Dietary Energy Levels Affect Growth Performance through Growth Hormone and Insulin-Like Growth Factor 1 in Yak (Bos grunniens)

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Diets
2.2. Experimental Procedure and Analytical Methods
2.3. Sample Collection and Analysis
2.4. RNA Extraction and Quantitative Real-time PCR
2.5. Statistical Analysis
3. Results
3.1. Effects of Dietary Energy Levels on Growth Performance of Yak
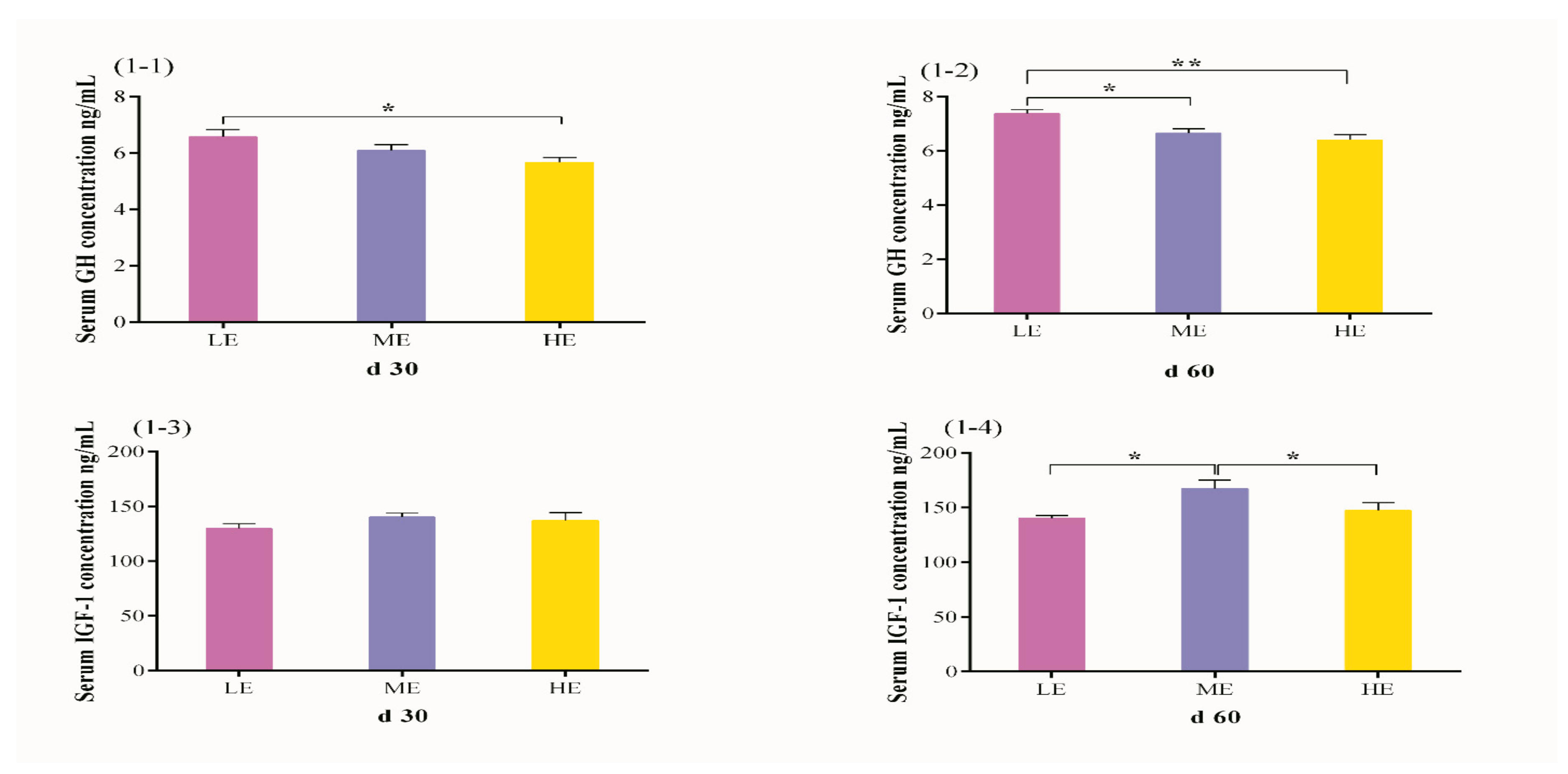
3.2. Effects of Dietary Energy Levels on Serum GH and IGF-1 Concentrations in Yak
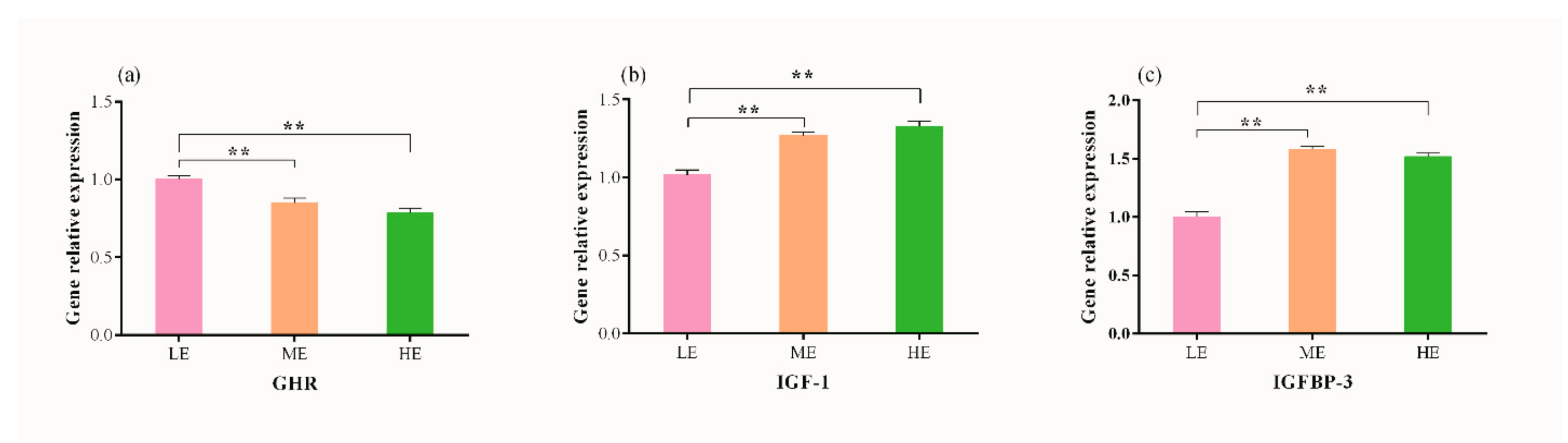
3.3. Effects of Dietary Energy Levels on Hepatic mRNA Abundance of GHR, IGF-1 and IGFBP-3 in Yaks
3.4. Correlation Analysis between Serum Hormone Levels and Expression Levels of Genes on Growth Performance of Yak
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ding, X.Z.; Guo, X.; Yan, P.; Liang, C.N.; Bao, P.J.; Chu, M. Seasonal and nutrients intake regulation of lipoprotein lipase (LPL) activity in grazing yak (Bos grunniens) in the Alpine Regions around Qinghai Lake. Livest. Sci. 2012, 143, 29–34. [Google Scholar] [CrossRef]
- Long, R.J.; Ding, L.M.; Shang, Z.H.; Guo, X.H. The yak grazing system on the Qinghai-Tibetan plateau and its status. Rangel. J. 2008, 30, 241–246. [Google Scholar] [CrossRef]
- Barreto, L.M.G.; Medeiros, A.N.D.; Ângela, M.V.B.; Araújo, G.G.L.D.; Furtado, D.A.; Lisboa, A.C.C.; Paulo, J.L.A.; Alves, A.R. Growth performance of native goats fed diets containing different levels of energy. Rev. Bras. Zootecn. 2012, 41, 675–682. [Google Scholar] [CrossRef] [Green Version]
