Characterization of a Novel Porcine CSN2 Polymorphism and Its Distribution in Five European Breeds


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
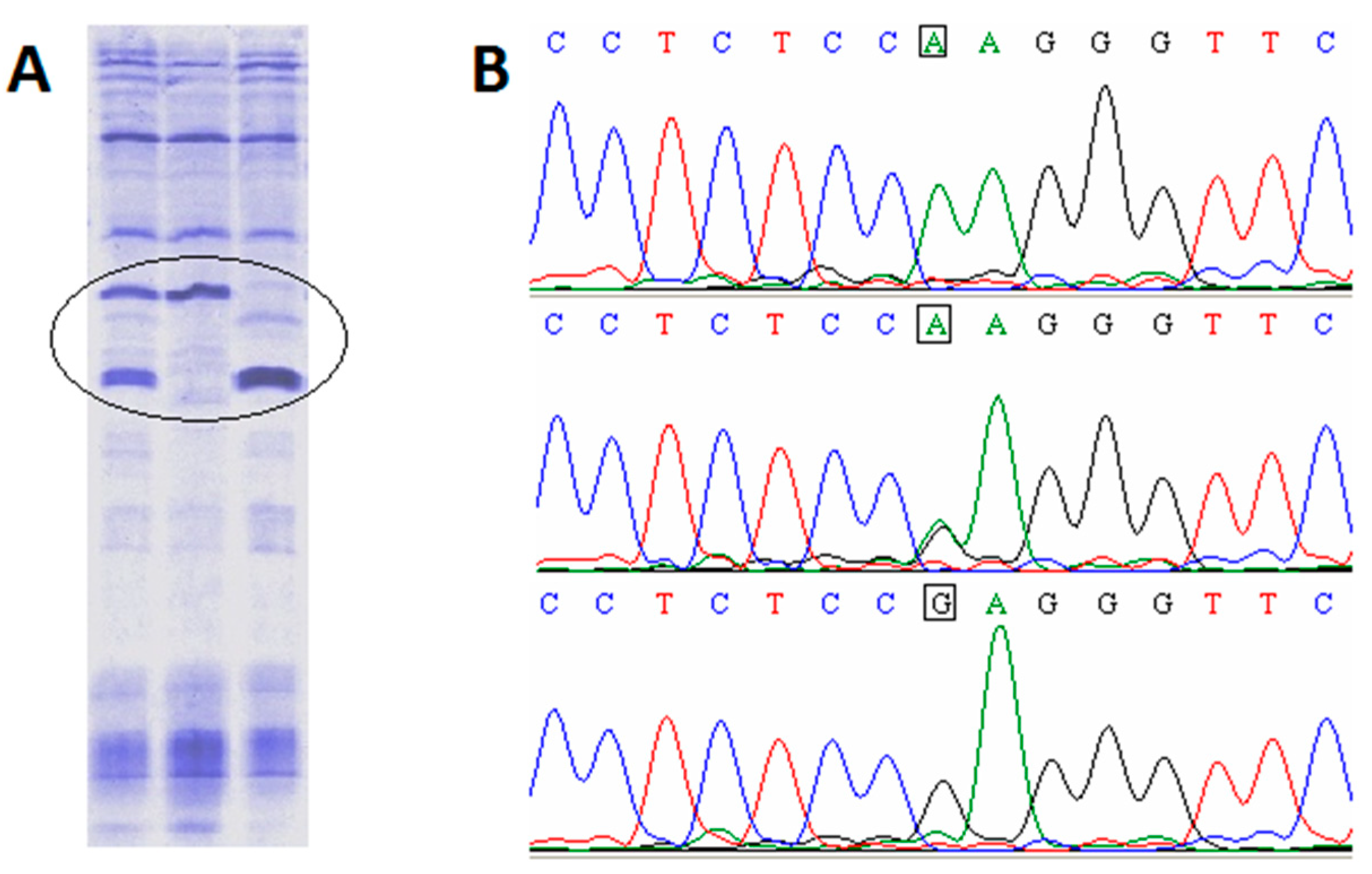
2.1. SNP Characterization
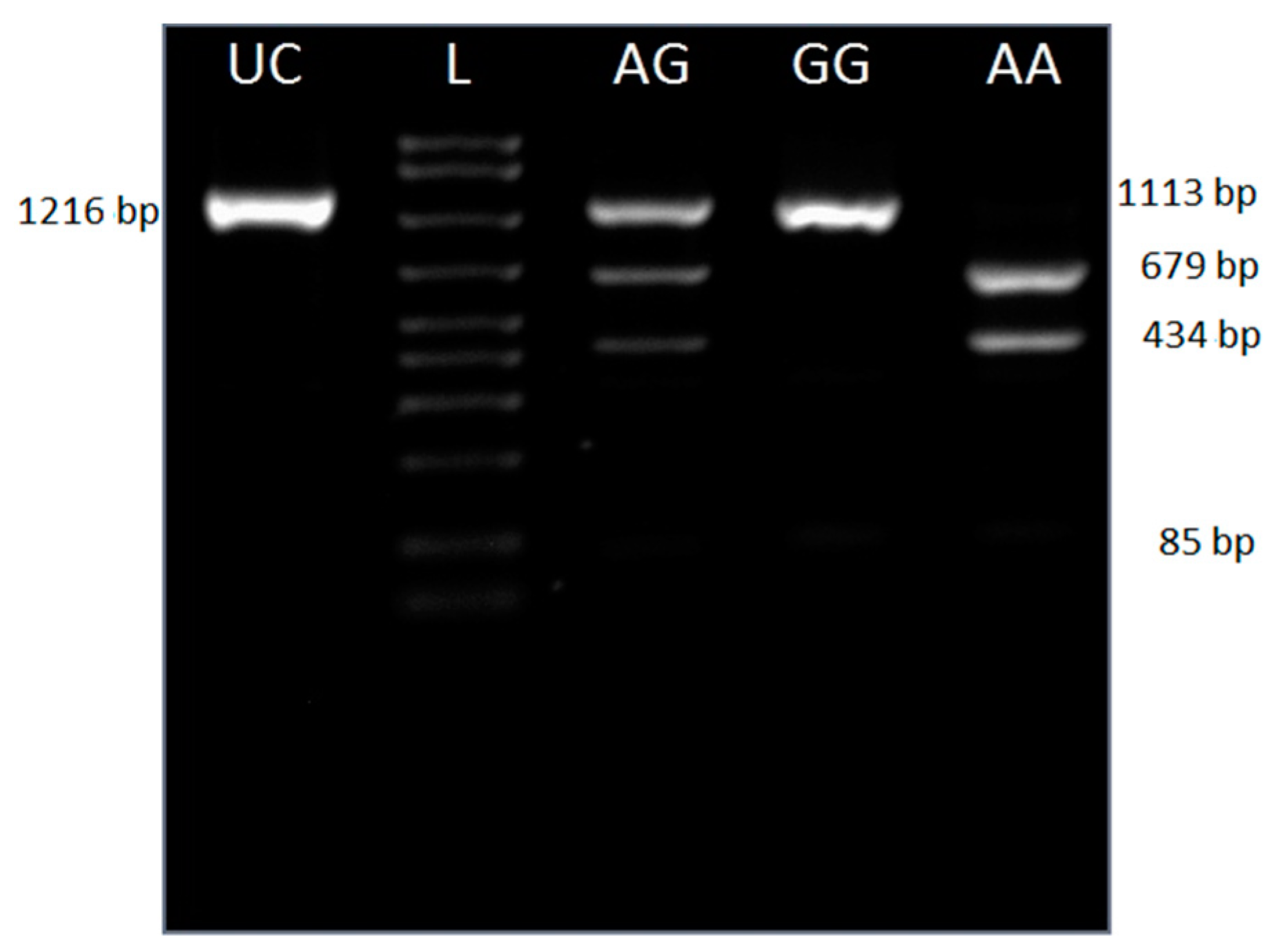
2.2. Genotyping
2.3. Statistical Analysis
3. Results
3.1. SNP Characterization
3.2. Genotyping
3.3. Statistical Analysis
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Aimutis, W.R.; Kornegay, E.T.; Eigel, W.N. Electrophoretic and biochemical comparison of casein and whey protein from porcine colostrum and milk. J. Dairy Sci. 1982, 65, 1874–1881. [Google Scholar] [CrossRef]
- Lee, P.; Chung, H.K.; Lee, H.C.; Woo, J.S.; Lee, S.; Jo, S.J.; Chang, W.K.; Lee, H.T.; Kwon, M.; Park, J.K. Cloning and characterization of 5’-untranslated region of porcine beta casein gene (CSN2). Domest. Anim. Endocrinol. 2008, 35, 245–253. [Google Scholar] [CrossRef] [PubMed]
