] [CrossRef]
- Conrad, J.E.; Hirata, R.; Johansson, P.O.; Nonner, J.C.; Romijn, E.; Weaver, J.M.C. Groundwater Contamination Inventory: A Methodological Guide; Zaporozec, A., Ed.; UNESCO: Paris, France, 2002. [Google Scholar]
- Stein, H.; Kellermann, C.; Schmidt, S.I.; Brielmann, H.; Steube, C.; Berkhoff, S.E.; Fuchs, A.; Hahn, H.J.; Thulin, B.; Griebler, C. The potential use of fauna and bacteria as ecological indicators for the assessment of groundwater quality. J. Environ. Monit. 2010, 12, 242–254. [Google Scholar] [CrossRef]
- Aksever, F.; Davraz, A.; Karagüzel, R. Relations of hydrogeologic factors and temporal variations of nitrate contents in groundwater, Sandıklı basin, Turkey. Environ. Earth Sci. 2015, 73, 2179–2196. [Google Scholar] [CrossRef]
- Ma, F.; Chen, J.; Chen, J.; Wang, T.; Han, L.; Zhang, X.; Yan, J. Evolution of the hydro-ecological environment and its natural and anthropogenic causes during 1985–2019 in the Nenjiang River basin. Sci. Total Environ. 2021, 799, 149256. [Google Scholar] [CrossRef]
- Nisi, B.; Raco, B.; Dotsika, E. Groundwater Contamination Studies by Environmental Isotopes: A review. In Threats to the Quality of Groundwater Resources: Prevention and Control; Scozzari, A., Dotsika, E., Eds.; Springer: Berlin/Heidelberg, Germany, 2016; pp. 115–150. ISBN 978-3-662-48596-5. [Google Scholar]
- Sankoh, A.A.; Derkyi, N.; Frazer-williams, R.; Laar, C.; Kamara, I. A Review on the Application of Isotopic Techniques to Trace Groundwater Pollution Sources within Developing Countries. Water 2022, 14, 35. [Google Scholar] [CrossRef]
- Brunke, M.; Gonser, T.O.M. The ecological significance of exchange processes between rivers and groundwater. Freshw. Biol. 1997, 37, 1–33. [Google Scholar] [CrossRef]
- Soulsby, C.; Malcolm, R.; Gibbins, C.; Dilks, C. Seasonality, water quality trends and biological responses in four streams in the Cairngorm Mountains, Scotland. Hydrol. Earth Syst. Sci. 2001, 5, 433–450. [Google Scholar] [CrossRef]
- Ma, J.; Ibekwe, M.A.; Yang, C.H.; Crowley, D.E. Bacterial diversity and composition in major fresh produce growing soils affected by physiochemical properties and geographic locations. Sci. Total Environ. 2016, 563–564, 199–209. [Google Scholar] [CrossRef]
- Wright, J.; Kirchner, V.; Bernard, W.; Ulrich, N.; McLimans, C.; Campa, M.F.; Mackelprang, R. Bacterial community dynamics in dichloromethane-contaminated groundwater undergoing natural attenuation. Front. Microbiol. 2017, 8, 2300. [Google Scholar] [CrossRef] [PubMed]
- Kolar, B. The threshold concentration for nitrate in groundwater as a habitat of Proteus anguinus. Nat. Slov. 2018, 20, 39–42. [Google Scholar]
- Brkić, Ž.; Kuhta, M.; Larva, O.; Gottstein, S. Groundwater and connected ecosystems: An overview of groundwater body status assessment in Croatia. Environ. Sci. Eur. 2019, 31, 75. [Google Scholar] [CrossRef]
- Moral, F.; Cruz-Sanjulián, J.J.; Olías, M. Geochemical evolution of groundwater in the carbonate aquifers of Sierra de Segura (Betic Cordillera, southern Spain). J. Hydrol. 2008, 360, 281–296. [Google Scholar] [CrossRef]
- Gastmans, D.; Chang, H.K.; Hutcheon, I. Groundwater geochemical evolution in the northern portion of the Guarani Aquifer System (Brazil) and its relationship to diagenetic features. Appl. Geochem. 2010, 25, 16–33. [Google Scholar] [CrossRef]
