Single Nucleotide Polymorphisms in SLC5A1, CCNA1, and ABCC1 and the Association with Litter Size in Small-Tail Han Sheep


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Detection of SLC5A1, CCNA1, and ABCC1 Expression by Real-Time PCR
2.3. Genotyping
2.4. Statistical Analysis
3. Results
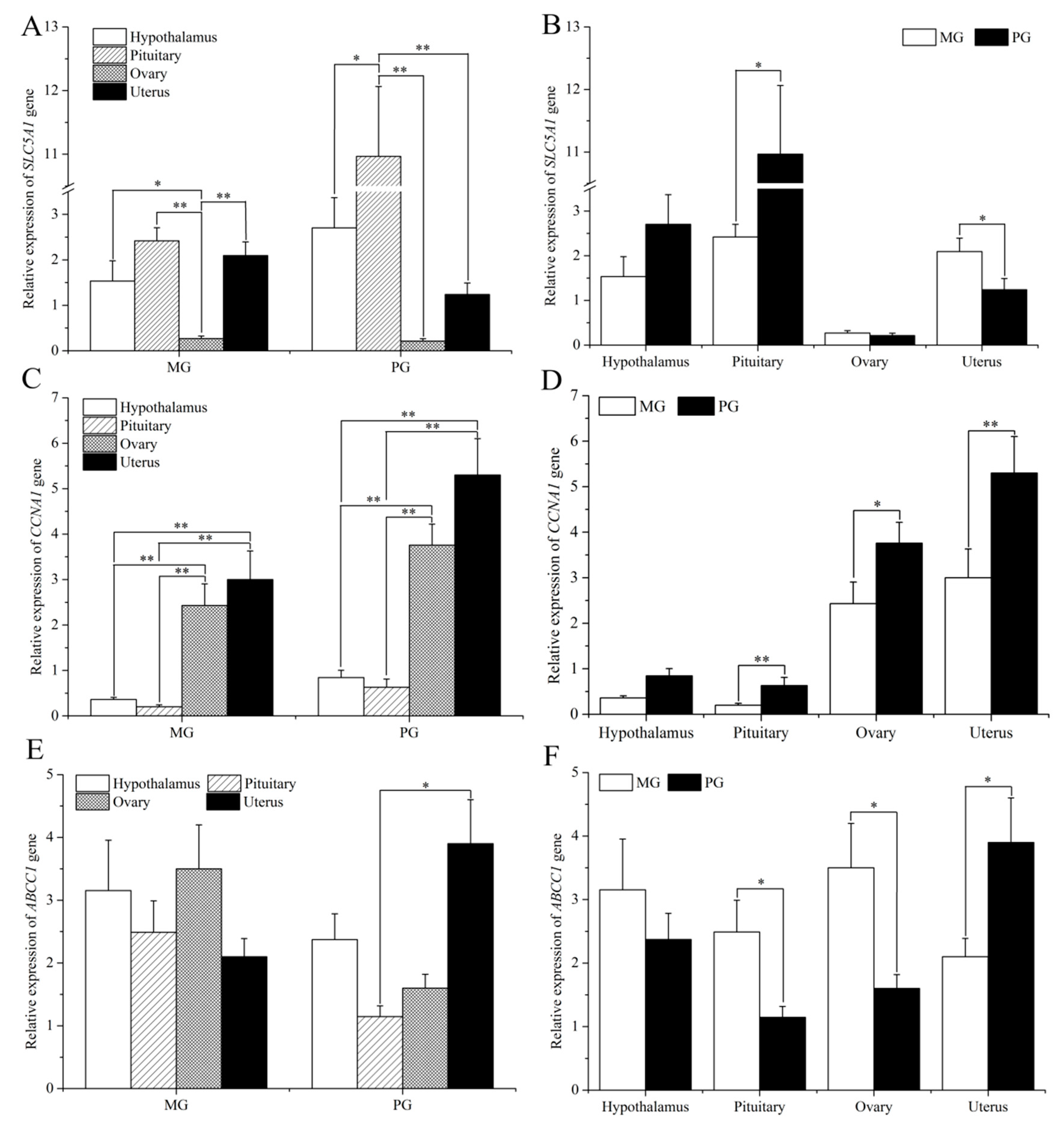
3.1. Expression of SLC5A1, CCNA1, and ABCC1 in the HPG Axis of Small-Tail Han Sheep with Different Litter Sizes
3.2. Population Genetic Analysis of Polymorphism in the SLC5A1, CCNA1 and ABCC1 Genes
3.3. Association Analysis of SNPs with Litter Size in Small-Tail Han Sheep
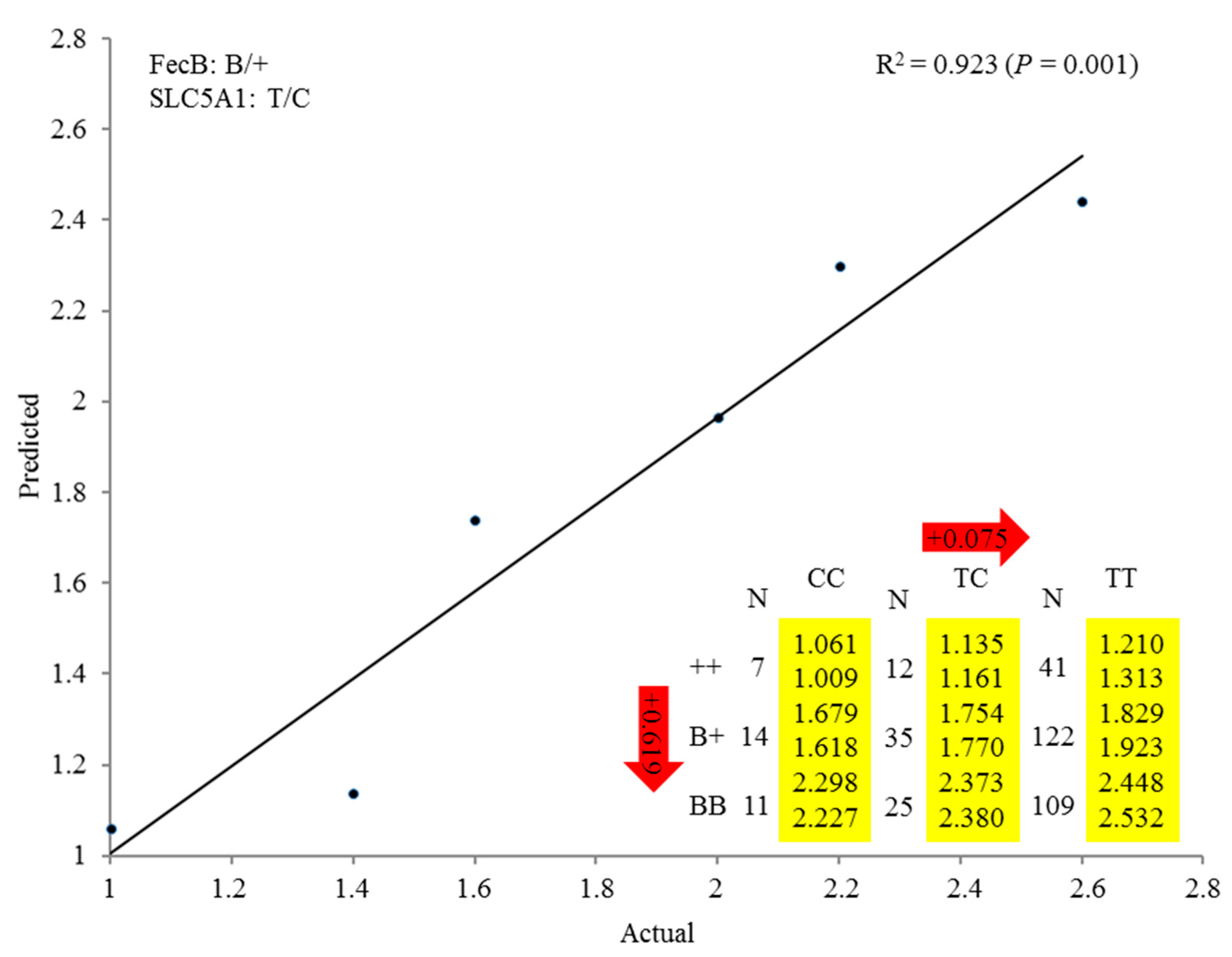
3.4. Regression Analysis of SLC5A1 and BMPR-IB Associated with Average Litter Size in Small-Tail Han Sheep
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Chen, H.Y.; Shen, H.; Jia, B.; Zhang, Y.S.; Wang, X.H.; Zeng, X.C. Differential gene expression in ovaries of Qira black sheep and Hetian sheep using RNA-Seq technique. PLoS ONE 2015, 10, e0120170. [Google Scholar] [CrossRef] [PubMed]
- Miao, X.Y.; Luo, Q.M.; Zhao, H.J.; Qin, X.Y. Co-expression analysis and identification of fecundity-related long non-coding RNAs in sheep ovaries. Sci. Rep. 2016, 6, 39398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bravo, S.; Larama, G.; Paz, E.; Inostroza, K.; Montaldo, H.H.; Sepulveda, N. Polymorphism of the GDF9 gene associated with litter size in Araucana creole sheep. Anim. Genet. 2016, 47, 390–391. [Google Scholar] [CrossRef] [PubMed]
- Davis, G.H. Major genes affecting ovulation rate in sheep. Genet. Sel. Evol. 2005, 37, S11–S23. [Google Scholar] [CrossRef] [PubMed]
- Chu, M.X.; Liu, Z.H.; Jiao, C.L.; He, Y.Q.; Fang, L.; Ye, S.C.; Chen, G.H.; Wang, J.Y. Mutations in BMPR-IB and BMP-15 genes are associated with litter size in Small Tailed Han sheep (Ovis aries). J. Anim. Sci. 2007, 85, 598–603. [Google Scholar] [CrossRef] [PubMed]
- Mulsant, P.; Lecerf, F.; Fabre, S.; Schibler, L.; Monget, P.; Lanneluc, I.; Pisselet, C.; Riquet, J.; Monniaux, D.; Callebaut, I.; et al. Mutation in bone morphogenetic protein Receptor-IB is associated with increased ovulation rate in Booroola Merino ewes. Proc. Natl. Acad. Sci. USA 2001, 98, 5104–5109. [Google Scholar] [CrossRef]
- Souza, C.J.H.; MacDougall, C.; Campbell, B.K.; McNeilly, A.S.; Baird, D.T. The Booroola (FecB) phenotype is associated with a mutation in the bone morphogenetic receptor type 1B (BMPR1B) gene. J. Endocrinol. 2001, 169, R1–R6. [Google Scholar] [CrossRef]
- Bodin, L.; Di Pasquale, E.; Fabre, S.; Bontoux, M.; Monget, P.; Persani, L.; Mulsant, P. A novel mutation in the bone morphogenetic protein 15 gene causing defective protein secretion is associated with both increased ovulation rate and sterility in Lacaune sheep. Endocrinology 2007, 148, 393–400. [Google Scholar] [CrossRef]
