Differences in Pre-Laying Behavior between Floor-Laying and Nest-Laying Pekin Ducks


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
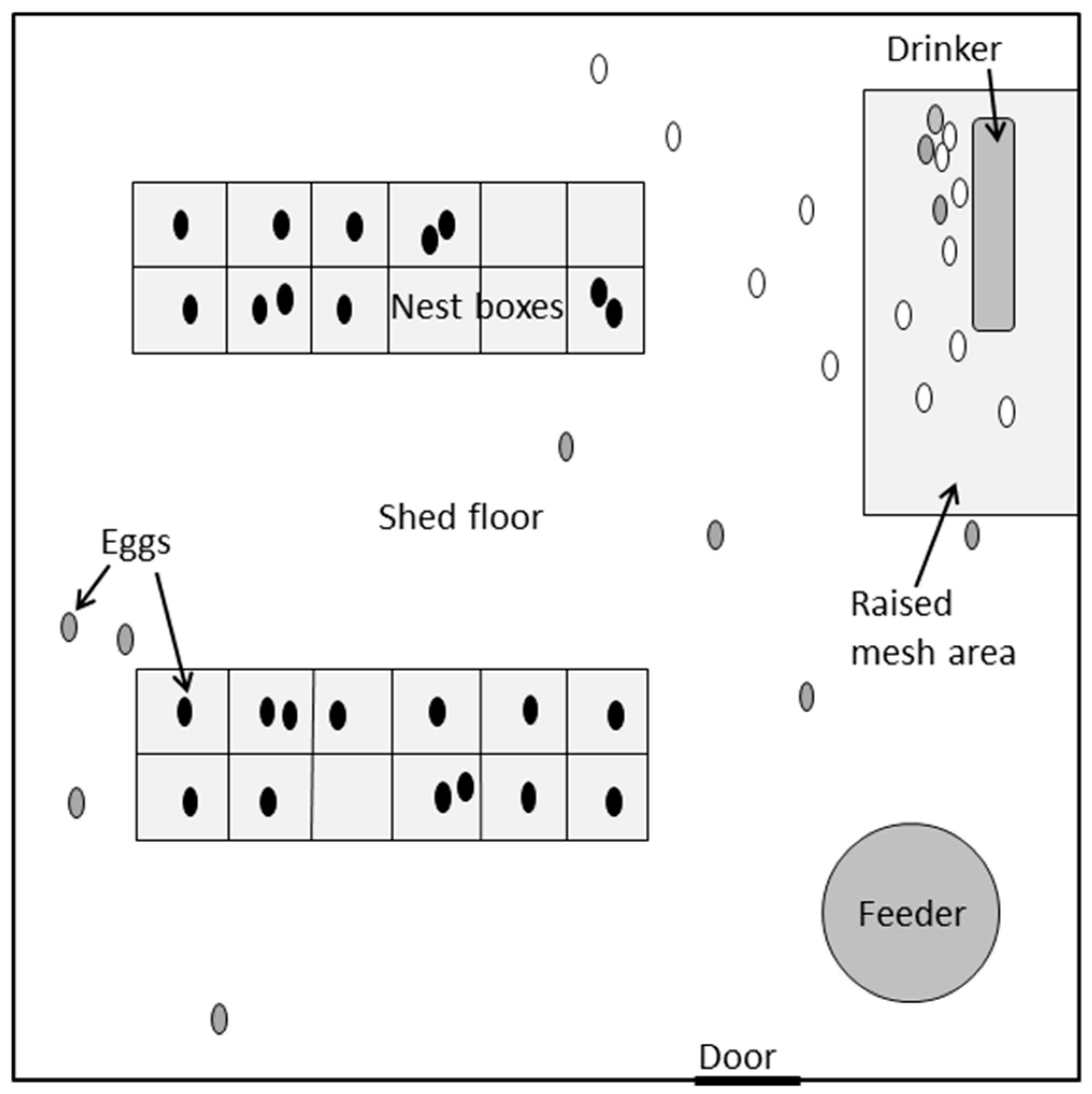
2.1. Animals and Husbandry
2.2. Video Recording and Quantification of Pre-Laying Behavior
2.3. Statistical Analysis
3. Results
3.1. Nest Box Interactions
3.2. Nest-Building Behavior
3.3. Behavior Outside of Nest Boxes