- Li, X.; Huang, X.; Liao, X.; Zhang, Y. Hydrogeochemical Characteristics and Conceptual Model of the Geothermal Waters in the Xianshuihe Fault Zone, Southwestern China. Int. J. Environ. Res. Public Health 2020, 17, 500. [Google Scholar] [CrossRef]
- Foster, S.S.D.; Chilton, P.J. Groundwater: The processes and global significance of aquifer degradation. Philos. Trans. R. Soc. Lond. Ser. B-Biol. 2003, 358, 1957–1972. [Google Scholar] [CrossRef]
- Komatina, S.M. Geophysical methods application in groundwater natural protection against pollution. Environ. Geol. 1994, 23, 53–59. [Google Scholar] [CrossRef]
- Conboy, M.J.; Goss, M. Natural protection of groundwater against bacteria of fecal origin. J. Contam. Hydrol. 2000, 43, 1–24. [Google Scholar] [CrossRef]
- Morris, B.; Foster, S. Cryptosporidium contamination hazard assessment and risk management for British groundwater sources. Water Sci. Technol. 2000, 41, 67–77. [Google Scholar] [CrossRef]
- Haag, D.; Kaupenjohann, M. Landscape fate of nitrate fluxes and emissions in Central Europe—A critical review of concepts, data, and models for transport and retention. Agric. Ecosyst. Environ. 2001, 86, 1–21. [Google Scholar] [CrossRef]
- Foster, S.; Hirata, R. Groundwater Pollution Risk Assessment—A Methodology Using Available Data; CEPIS: Lima, Peru, 1998. [Google Scholar]
- Scow, K.M.; Hicks, K.A. Natural attenuation and enhanced bioremediation of organic contaminants in groundwater. Curr. Opin. Biotechnol. 2005, 16, 246–253. [Google Scholar] [CrossRef] [PubMed]
- Karczewski, K.; Göbel, P.; Meyer, E.I. Do composition and diversity of bacterial communities and abiotic conditions of spring water reflect characteristics of groundwater ecosystems exposed to different agricultural activities? Microbiologyopen 2019, 8, e00681. [Google Scholar] [CrossRef]
- Sasakova, N.; Gregova, G.; Takacova, D.; Mojzisova, J.; Papajova, I.; Venglovsky, J.; Szaboova, T.; Kovacova, S. Pollution of Surface and Ground Water by Sources Related to Agricultural Activities. Front. Sustain. Food Syst. 2018, 2, 42. [Google Scholar] [CrossRef]
- Saccò, M.; Blyth, A.; Bateman, P.W.; Hua, Q.; Mazumder, D.; White, N.; Humphreys, W.F.; Laini, A.; Griebler, C.; Grice, K. New light in the dark—A proposed multidisciplinary framework for studying functional ecology of groundwater fauna. Sci. Total Environ. 2019, 662, 963–977. [Google Scholar] [CrossRef]
- Hahn, H.J.; Fuchs, A. Distribution patterns of groundwater communities across aquifer types in south-western Germany. Freshw. Biol. 2009, 54, 848–860. [Google Scholar] [CrossRef]
- Maurice, L.; Bloomfield, J. Stygobitic Invertebrates in Groundwater—A Review from a Hydrogeological Perspective. Freshw. Rev. 2012, 5, 51–71. [Google Scholar] [CrossRef]
- Griebler, C.; Fillinger, L.; Karwautz, C.; Hose, G.C. Knowledge Gaps, Obstacles, and Research Frontiers in Groundwater Microbial Ecology. In Encyclopedia of Inland Waters; Elsevier: Amsterdam, The Netherlands, 2022; pp. 611–624. ISBN 9780128220412. [Google Scholar]
- Cardinale, B.J.; Srivastava, D.S.; Emmett Duffy, J.; Wright, J.P.; Downing, A.L.; Sankaran, M.; Jouseau, C. Effects of biodiversity on the functioning of trophic groups and ecosystems. Nature 2006, 443, 989–992. [Google Scholar] [CrossRef]
- Gamfeldt, L.; Snäll, T.; Bagchi, R.; Jonsson, M.; Gustafsson, L.; Kjellander, P.; Ruiz-Jaen, M.C.; Fröberg, M.; Stendahl, J.; Philipson, C.D.; et al. Higher levels of multiple ecosystem services are found in forests with more tree species. Nat. Commun. 2013, 4, 1340. [Google Scholar] [CrossRef]