- Galloway, S.M.; McNatty, K.P.; Cambridge, L.M.; Laitinen, M.P.E.; Juengel, J.L.; Jokiranta, T.S.; McLaren, R.J.; Luiro, K.; Dodds, K.G.; Montgomery, G.W.; et al. Mutations in an oocyte-derived growth factor gene (BMP15) cause increased ovulation rate and infertility in a dosage-sensitive manner. Nat. Genet. 2000, 25, 279–283. [Google Scholar] [CrossRef]
- Hanrahan, J.P.; Gregan, S.M.; Mulsant, P.; Mullen, M.; Davis, G.H.; Powell, R.; Galloway, S.M. Mutations in the genes for oocyte-derived growth factors GDF9 and BMP15 are associated with both increased ovulation rate and sterility in Cambridge and Belclare sheep (Ovis aries). Biol. Reprod. 2004, 70, 900–909. [Google Scholar] [CrossRef]
- Chu, M.X.; Li, B.X.; Wang, J.Y.; Ye, S.C.; Fang, L. Association between PCR-SSCP of growth differentiation factor 9 gene and high prolificacy in Small-Tail Han sheep. Anim. Biotechnol. 2004, 15, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Silva, B.D.M.; Castro, E.A.; Souza, C.J.H.; Paiva, S.R.; Sartori, R.; Franco, M.M.; Azevedo, H.C.; Silva, T.A.S.N.; Vieira, A.M.C.; Neves, J.P.; et al. A new polymorphism in the Growth and Differentiation Factor 9 (GDF9) gene is associated with increased ovulation rate and prolificacy in homozygous sheep. Anim. Genet. 2011, 42, 89–92. [Google Scholar] [CrossRef] [Green Version]
- Chu, M.X.; Yang, J.; Feng, T.; Cao, G.L.; Fang, L.; Di, R.; Huang, D.W.; Tang, Q.Q.; Ma, Y.H.; Li, K.; et al. GDF9 as a candidate gene for prolificacy of Small-Tail Han sheep. Mol. Biol. Rep. 2011, 38, 5199–5204. [Google Scholar] [CrossRef] [PubMed]
- Al-Lawama, M.; Albaramki, J.; Altamimi, M.; El-Shanti, H. Congenital glucose-galactose malabsorption: A case report with a novel SLC5A1 mutation. Clin. Case Rep. 2019, 7, 51–53. [Google Scholar] [CrossRef] [PubMed]
- Kevorkova, O.; Ethier-Chiasson, M.; Lafond, J. Differential expression of glucose transporters in rabbit placenta: Effect of hypercholesterolemia in dams. Biol. Reprod. 2007, 76, 487–495. [Google Scholar] [CrossRef]
- Girard, J.; Ferre, P.; Pegorier, J.P.; Duee, P.H. Adaptations of glucose and fatty acid metabolism during perinatal period and suckling-weaning transition. Physiol. Rev. 1992, 72, 507–562. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Gu, Y.; Zhang, Y.P.; Lucas, M.J.; Wang, Y.P. High glucose levels down-regulate glucose transporter expression that correlates with increased oxidative stress in placental trophoblast cells in vitro. J. Soc. Gynecol. Investig. 2004, 11, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Joost, H.G.; Thorens, B. The extended GLUT-family of sugar/polyol transport facilitators: Nomenclature, sequence characteristics, and potential function of its novel members (review). Mol. Membr. Biol. 2001, 18, 247–256. [Google Scholar] [CrossRef]