3.4. Behavior Inside Nest Boxes
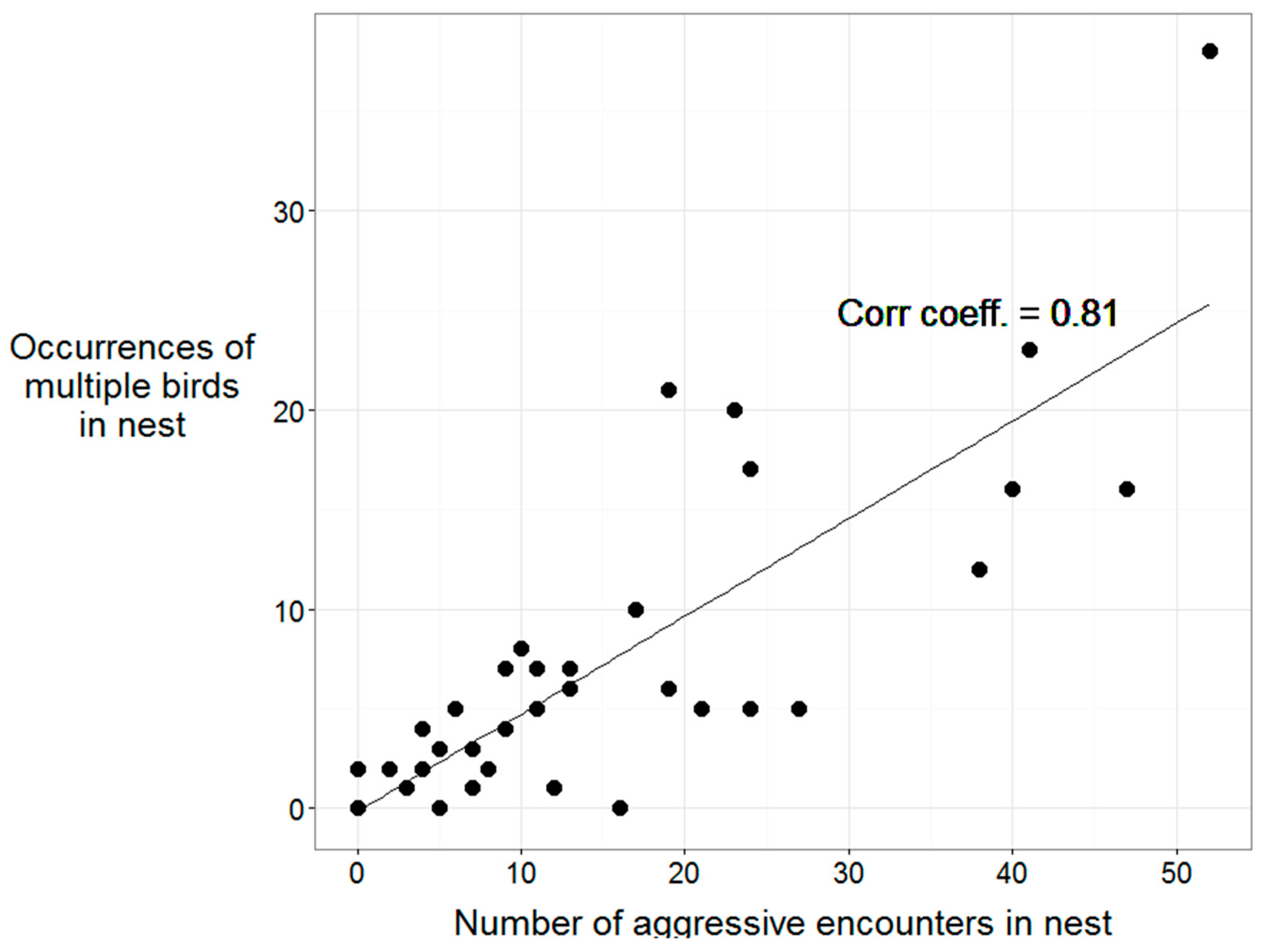
3.5. Aggressive Interactions
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cherry, P.P.; Morris, T.R. Domestic Duck Production: Science and Practice; CABI: Wallingford, UK, 2008; pp. 205–209. [Google Scholar]
- McLachlan, G.; (Pepe’s Ducks Ltd., Sydney, NSW, Australia). Personal communication, 2013.