- Savio, D.; Stadler, P.; Reischer, G.H.; Kirschner, A.K.T.; Demeter, K.; Linke, R.; Blaschke, A.P.; Sommer, R.; Szewzyk, U.; Wilhartitz, I.C.; et al. Opening the black box of spring water microbiology from alpine karst aquifers to support proactive drinking water resource management. WIREs Water 2018, 5, e1282. [Google Scholar] [CrossRef]
- Hose, G.C.; Lategan, M.J. Sampling Strategies for Biological Assessment of Groundwater System; CRC for Contamination Assessment and and Remediation of the Environment: Adelaide, Australia, 2012. [Google Scholar]
- Malard, F.; Mathieu, J.; Reygrobellet, J.-L.; Lafont, M. Biomotoring groundwater contamination: Application to a karst area in Southern France. Aquat. Sci. 1996, 58, 158–187. [Google Scholar] [CrossRef]
- Alqaragholi, S.A.; Kanoua, W.; Göbel, P. Comparative Investigation of Aquatic Invertebrates in Springs in Münsterland Area (Western Germany). Water 2021, 13, 359. [Google Scholar] [CrossRef]
- Hahn, H.J. The GW-Fauna-Index: A first approach to a quantitative ecological assessment of groundwater habitats. Limnologica 2006, 36, 119–137. [Google Scholar] [CrossRef]
- Fillinger, L.; Hug, K.; Trimbach, A.M.; Wang, H.; Kellermann, C.; Meyer, A.; Bendinger, B.; Griebler, C. The D-A-(C) index: A practical approach towards the microbiological-ecological monitoring of groundwater ecosystems. Water Res. 2019, 163, 114902. [Google Scholar] [CrossRef] [PubMed]
- Besmer, M.D.; Hammes, F. Short-term microbial dynamics in a drinking water plant treating groundwater with occasional high microbial loads. Water Res. 2016, 107, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Hammes, F.; Berney, M.; Wang, Y.; Vital, M.; Köster, O.; Egli, T. Flow-cytometric total bacterial cell counts as a descriptive microbiological parameter for drinking water treatment processes. Water Res. 2008, 42, 269–277. [Google Scholar] [CrossRef]
- Parkhurst, D.L.; Appelo, C.A.J. Description of Input for PHREEQC Version 3—A Computer Program for Speciation, Batch-Reaction, One-Dimensional Transport, and Inverse Geochemical Calculations; US Geological Survey, Water Resources Division: Denver, CO, USA, 2013.
- Foulquier, A.; Malard, F.; Mermillod-Blondin, F.; Montuelle, B.; Dolédec, S.; Volat, B.; Gibert, J. Surface Water Linkages Regulate Trophic Interactions in a Groundwater Food Web. Ecosystems 2011, 14, 1339–1353. [Google Scholar] [CrossRef]
- Bero, N.J.; Ruark, M.D.; Lowery, B. Bromide and chloride tracer application to determine sufficiency of plot size and well depth placement to capture preferential flow and solute leaching. Geoderma 2016, 262, 94–100. [Google Scholar] [CrossRef]
- Graig, H. Isotopic Variations in Meteoric Waters. Science 1961, 133, 1702–1703. [Google Scholar]
- GNIP. Global Network of Isotopes in Precipitation. 2022. Available online: https://www.iaea.org/services/networks/gnip (accessed on 1 April 2020).
- DWD. Deutcher Wetterdienst, 2018–2019. Available online: https://www.dwd.de/DE/Home/home_node.html (accessed on 1 February 2018).
- ELWAS. Elektronisches Wasserwirtschaftliches Verbundsystem für die Wasserwirtschaftsverwaltung in NRW. 2018. Available online: https://www.elwasweb.nrw.de/elwas-web/index.xhtml;jsessionid=7CBE640FD1D179B191EBF354D0F3FD13 (accessed on 1 January 2020).