- Wu, G.Y.; Bazer, F.W.; Satterfield, M.C.; Li, X.L.; Wang, X.Q.; Johnson, G.A.; Burghardt, R.C.; Dai, Z.L.; Wang, J.J.; Wu, Z.L. Impacts of arginine nutrition on embryonic and fetal development in mammals. Amino Acids 2013, 45, 241–256. [Google Scholar] [CrossRef]
- Freking, B.A.; Leymaster, K.A.; Vallet, J.L.; Christenson, R.K. Number of fetuses and conceptus growth throughout gestation in lines of pigs selected for ovulation rate or uterine capacity. J. Anim. Sci. 2007, 85, 2093–2103. [Google Scholar] [CrossRef]
- Fotiadis, D.; Kanai, Y.; Palacin, M. The SLC3 and SLC7 families of amino acid transporters. Mol. Asp. Med. 2013, 34, 139–158. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.F.; Chen, L.Y.; Cheng, C.S.; Chen, H.; Meng, Z.Q.; Chen, Z. SLC5A1 promotes growth and proliferation of pancreatic carcinoma via glucose-dependent AMPK/mTOR signaling. Cancer Manag. Res. 2019, 11, 3171–3185. [Google Scholar] [CrossRef] [PubMed]
- Bazer, F.W.; Kim, J.; Ka, H.; Johnson, G.A.; Wu, G.; Song, G. Select nutrients in the uterine lumen of sheep and pigs affect conceptus development. J. Reprod. Dev. 2012, 58, 180–188. [Google Scholar] [CrossRef] [PubMed]
- Pisani, L.F.; Antonini, S.; Pocar, P.; Ferrari, S.; Brevini, T.A.; Rhind, S.M.; Gandolfi, F. Effects of pre-mating nutrition on mRNA levels of developmentally relevant genes in sheep oocytes and granulosa cells. Reproduction 2008, 136, 303–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miftakhova, R.; Hedblom, A.; Semenas, J.; Robinson, B.; Simoulis, A.; Malm, J.; Rizvanov, A.; Heery, D.M.; Mongan, N.P.; Maitland, N.J.; et al. Cyclin A1 and P450 aromatase promote metastatic homing and growth of stem-like prostate cancer cells in the bone marrow. Cancer Res. 2016, 76, 2453–2464. [Google Scholar] [CrossRef] [PubMed]
- Kawamata, M.; Nishimori, K. Mice deficient in Dmrt7 show infertility with spermatogenic arrest at pachytene stage. FEBS Lett. 2006, 580, 6442–6446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sweeney, C.; Murphy, M.; Kubelka, M.; Ravnik, S.E.; Hawkins, C.F.; Wolgemuth, D.J.; Carrington, M. A distinct cyclin A is expressed in germ cells in the mouse. Development 1996, 122, 53–64. [Google Scholar] [PubMed]
- Ravnik, S.E.; Wolgemuth, D.J. Regulation of meiosis during mammalian spermatogenesis: The A-type cyclins and their associated cyclin-dependent kinases are differentially expressed in the germ-cell lineage. Dev. Biol. 1999, 207, 408–418. [Google Scholar] [CrossRef]
- Liu, D.; Matzuk, M.M.; Sung, W.K.; Guo, Q.X.; Wang, P.; Wolgemuth, D.J. Cyclin A1 is required for meiosis in the male mouse. Nat. Genet. 1998, 20, 377–380. [Google Scholar] [CrossRef]