- Gallagher, D.P.; Cotter, P.F.; Mulvihill, D.M. Porcine milk proteins: A review. Int. Dairy J. 1997, 7, 99–118. [Google Scholar] [CrossRef]
- Martin, P.; Bianchi, L.; Cebo, C.; Miranda, G. Genetic polymorphism of milk proteins. In Advanced Dairy Chemistry; McSweeney, P., Fox, P., Eds.; Springer: Boston, MA, USA, 2013. [Google Scholar]
- Alexander, L.J.; Beattie, C.W. The sequence of porcine β-casein cDNA. Anim. Genet. 1992, 23, 369–371. [Google Scholar] [CrossRef] [PubMed]
- Archibald, A.L.; Couperwhite, S.; Haley, C.S.; Beattie, C.W.; Alexander, L.J. RFLP and linkage analysis of the porcine casein loci—CASAS1, CASAS2, CASB and CASK. Anim. Genet. 1994, 25, 349–351. [Google Scholar] [CrossRef]
- Șuteu, M.; Vlaic, A.; Reneville, R. Characterization of porcine β-casein G allele (CSN2G). ABAH Bioflux 2011, 3, 105–109. [Google Scholar]
- Cieslak, J.; Mankowska, M.; Switonski, M. Between-breed variation in frequency of five novel missense SNPs in porcine casein beta (CSN2) and casein kappa (CSN3) genes. Anim. Genet. 2012, 43, 363–364. [Google Scholar] [CrossRef]
- Șuteu, M.; Vlaic, A.; Bâlteanu, V.A.; Wavreille, J.; Renaville, R. Evidence of alternative splicing of porcine β-casein (CSN2). Anim. Genet. 2012, 43, 474–475. [Google Scholar] [CrossRef]
- Șuteu, M.; Vlaic, A.; Renaville, R. A 44 amino acid porcine β-casein isoform. Porcine Res. 2012, 2, 1–3. [Google Scholar]
- Lee, S.M.; Kim, H.M.; Moon, S.J.; Kang, M.J. Cloning and molecular characterization of porcine β-casein gene (CNS2). Asian Australas. J. Anim. Sci. 2012, 25, 421–427. [Google Scholar] [CrossRef]
- Șuteu, M.; Vlaic, A.; Bâlteanu, V.; Pop, F. Porcine milk protein polymorphisms detection by means of isoelectric focusing. Bull. UASVM Anim. Sci. Biotechnol. 2009, 66, 146–151. [Google Scholar]
- RestrictionMapper. Available online: http://www.restrictionmapper.org/ (accessed on 13 May 2019).