- Shen, Z.; Zhu, Y.; Zhong, Y. Hydrogeochemistry; Geological Publishing House: Beijing, China, 1993. [Google Scholar]
- Appelo, C.; Postma, D. Geochemistry, Groundwater and Pollution; Balkema Publishers: Amsterdam, The Netherlands, 2005; pp. 1–634. [Google Scholar]
- Einsiedl, F.; Pilloni, G.; Ruth-Anneser, B.; Lueders, T.; Griebler, C. Spatial distributions of sulphur species and sulphate-reducing bacteria provide insights into sulphur redox cycling and biodegradation hot-spots in a hydrocarbon-contaminated aquifer. Geochim. Cosmochim. Acta 2015, 156, 207–221. [Google Scholar] [CrossRef]
- Göbel, P.; Römer, M.; Weckwert, N.; Alqaragholi, S.A.; Hahn, H.J.; Meyer, E.I.; Knöller, K.; Strauss, H. Hydro(geo)chemische und ökologische Bestandsaufnahme von Quellregionen als isolierte Grundwasser-Ökosysteme. Grund. Z. Fachsekt. Hydrogeol. 2022, 27, 277–293. [Google Scholar] [CrossRef]
- Vitòria, L.; Otero, N.; Soler, A.; Canals, A. Fertilizer characterization: Isotopic data (N, S, O, C, and Sr). Environ. Sci. Technol. 2004, 38, 3254–3262. [Google Scholar] [CrossRef] [PubMed]
- Stenger, R.; Clague, J.; Woodward, S.; Moorhead, B.; Wilson, S.; Shokri, A.; Wöhling, T.; Canard, H. Dentrification—The Key Component of a Groundwter Systems Assimplative Carpacity for Nitrate; Fertilizer and Lime Research Centre, Massey University: Palmerston North, New Zealand, 2013. [Google Scholar]
- Schwientek, M.; Einsiedl, F.; Stichler, W.; Stögbauer, A.; Strauss, H.; Maloszewski, P. Evidence for denitrification regulated by pyrite oxidation in a heterogeneous porous groundwater system. Chem. Geol. 2008, 255, 60–67. [Google Scholar] [CrossRef]
- Cravotta, C.A. Use of Stable Isotopes of Carbon, Nitrogen, and Sulfur to Identify Sources of Nitrogen in Surface Waters in the Lower Susquehanna River Basin, Pennsylvania; For sale by the U.S. Geological Survey Branch of Information Services: Washington, DC, USA; Denver, CO, USA, 1997; ISBN 0607872071.
- Toomanian, N.; Jalalian, A.; Eghbal, M.K. Genesis of gypsum enriched soils in north-west Isfahan, Iran. Geoderma 2001, 99, 199–224. [Google Scholar] [CrossRef]
- Buss, J.; Achten, C. Spatiotemporal variations of surface water quality in a medium-sized river catchment (Northwestern Germany) with agricultural and urban land use over a five-year period with extremely dry summers. Sci. Total Environ. 2022, 818, 151730. [Google Scholar] [CrossRef]
- Kendall, C.; Emily, M.E.; Scott, D.W. Tracing anthropogenic inputs of nitrogen to ecosytems. In Stable Isotopes in Ecology and Environmental Science; Michener, R., Lajtha, K., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2007. [Google Scholar]
- Holman, I.P.; Whelan, M.J.; Howden, N.J.K.; Bellamy, P.H.; Willby, N.J.; Rivas-Casado, M.; McConvey, P. Phosphorus in groundwater-an overlooked contributor to eutrophication? Hydrol. Process. 2008, 22, 5121–5127. [Google Scholar] [CrossRef]