- Liao, C.; Li, S.Q.; Wang, X.Y.; Muhlrad, S.; Bjartell, A.; Wolgemuth, D.J. Elevated levels and distinct patterns of expression of A-type cyclins and their associated cyclin-dependent kinases in male germ cell tumors. Int. J. Cancer 2004, 108, 654–664. [Google Scholar] [CrossRef]
- Fuchimoto, D.; Mizukoshi, A.; Schultz, R.M.; Sakai, S.; Aoki, F. Posttranscriptional regulation of cyclin A1 and cyclin A2 during mouse oocyte meiotic maturation and preimplantation development. Biol. Reprod. 2001, 65, 986–993. [Google Scholar] [CrossRef]
- Hipfner, D.R.; Deeley, R.G.; Cole, S.P. Structural, mechanistic and clinical aspects of MRP1. Biochim. Biophys. Acta 1999, 1461, 359–376. [Google Scholar] [CrossRef] [Green Version]
- Weaver, D.A.; Crawford, E.L.; Warner, K.A.; Elkhairi, F.; Khuder, S.A.; Willey, J.C. ABCC5, ERCC2, XPA and XRCC1 transcript abundance levels correlate with cisplatin chemoresistance in non-small cell lung cancer cell lines. Mol. Cancer 2005, 4, 18. [Google Scholar] [CrossRef]
- Wijnholds, J.; Mol, C.A.A.M.; van Deemter, L.; de Haas, M.; Scheffer, G.L.; Baas, F.; Beijnen, J.H.; Scheper, R.J.; Hatse, S.; De Clercq, E.; et al. Multidrug-resistance protein 5 is a multispecific organic anion transporter able to transport nucleotide analogs. Proc. Natl. Acad. Sci. USA 2000, 97, 7476–7481. [Google Scholar] [CrossRef] [Green Version]
- Pan, Z.; Li, S.; Liu, Q.; Wang, Z.; Zhou, Z.; Di, R.; Miao, B.; Hu, W.; Wang, X.; Hu, X.; et al. Whole-genome sequences of 89 Chinese sheep suggest role of RXFP2 in the development of unique horn phenotype as response to semi-feralization. GigaScience 2018, 7, 1–15. [Google Scholar] [CrossRef]
- Zhou, M.; Pan, Z.; Cao, X.; Guo, X.; He, X.; Sun, Q.; Di, R.; Hu, W.; Wang, X.; Zhang, X.; et al. Single nucleotide polymorphisms in the HIRA gene affect litter size in Small-Tail Han Sheep. Animals 2018, 8, 71. [Google Scholar] [CrossRef]
- Johansen, P.; Andersen, J.D.; Borsting, C.; Morling, N. Evaluation of the iPLEX (R) Sample ID Plus Panel designed for the Sequenom MassARRAY (R) system. A SNP typing assay developed for human identification and sample tracking based on the SNP for ID panel. Forensic Sci. Int. Genet. 2013, 7, 482–487. [Google Scholar] [CrossRef]
- Ortega, M.S.; Denicol, A.C.; Cole, J.B.; Null, D.J.; Hansen, P.J. Use of single nucleotide polymorphisms in candidate genes associated with daughter pregnancy rate for prediction of genetic merit for reproduction in Holstein cows. Anim. Genet. 2016, 47, 288–297. [Google Scholar] [CrossRef]
- Jiang, Z.; Michal, J.J.; Chen, J.; Daniels, T.F.; Kunej, T.; Garcia, M.D.; Gaskins, C.T.; Busboom, J.R.; Alexander, L.J.; Wright, R.W.; et al. Discovery of novel genetic networks associated with 19 economically important traits in beef cattle. Int. J. Biol. Sci. 2009, 5, 528–542. [Google Scholar] [CrossRef] [Green Version]