- Appleby, M.C.; Mench, J.A.; Hughes, B.O. Poultry Behaviour and Welfare; CABI: Wallingford, UK, 2004; pp. 91–117. [Google Scholar]
- Mench, J.A. Behaviour of fowl and other domesticated birds. In The Ethology of Domestic Animals; Jensen, P., Ed.; CABI: Wallingord, UK, 2009; pp. 121–136. [Google Scholar]
- Mason, G.; Bateson, M. Motivation and the organisation of behaviour. In The Ethology of Domestic Animals; Jensen, P., Ed.; CABI: Wallingord, UK, 2009; pp. 38–56. [Google Scholar]
- Lokemoen, J.T.; Duebbert, H.F.; Sharp, D.E. Nest spacing, habitat selection, and behavior of waterfowl on Miller Lake Island, North Dakota. J. Wildl. Manag. 1984, 48, 309–321. [Google Scholar] [CrossRef]
- Makagon, M.M.; Tucker, C.B.; Mench, J.A. Factors affecting nest choice by Pekin ducks. Appl. Anim. Behav. Sci. 2011, 129, 121–128. [Google Scholar] [CrossRef]
- Cooper, J.J.; Appleby, M.C. Individual variation in prelaying behaviour and the incidence of floor eggs. Br. Poult. Sci. 1996, 37, 245–253. [Google Scholar] [CrossRef]
- Guinebretière, M.; Huneau-Salaun, A.; Hunnic, D.; Michel, V. Cage hygiene, laying location, and egg quality: The effects of linings and litter provision in furnished cages for laying hens. Poult. Sci. 2012, 91, 808–816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stämpfli, K.; Buchwalder, T.; Frohlich, E.K.F.; Roth, B.A. Design of nest access grids and perches in front of the nests: Influence on the behavior of laying hens. Poult. Sci. 2013, 92, 890–899. [Google Scholar] [CrossRef] [Green Version]
- Cooper, J.J.; Appleby, M.C. Nesting behaviour of hens: Effects of experience on motivation. Appl. Anim. Behav. Sci. 1995, 42, 283–295. [Google Scholar] [CrossRef]
- Sherwin, C.M.; Nicol, C.J. Factors influencing floor-laying by hens in modified cages. Appl. Anim. Behav. Sci. 1993, 36, 211–222. [Google Scholar] [CrossRef]
- Hunniford, M.E.; Widowski, T.M. Rearing environment and laying location affect pre-laying behaviour in enriched cages. Appl. Anim. Behav. Sci. 2016, 181, 205–213. [Google Scholar] [CrossRef]
- Wall, H. Production performance and proportion of nest eggs in layer hybrids housed in different designs of furnished cages. Poult. Sci. 2011, 90, 2153–2161. [Google Scholar] [CrossRef] [Green Version]
- Guesdon, V.; Faure, J.M. Laying performance and egg quality in hens kept in standard or furnished cages. Anim. Res. 2004, 53, 45–57. [Google Scholar] [CrossRef] [Green Version]
- Makagon, M.M.; Mench, J.A. Floor laying by Pekin ducks: Effects of nest box ratio and design. Poult. Sci. 2011, 90, 1179–1184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freire, R.; Appleby, M.C.; Hughes, B.O. Effects of social interactions on pre-laying behaviour in hens. Appl. Anim. Behav. Sci. 1998, 56, 47–57. [Google Scholar] [CrossRef]
- McBride, A.; (Luv-a-Duck Ltd., Nhil, Vic, Australia). Personal communication, 2013.