- Lewandowski, J.; Meinikmann, K.; Nützmann, G.; Rosenberry, D.O. Groundwater—The disregarded component in lake water and nutrient budgets. Part 2: Effects of groundwater on nutrients. Hydrol. Process. 2015, 29, 2922–2955. [Google Scholar] [CrossRef]
- Griebler, C.; Stein, H.; Kellermanna, C.; Berkhoff, S.E.; Brielmanna, H. Ecological assessment of groundwater ecosystems—Vision or illusion? Ecol. Eng. 2010, 36, 1174–1190. [Google Scholar] [CrossRef]
- Fadiran, A.O.; Dlamini, S.C.; Mavuso, A. A comparative study of the phosphate levels in some surface and groundwater bodies of Swaziland. Bull. Chem. Soc. Ethiop. 2008, 22, 197–206. [Google Scholar] [CrossRef]
- Subhas, A.V.; Adkins, J.F.; Rollins, N.E.; Naviaux, J.; Erez, J.; Berelson, W.M. Catalysis and chemical mechanisms of calcite dissolution in seawater. Proc. Natl. Acad. Sci. USA 2017, 114, 8175–8180. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | PC1 | PC2 | PC3 | PC4 | PC5 |
|---|---|---|---|---|---|
| Na+ | 0.98 | ||||
| SiO32− | −0.96 | ||||
| Mg2+ | 0.86 | ||||
| Sr2+ | 0.84 | ||||
| NO3− | 0.85 | ||||
| SO42− | −0.81 | ||||
| Detritus | −0.81 | ||||
| Ca2+ | 0.80 | ||||
| ATP | 0.90 | ||||
| hGW | 0.89 | ||||
| δ18OH2O | −0.89 | ||||
| Cl− | −0.87 | ||||
| HCO3− | 0.85 | ||||
| −0.84 | |||||
| PO43− | 0.89 | ||||
| sb/ | 0.79 | ||||
| GFI | 0.78 | ||||
| sf/ | 0.73 | ||||
| % of Variance | 25.42 | 22.23 | 14.68 | 11.71 | 8.14 |
| Cumulative% | 25.42 | 47.65 | 62.34 | 74.05 | 82.20 |
| Parameter | PC1 | PC2 | PC3 | PC4 | PC5 |
|---|---|---|---|---|---|
| EC | 0.90 | ||||
| δ34SSO4 | −0.87 | ||||
| K+ | 0.82 | ||||
| Detritus | −0.82 | ||||
| δ18OSO4 | −0.81 | ||||
| Ca2+ | 0.81 | ||||
| Temp. | 0.71 | ||||
| DO | −0.82 | ||||
| δ18OH2O | 0.80 | ||||
| hGW | −0.73 | ||||
| ATP | −0.72 | ||||
| δ2HH2O | 0.71 | ||||
| Na+ | 0.89 | ||||
| Mg2+ | 0.84 | ||||
| SiO32− | −0.80 | ||||
| δ13CDIC | 0.96 | ||||
| Fe2+ | 0.94 | ||||
| Al3+ | 0.92 | ||||
| Cl− | 0.89 | ||||
| sb/ | −0.82 | ||||
| % of Variance | 29.96 | 22.20 | 12.87 | 11.03 | 8.11 |
| Cumulative% | 29.96 | 52.16 | 65.03 | 76.06 | 84.17 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alqaragholi, S.A.; Kanoua, W.; Strauss, H.; Göbel, P. Does Microbial and Faunal Pattern Correspond to Dynamics in Hydrogeology and Hydrochemistry? Comparative Study of Two Isolated Groundwater Ecosystems in Münsterland, Germany. Geosciences 2023, 13, 140. https://doi.org/10.3390/geosciences13050140
Alqaragholi SA, Kanoua W, Strauss H, Göbel P. Does Microbial and Faunal Pattern Correspond to Dynamics in Hydrogeology and Hydrochemistry? Comparative Study of Two Isolated Groundwater Ecosystems in Münsterland, Germany. Geosciences. 2023; 13(5):140. https://doi.org/10.3390/geosciences13050140
Chicago/Turabian StyleAlqaragholi, Sura Abdulghani, Wael Kanoua, Harald Strauss, and Patricia Göbel. 2023. "Does Microbial and Faunal Pattern Correspond to Dynamics in Hydrogeology and Hydrochemistry? Comparative Study of Two Isolated Groundwater Ecosystems in Münsterland, Germany" Geosciences 13, no. 5: 140. https://doi.org/10.3390/geosciences13050140