- Wang, N.; Li, S.; Guo, X.C.; Li, J.Y.; Ren, G.P.; Li, D.S. Fibroblast growth factor 21 improves glucose homeostasis partially via down-regulation of Na+-d-glucose cotransporter SGLT1 in the small intestine. Biomed. Pharm. 2019, 109, 1070–1077. [Google Scholar] [CrossRef]
- Lin, J.T.; Kormanec, J.; Wehner, F.; Wielert-Badt, S.; Kinne, R.K.H. High-level expression of Na+/D-glucose cotransporter (SGLT1) in a stably transfected Chinese hamster ovary cell line. Biochim. Biophys. Acta 1998, 1373, 309–320. [Google Scholar] [CrossRef]
- Yang, R.; Morosetti, R.; Koeffler, H.P. Characterization of a second human cyclin a that is highly expressed in testis and in several leukemic cell lines. Cancer Res. 1997, 57, 913–920. [Google Scholar]
- Lele, K.A.; Wolgemuth, D.J. Distinct regions of the mouse cyclin A1 gene, Ccna1, confer male germ-cell specific expression and enhancer function. Biol. Reprod. 2004, 71, 1340–1347. [Google Scholar] [CrossRef]
- Gao, X.; Sun, L.H.; Zhang, N.Y.; Li, C.; Zhang, J.C.; Xiao, Z.H.; Qi, D.S. Gestational zearalenone exposure causes reproductive and developmental toxicity in pregnant rats and female offspring. Toxins 2017, 9, 21. [Google Scholar] [CrossRef]
- Loncar, J.; Popovic, M.; Zaja, R.; Smital, T. Gene expression analysis of the ABC efflux transporters in rainbow trout (Oncorhynchus mykiss). Comp. Biochem. Physiol. C Toxicol. Pharm. 2010, 151, 209–215. [Google Scholar] [CrossRef]
- Camille Melon, L.; Maguire, J. GABAergic regulation of the HPA and HPG axes and the impact of stress on reproductive function. J. Steroid Biochem. Mol. Biol. 2016, 160, 196–203. [Google Scholar] [CrossRef]
- Hua, G.H.; Yang, L.G. A review of research progress of FecB gene in Chinese breeds of sheep. Anim. Reprod. Sci. 2009, 116, 1–9. [Google Scholar] [CrossRef]
- Wang, W.M.; Liu, S.J.; Li, F.D.; Pan, X.Y.; Li, C.; Zhang, X.X.; Ma, Y.J.; La, Y.F.; Xi, R.; Li, T.F. Polymorphisms of the ovine BMPR-IB, BMP-15 and FSHR and their associations with litter size in two Chinese indigenous sheep breeds. Int. J. Mol. Sci. 2015, 16, 11385–11397. [Google Scholar] [CrossRef]
- Davis, G.H.; Balakrishnan, L.; Ross, I.K.; Wilson, T.; Galloway, S.M.; Lumsden, B.M.; Hanrahan, J.P.; Mullen, M.; Mao, X.Z.; Wang, G.L.; et al. Investigation of the Booroola (FecB) and Inverdale (FecXI) mutations in 21 prolific breeds and strains of sheep sampled in 13 countries. Anim. Reprod. Sci. 2006, 92, 87–96. [Google Scholar] [CrossRef]
- Chu, M.X.; Wang, J.Z.; Wang, A.G.; Li, N.; Fu, J.L. Cloning and sequencing of four microsatellite loci in Small-Tail Han Sheep. Acta Genet. Sin. 2002, 29, 402–405. [Google Scholar]