- Malecki, I.A.; School of Animal Biology, The University of Western Australia, Perth, WA, Australia. Unpublished Data, 2014.
- Martin, P.; Bateson, P. Measuring Behaviour: An Introductory Guide; Cambridge University Press: Cambridge, UK, 1986; pp. 121–134. [Google Scholar]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- Rietveld-Piepers, B.; Blokhuis, H.J.; Wiepkema, P.R. Egg-laying behaviour and nest-site selection of domestic hens kept in small floor-pens. Appl. Anim. Behav. Sci. 1985, 14, 75–88. [Google Scholar] [CrossRef]
- Freire, R.; Appleby, M.C.; Hughes, B.O. Assessment of pre-laying motivation in the domestic hen using social interaction. Anim. Behav. 1997, 54, 313–319. [Google Scholar] [CrossRef]
- Pagel, M.; Dawkins, M. Peck orders and group size in laying hens: Future contracts for non-aggresion. Behav. Proc. 1997, 40, 13–25. [Google Scholar] [CrossRef]
- Price, E.O. Animal Domestication and Behavior; CABI Publishing: Wallingford, UK, 2002; pp. 73–82. [Google Scholar]
- Blem, C.R. Energy balance. In Sturkie’s Avian Physiology; Whittow, C., Ed.; Academic Press: San Diego, CA, USA, 2000; pp. 327–343. [Google Scholar]
- Lundberg, A.; Keeling, L.J. The impact of social factors on nesting in laying hens (Gallus gallus domesticus). Appl. Anim. Behav. Sci. 1999, 64, 57–69. [Google Scholar] [CrossRef]
- Kite, V.G.; Cumming, R.B.; Wodzicka-Tomaszewska, M. Nesting behaviour of hens in relation to the problem of floor eggs. In Proceedings of the Symposium Behaviour in Relation to Reproduction, Management and Welfare of Farm Animals, Armidale, NSW, Australia, September 1979. [Google Scholar]
- Riber, A.B. Development with age of nest box use and gregarious nesting in laying hens. Appl. Anim. Behav. Sci. 2010, 123, 24–31. [Google Scholar] [CrossRef]
- Millam, J.R. Preference of turkey hens for nest-boxes of different levels of interior illimination. Appl. Anim. Behav. Sci. 1987, 18, 341–348. [Google Scholar] [CrossRef]
- Schmid, I.; Wechsler, B. Behaviour of Japanese quail (Coturnix japonica) kept in semi-natural aviaries. Appl. Anim. Behav. Sci. 1997, 55, 103–112. [Google Scholar] [CrossRef]
- Makagon, M.M. Nest Site Selection by Pekin Ducks. Ph.D. Thesis, University of California, Davis, CA, USA, 2010. [Google Scholar]
- Denk, A.G. Male and Female Reproductive Tactics in Mallards (Anas platyrhynchos L.): Sperm Competition and Cryptic Female Choice. Ph.D. Thesis, LMU Munich, Munich, Germany, 2005. [Google Scholar]
- Kreisinger, J.; Munclinger, P.; Javurkova, V.; Albrecgt, T. Analysis of extra-pair paternity and conspecific brood parasitism in Mallards Anas platyrhynchos using non-invasive techniques. J. Avian Biol. 2010, 41, 551–557. [Google Scholar] [CrossRef]
- Huber, H.U.; Folsch, D.W.; Stahli, U. Influence of Various Nesting Materials on Nest Site Selection of the Domestic Hen. Br. Poult. Sci. 1985, 26, 367–373. [Google Scholar] [CrossRef] [PubMed]
- Struelens, E.; Tuyttens, F.A.M.; Janssen, A.; Leroy, T.; Audoorn, L.; Vranken, E.; De Baere, K.; Odberg, F.; Berckmans, D.; Zoons, J.; et al. Design of laying nests in furnished cages: Influence of nesting material, nest box position and seclusion. Br. Poult. Sci. 2005, 46, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Struelens, E.; Van Nuffel, A.; Tuyttens, F.A.M.; Audoorn, L.; Vranken, E.; Zoons, J.; Berckmans, D.; Odber, F.; Van Dongen, S.; Sonck, B. Influence of nest seclusion and nesting material in pre-laying behaviour of laying hens. Appl. Anim. Behav. Sci. 2008, 112, 106–119. [Google Scholar] [CrossRef]
- Meijsser, F.M.; Hughes, B.O. Comparative analysis of pre-laying behaviour in battery cages and in three alternative systems. Br. Poult. Sci. 1989, 30, 747–760. [Google Scholar] [CrossRef]
- Cronin, G.M.; Barnett, J.; Storey, T.H.; Thomson, P.C.; Hemsworth, P.H. The relationship between pre-laying activity and corticosterone concentrations, and the interpretation for laying hen welfare. In Proceedings of the 23rd Annual Australian Poultry Science Symposium, Sydney, Australia, 19–22 February 2012. [Google Scholar]
- Brummelmann, M.; (Wagin Duck & Game, Wagin, WA, Australia). Personal communication, 2014.