- Yeh, F.C.; Boyle, T.J.B. Population genetic analysis of co-dominant and dominant markers and quantitative traits. Belg. J. Bot. 1997, 129, 157–163. [Google Scholar]
- Selvaggi, M.; Pesce Delfino, A.R.; Dario, C. Exon 1 polymorphisms in the equine CSN3 gene: SNPs distribution analysis in Murgese horse breed. Anim. Biotechnol. 2010, 21, 252–256. [Google Scholar] [CrossRef] [PubMed]
- Vlaic, A. Proposal for the nomenclature of porcine (Sus scrofa) β-casein alleles. Bull. UASVM Anim. Sci. Biotechnol. 2012, 69, 299–300. [Google Scholar]
- Ciobanu, D.C.; Day, A.E.; Nagy, A.; Wales, R.; Rothschild, M.F.; Plastow, G.S. Genetic variation in two conserved local Romanian pig breeds using type 1 DNA markers. Genet. Sel. Evol. 2001, 33, 417–432. [Google Scholar] [CrossRef] [PubMed]
- Dornea, M.M.; Mateoc-Sirb, N.; Gorun, L.M.; Raicov, M. Study of rare traditional pork breeds concerning the aspect of biodiversity conservation. Bull. UASVM Anim. Sci. Biotechnol. 2012, 69, 308–310. [Google Scholar]
- Zăhan, M.; Raica, P.; Miclea, V.; Miclea, I.; Renaville, R.; Duterme, O.; Mihaiescu, M.; Nagy, A. Results concerning genetic characterization of Mangalita breed using microsatellite markers. Sci. Pap. Anim. Sci. Biotechnol. 2009, 42, 136–140. [Google Scholar]
- Mahan, D.C.; Lepine, A.J. Effect of pig weaning weight and associated nursery feeding programs on subsequent performance to 105 kilograms body weight. J. Anim. Sci. 1991, 69, 1370–1378. [Google Scholar] [CrossRef]
- Wolter, B.F.; Ellis, M. The effects of weaning weight and rate of growth immediately after weaning on subsequent pig growth performance and carcass characteristics. Can. J. Anim. Sci. 2001, 81, 363–369. [Google Scholar] [CrossRef]
- Farmer, C.; Charagu, P.; Palin, M.F. Influence of genotype on metabolic variables, colostrum and milk composition of primiparous sows. Can. J. Anim. Sci. 2007, 87, 511–515. [Google Scholar] [CrossRef]
- Skrzypczak, E.; Babicz, M.; Szulc, K.; Walendowska, A.; Panek, A. Impact of sow milk protein polymorphism on piglet rearing. Ann. Anim. Sci. 2012, 12, 335–347. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Breed | N | Number of Sows | Genotype Frequency | Gene Frequency | |||||
|---|---|---|---|---|---|---|---|---|---|
| AA | AG | GG | PAA | HAG | QGG | pA | qG | ||
| Landrace | 67 | 0 | 3 | 64 | 0 | 0.07 | 0.93 | 0.035 | 0.965 |
| Pietrain | 40 | 0 | 0 | 40 | 0 | 0 | 1 | 0 | 1 |
| Large White | 36 | 2 | 17 | 17 | 0.06 | 0.47 | 0.47 | 0.295 | 0.705 |
| Mangalita | 13 | 0 | 3 | 10 | 0 | 0.23 | 0.77 | 0.115 | 0.885 |
| Bazna | 11 | 1 | 8 | 2 | 0.09 | 0.73 | 0.18 | 0.445 | 0.545 |
| Breed | Gene Homozygosity (Ho) | Gene Heterozygosity (He) | Effective Number of Alleles (Ne) | Fixation Index (FIS) | Hardy-Weinberg Equilibrium (χ2) |
|---|---|---|---|---|---|
| Landrace | 0.9552 | 0.0448 | 1.0458 | −0.0229 | 0.0232 |
| Large White | 0.5278 | 0.4722 | 1.7041 | −0.1429 | 0.6044 |
| Mangalita | 0.2727 | 0.7273 | 1.9836 | −0.4667 | 1.9394 |
| Bazna | 0.7692 | 0.2308 | 1.2565 | −0.1304 | 0.1423 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Șuteu, M.; Vlaic, A.; Dărăban, S.V. Characterization of a Novel Porcine CSN2 Polymorphism and Its Distribution in Five European Breeds. Animals 2019, 9, 419. https://doi.org/10.3390/ani9070419
Șuteu M, Vlaic A, Dărăban SV. Characterization of a Novel Porcine CSN2 Polymorphism and Its Distribution in Five European Breeds. Animals. 2019; 9(7):419. https://doi.org/10.3390/ani9070419
Chicago/Turabian StyleȘuteu, Mihai, Augustin Vlaic, and Stelian Vasile Dărăban. 2019. "Characterization of a Novel Porcine CSN2 Polymorphism and Its Distribution in Five European Breeds" Animals 9, no. 7: 419. https://doi.org/10.3390/ani9070419