- Steinhauser, C.B.; Wing, T.T.; Gao, H.; Li, X.; Burghardt, R.C.; Wu, G.; Bazer, F.W.; Johnson, G.A. Identification of appropriate reference genes for qPCR analyses of placental expression of SLC7A3 and induction of SLC5A1 in porcine endometrium. Placenta 2017, 52, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Dorniak, P.; Bazer, F.W.; Wu, G.; Spencer, T.E. Conceptus-derived prostaglandins regulate endometrial function in sheep. Biol. Reprod. 2012, 87, 1–7. [Google Scholar]
- Al-Suyufi, Y.; ALSaleem, K.; Al-Mehaidib, A.; Banemai, M.; Aldekhail, W.M.; Al-Muhandes, A.; Mohammed, M.; Allam, R.; Jambi, A.; Ramzan, K.; et al. SLC5A1 Mutations in Saudi Arabian patients with congenital glucose-galactose malabsorption. J. Pediatr. Gastroenterol. Nutr. 2018, 66, 250–252. [Google Scholar] [CrossRef]
- Atay, F.Y.; Derme, T.; Uras, N.; Ceylaner, G.; Ceylaner, S.; Sari, F.N.; Oguz, S.S. Congenital glucose-galactose malabsorption in a Turkish newborn: A novel mutation of Na+/glucose cotransporter gene. Dig. Dis. Sci. 2017, 62, 280–281. [Google Scholar] [CrossRef]
- Chen, S.X.; Wang, F.G.; Liu, Z.; Zhao, Y.; Jiang, Y.W.; Chen, L.; Li, C.J.; Zhou, X. Brain-derived neurotrophic factor promotes proliferation and progesterone synthesis in bovine granulosa cells. J. Cell. Physiol. 2019, 234, 8776–8787. [Google Scholar] [CrossRef]
- Silva-Neta, H.L.; Torrezan, E.; de Araujo Leite, J.C.; Santi-Gadelha, T.; Marques-Santos, L.F. Involvement of ABCB1 and ABCC1 transporters in sea urchin Echinometra lucunter fertilization. Mol. Reprod. Dev. 2012, 79, 861–869. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Primer Name | Primer Sequence | Product Size | Usage |
|---|---|---|---|
| SLC5A1-F1 | ACGTTGGATGTTTCAGGCTTAACCCGAGTG | 98 | PCR for g.70067210 T > C |
| SLC5A1-R1 | ACGTTGGATGGGTAAAGCATATTCCCCCAG | ||
| SLC5A1-E | TCCCCCAGAACCATTGCCTTG | Extension reaction | |
| CCNA1-F1 | ACGTTGGATGGCTTCTCTAAGGTACTGATG | 112 | PCR for g.25360220 T > C |
| CCNA1-R1 | ACGTTGGATGCACTCCCAGTCTGAAGATGC | ||
| CCNA1-E1 | ACTGATGAATTTCTTCAGCATA | Extension reaction | |
| CCNA1-F2 | ACGTTGGATGCTAGTTTGACACATCATCTC | 104 | PCR for g.25350431 C > T |
| CCNA1-R2 | ACGTTGGATGACAATTCAGTGTGGCGGTTC | ||
| CCNA1-E2 | TTTGACACATCATCTCAGTGTTA | Extension reaction | |
| ABCC1-F1 | ACGTTGGATGCTAATGTGGCCGTGTCTCTG | 125 | PCR for g.14413132 C > T |
| ABCC1-R1 | ACGTTGGATGTGCCGCCACTCATGACTATG | ||
| ABCC1-E | CACGCATGGCATCAG | Extension reaction | |
| BMPR-IB-F | GTCGCTATGGGGAAGTTTGGATG | 140 | TaqMan for A746G |
| BMPR-IB-R | CAAGATGTTTTCATGCCTCATCAACACGGTC | ||
| SLC5A1-F2 | TCAGCACAAAGTGGACAGCTC | 192 | qPCR |
| SLC5A1-R2 | CCCGGTTCCATAGGCAAACTCG | ||
| CCNA1-F3 | ACAGTTTCCCCTATGCTGGT | 164 | qPCR |
| CCNA1-R3 | TTCCTCATGTAGTGTGCCTT | ||
| ABCC1-F2 | ATGACGCATCTCAACAAAGCC | 101 | qPCR |
| ABCC1-R2 | TTGCCCATACTTCTTTCCCAG | ||