- Hansen, S.; af Hagelsrum, D.; Louise, J. Emergence of displacement activites in the male rat following thwartingof sexual behavior. Behav. Neurosci. 1984, 98, 868–883. [Google Scholar] [CrossRef]
- Duncan, I.J.H.; Wood-Gush, D.G.M. Thwarting of feeding behaviour in the domestiv fowl. Anim. Behav. 1972, 20, 444–451. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Behavior | Description |
|---|---|
| Primary Observations | |
| States: | |
| Walking | Duck is outside of the nest box, moving around the shed floor. |
| Sitting | Duck is sitting down quietly with wings tucked in, either awake or asleep. Occurs either inside or outside of the nest box. |
| Maintenance behavior | Duck is engaged in either feeding, drinking or preening. Each of these is further defined as: duck is standing at the feeder actively engaged in bouts of food consumption; duck is standing at the drinker, actively engaged in bouts of drinking; duck is engaged in self-directed grooming/cleaning of body. |
| Nest-building behavior | Duck is using body and feet to create a hollow in the substrate and/or manipulating substrate with beak in the nest hollow. Occurs either outside nest boxes on the floor, or inside a nest box. |
| Events: | |
| Nest box entry | Duck places full body inside a nest box. |
| Nest box exit | Duck places full body outside a nest box. |
| Nest box investigation | Duck stands outside of nest box with neck extended and head inside box, may or may not engage with box substrate. |
| Multiple birds in nest box | Subject bird plus 1 or more birds with full bodies in the nest box concurrently. |
| Aggressive behavior | Subject duck is engaged with other bird/s, overt aggression (e.g., grabbing neck skin, feather pecking, chasing) is seen. Subject either receives or initiates aggression. |
| Non-aggressive behavior | Subject duck is engaged with other bird/s (e.g., sitting in physical contact with another bird, permitting or performing investigative behavior from/towards another bird as passing by), but no overt aggression is seen. Subject either receives or initiates non-aggressive behavior. |
| Secondary Variables | |
| Time in nest boxes | Duration of nest box visits, calculated between occurrences of nest box entries and exits. |
| Aggression in nest box | Event where subject duck was concurrently in the nest box and involved in aggressive encounter. Subject either receives or initiates aggression. |
| Nest box exit due to aggression | Event where a duck involved in an aggressive encounter co-occurred with that duck exiting the nest box. Subject may either be initiating or receiving aggression. |
| Behavior | FL-Out (n = 13) | FL-In (n = 11) | NL (n = 24) | |||
|---|---|---|---|---|---|---|
| Median | (Min–Max) | Median | (Min–Max) | Median | (Min–Max) | |
| Nest box entries | ||||||