| β-Actin-F | CCAACCGTGAGAAGATGACC | 97 | qPCR |
| β-Actin-R | CCCGAGGCGTACAGGGACAG |
| Gene Name | Locus | Breed | Genotype Frequency | Allele Frequency | PIC | HE | NE | Chi-Square Test (p-Value) | |||
|---|---|---|---|---|---|---|---|---|---|---|---|
| SLC5A1 | g.70067210T > C | TT | TC | CC | T | C | |||||
| Small-Tail Han sheep | 0.72(275) | 0.20(75) | 0.08(34) | 0.81 | 0.19 | 0.26 | 0.30 | 1.43 | 0.00 | ||
| Tan sheep | 0.71(17) | 0.29(7) | 0.00(0) | 0.85 | 0.15 | 0.22 | 0.25 | 1.33 | 0.70 | ||
| Sunite sheep | 0.78 (14) | 0.22(4) | 0.00(0) | 0.89 | 0.11 | 0.18 | 0.20 | 1.25 | 0.87 | ||
| Suffolk sheep | 0.73(46) | 0.16(10) | 0.11(7) | 0.81 | 0.19 | 0.26 | 0.31 | 1.45 | 0.00 | ||
| Hu sheep | 0.72(62) | 0.21(18) | 0.07(6) | 0.83 | 0.17 | 0.25 | 0.29 | 1.40 | 0.04 | ||
| Cele black sheep | 0.64(46) | 0.23(16) | 0.13(9) | 0.76 | 0.24 | 0.30 | 0.36 | 1.57 | 0.00 | ||
| Prairie Tibetan sheep | 0.70(58) | 0.26(21) | 0.04(3) | 0.84 | 0.16 | 0.24 | 0.28 | 1.38 | 0.82 | ||
| CCNA1 | g.25350431 C > T | CC | CT | TT | C | T | |||||
| Small-Tail Han sheep | 0.02(7) | 0.24(91) | 0.74(277) | 0.14 | 0.86 | 0.21 | 0.24 | 1.32 | 0.99 | ||
| Tan sheep | 0.05(1) | 0.19(4) | 0.76(16) | 0.14 | 0.86 | 0.21 | 0.24 | 1.32 | 0.60 | ||
| Sunite sheep | 0.14(3) | 0.24(5) | 0.62(13) | 0.26 | 0.74 | 0.31 | 0.39 | 1.63 | 0.21 | ||
| Suffolk sheep | 0.00(0) | 0.13(8) | 0.87(52) | 0.07 | 0.93 | 0.12 | 0.12 | 1.14 | 0.86 | ||
| Hu sheep | 0.04(3) | 0.18 (15) | 0.78(62) | 0.13 | 0.87 | 0.20 | 0.23 | 1.30 | 0.28 | ||
| Cele black sheep | 0.03(2) | 0.22(14) | 0.75(48) | 0.14 | 0.86 | 0.21 | 0.24 | 1.32 | 0.75 | ||
| Prairie Tibetan sheep | 0.05(4) | 0.34 (27) | 0.61(48) | 0.22 | 0.78 | 0.29 | 0.34 | 1.53 | 0.99 | ||
| g.25360220 T > C | TT | CT | CC | T | C | ||||||
| Small-Tail Han sheep | 0.06(23) | 0.34(129) | 0.60(232) | 0.23 | 0.77 | 0.29 | 0.35 | 1.54 | 0.67 | ||
| Tan sheep | 0.08(2) | 0.40(10) | 0.52(12) | 0.29 | 0.71 | 0.32 | 0.41 | 1.70 | 0.99 | ||
| Sunite sheep | 0.05(0) | 0.53(10) | 0.42(8) | 0.28 | 0.72 | 0.32 | 0.40 | 1.67 | 0.26 | ||
| Suffolk sheep | 0.14(9) | 0.30(19) | 0.56(35) | 0.29 | 0.71 | 0.33 | 0.41 | 1.71 | 0.10 | ||
| Hu sheep | 0.08(7) | 0.33(28) | 0.59(51) | 0.24 | 0.76 | 0.30 | 0.37 | 1.59 | 0.55 | ||
| Cele black sheep | 0.03(2) | 0.38(27) | 0.59(41) | 0.22 | 0.78 | 0.29 | 0.34 | 1.53 | 0.61 | ||
| Prairie Tibetan sheep | 0.01(1) | 0.25(20) | 0.74(61) | 0.13 | 0.87 | 0.21 | 0.23 | 1.30 | 0.90 | ||
| ABCC1 | g.14413132 C > T | CC | CT | TT | C | T | |||||
| Small-Tail Han sheep | 0.71(270) | 0.24(94) | 0.05(20) | 0.83 | 0.17 | 0.25 | 0.29 | 1.40 | 0.01 | ||
| Tan sheep | 0.96(23) | 0.04(1) | 0.00(0) | 0.98 | 0.02 | 0.04 | 0.04 | 1.04 | 0.99 | ||
| Sunite sheep | 0.56(10) | 0.44(8) | 0.00(0) | 0.78 | 0.22 | 0.29 | 0.35 | 1.53 | 0.48 | ||
| Suffolk sheep | 0.76(48) | 0.19(12) | 0.05(3) | 0.86 | 0.14 | 0.21 | 0.24 | 1.32 | 0.21 | ||
| Hu sheep | 0.56(48) | 0.34(29) | 0.10(9) | 0.73 | 0.27 | 0.32 | 0.40 | 1.66 | 0.38 | ||
| Cele black sheep | 0.58(41) | 0.35(25) | 0.07(5) | 0.75 | 0.25 | 0.30 | 0.37 | 1.59 | 0.91 | ||
| Prairie Tibetan sheep | 0.76(62) | 0.20(17) | 0.04(3) | 0.86 | 0.14 | 0.21 | 0.24 | 1.31 | 0.43 | ||
| Gene Name | Locus | Genotype | Litter Size (Means ± S.E.) | |||
|---|---|---|---|---|---|---|
| First Parity (N) | Second Parity (N) | Third Parity (N) | Average (N) | |||
| SLC5A1 | g.70067210 T > C | TT | 1.93 ± 0.04 a (275) | 2.19 ± 0.05 a (165) | 2.59 ± 0.11 a (64) | 2.29 ± 0.07 a (275) |
| TC | 1.92 ± 0.08 a (75) | 2.16 ± 0.12 a (44) | 2.52 ± 0.21 a (18) | 1.96 ± 0.10 b (75) | ||
| CC | 1.26 ± 0.12 b (34) | 1.50 ± 0.17 b (21) | 1.70 ± 0.28 b (10) | 1.34 ± 0.11 c (34) | ||
| CCNA1 | g.25360220 C > T | TT | 1.89 ± 0.05 (23) | 2.27 ± 0.08 (9) | 2.00 ± 0.05 (4) | 1.96 ± 0.13 (23) |
| CT | 1.77 ± 0.07 (129) | 1.93 ± 0.04 (89) | 2.28 ± 0.24 (25) | 1.87 ± 0.14 (129) | ||
| CC | 1.90 ± 0.07 (232) | 2.17 ± 0.14 (160) | 2.56 ± 0.19 (59) | 1.98 ± 0.25 (232) | ||
| g.25350431 C > T | TT | 2.00 ± 0.06 (7) | 2.25 ± 0.12 (5) | 2.50 ± 0.16 (3) | 2.17 ± 0.07 (7) | |
| CT | 1.83 ± 0.06 (91) | 2.06 ± 0.09 (50) | 2.46 ± 0.18 (24) | 1.93 ± 0.27 (91) | ||
| CC | 1.88 ± 0.08 (277) | 2.12 ± 0.15 (169) | 2.46 ± 0.26 (63) | 1.96 ± 0.16 (277) | ||
| ABCC1 | g.14413132C > T | TT | 1.95 ± 0.12 (20) | 2.20 ± 0.29 (10) | 2.33 ± 0.08 (3) | 2.01 ± 0.19 (20) |
| CT | 1.88 ± 0.14 (94) | 2.17 ± 0.16 (53) | 2.52 ± 0.20 (22) | 1.99 ± 0.13 (94) | ||
| CC | 1.86 ± 0.08 (270) | 2.09 ± 0.16 (166) | 2.40 ± 0.15 (67) | 1.94 ± 0.25 (270) | ||
| BMPR-IB | c.746 A > G | BB | 2.20 ± 0.05 a (147) | 2.53 ± 0.07 a (86) | 2.82 ± 0.12 a (39) | 2.33 ± 0.04 a (147) |
| B+ | 1.88 ± 0.04 b (177) | 2.07 ± 0.06 b (110) | 2.35 ± 0.11 b (43) | 1.95 ± 0.04 b (177) | ||
| ++ | 1.03 ± 0.07 c (60) | 1.15 ± 0.11 c (33) | 1.33 ± 0.24 c (9) | 1.08 ± 0.06 c (60) | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
La, Y.; Liu, Q.; Zhang, L.; Chu, M. Single Nucleotide Polymorphisms in SLC5A1, CCNA1, and ABCC1 and the Association with Litter Size in Small-Tail Han Sheep. Animals 2019, 9, 432. https://doi.org/10.3390/ani9070432
La Y, Liu Q, Zhang L, Chu M. Single Nucleotide Polymorphisms in SLC5A1, CCNA1, and ABCC1 and the Association with Litter Size in Small-Tail Han Sheep. Animals. 2019; 9(7):432. https://doi.org/10.3390/ani9070432
Chicago/Turabian StyleLa, Yongfu, Qiuyue Liu, Liping Zhang, and Mingxing Chu. 2019. "Single Nucleotide Polymorphisms in SLC5A1, CCNA1, and ABCC1 and the Association with Litter Size in Small-Tail Han Sheep" Animals 9, no. 7: 432. https://doi.org/10.3390/ani9070432