| Frequency | 0 | (0–0) | 10 | (2–15) | 7 | (1–26) |
| Percent time | - | - | 46.45 | (29.5–78.2) | 47.29 | (5.19–86.9) |
| Duration | - | - | 142.1 | (96.5–311.1) | 149.6 | (374.0–1535.0) |
| Nest box investigations | ||||||
| Frequency | 0 | (0–0) | 4 | (0–28) | 9 | (0–30) |
| Percent time | - | - | 0.3 | (0.0–3.4) | 0.9 | (0.1–7.5) |
| Durations (s) | - | - | 2.0 | (0.0–7.2) | 3.0 | (1.4–9.0) |
| Nest-building | ||||||
| Frequency | 8 | (1–24) | 7 | (3–12) | 6 | (0–23) |
| Percent time | 10.8 | (1.4–13.0) | 6.5 | (1.6–9.2) | 3.9 | (0.0–14.1) |
| Duration (s) | 31.9 | (15.5–89.4) | 31.2 | (19.5–49.0) | 22.3 | (0.0–63.8) |
| Walking | ||||||
| Frequency | 25 | (2–91) | 26 | (10–57) | 19 | (3–103) |
| Percent time | 2.8 ab | (0.4–13.4) | 21.1 a | (8.5–47.0) | 13.7 b | (1.1–33.5) |
| Duration (s) | 7.9 | (6.8–9.3) | 5.9 | (3.8–11.0) | 6.8 | (3.6–10.0) |
| Sitting outside box | ||||||
| Frequency | 11 a | (7–28) | 0 b | (0–4) | 2.5 b | (0–14) |
| Percent time | 61.5 a | (22.4–86.0) | 0.0 b | (0.0–38.7) | 19.5 b | (0.0–71.0) |
| Durations (s) | 185.1 a | (57.1–328.6) | 0.0 b | (0.0–200.1) | 80.4 ab | (0.0–391.6) |
| Sitting inside box | ||||||
| Frequency | - | - | 7 | (0–19) | 7 | (0–23) |
| Percent time | - | - | 34.9 * | (9.8–66.3) | 50.6 * | (0.0–86.8) |
| Duration (s) | - | - | 81.3 | (51.9–198.7) | 115.0 | (77.4–390.1) |
| Aggressive interactions | ||||||
| Frequency | 6 ab | (2–44) | 35 a | (12–87) | 30 b | (2–112) |
| Percent time | 0.61 ab | (0.0–4.5) | 5.6 a | (2.2–11.2) | 4.6 b | (0.6–14.1) |
| Duration (s) | 2.8 | (0.0–6.2) | 14 a | (2–43) | 10 b | (0–52) |
| Number initiated | 1 ab | (0–27) | 14 a | (2–43) | 10 b | (0–52) |
| Number received | 5 ab | (2–33) | 21 a | (10–44) | 24.5 b | (2–85) |
| % Total aggressive encounters occurring in nest box | - | - | 44.4 | (25.0–93.6) | 33.9 | (0.0–83.3) |
| % Total aggression occurring in nest box | - | - | 5.6 | (2.1–9.7) | 7.6 | (1.0–15.1) |
| % Time in aggression when in nest box | - | - | 52.5 | (28.9–94.1) | 53.1 | (16.0–89.0) |
| Aggression associated nest exits | ||||||
| Total | - | - | 7 | (1–10) | 4.5 | (0–27) |
| Aggression initiated | - | - | 2.5 | (1–4) | 2 | (1–5) |
| Aggression received | - | - | 5 | (2–9) | 5 | (1–22) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barrett, L.; Malecki, I.; Blache, D. Differences in Pre-Laying Behavior between Floor-Laying and Nest-Laying Pekin Ducks. Animals 2019, 9, 40. https://doi.org/10.3390/ani9020040
Barrett L, Malecki I, Blache D. Differences in Pre-Laying Behavior between Floor-Laying and Nest-Laying Pekin Ducks. Animals. 2019; 9(2):40. https://doi.org/10.3390/ani9020040
Chicago/Turabian StyleBarrett, Lorelle, Irek Malecki, and Dominique Blache. 2019. "Differences in Pre-Laying Behavior between Floor-Laying and Nest-Laying Pekin Ducks" Animals 9, no. 2: 40. https://doi.org/10.3390/ani9020040