- Haddad, S.G.; Husein, M.Q. Effect of dietary energy density on growth performance and slaughtering characteristics of fattening awassi lambs. Livest. Prod. Sci. 2004, 87, 171–177. [Google Scholar] [CrossRef]
- Radunz, A.E.; Fluharty, F.L.; Day, M.L.; Zerby, H.N.; Loerch, S.C. Prepartum dietary energy source fed to beef cows: I. effects on pre- and postpartum cow performance. J. Anim. Sci. 2010, 88, 2717–2728. [Google Scholar] [CrossRef] [PubMed]
- Peng, Q.H.; Wang, Z.S.; Tan, C.; Zhang, H.B.; Hu, Y.N.; Zou, H.W. Effects of different pomace and pulp dietary energy density on growth performance and intramuscular fat deposition relating mRNA expression in beef cattle. J. Food Agric. Environ. 2012, 10, 404–407. [Google Scholar]
- Beauchemin, K.A.; Mcclelland, L.A.; Sdm, J.; Kozub, G.C. Effects of crude protein content, protein degradability and energy concentration of the diet on growth and carcass characteristics of market lambs fed high concentrate diets. Can. J. Anim. Sci. 1995, 75, 387–395. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.; Liu, J.B.; Wu, X.Y.; Bao, P.J.; Long, R.J.; Guo, X.; Ding, X.Z.; Yan, P. The response of gene expression associated with lipid metabolism, fat deposition and fatty acid profile in the longissimus dorsi muscle of Gannan yaks to different energy levels of diets. PLoS ONE 2017, 12, e0187604. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, X.; Wang, Z.; Dong, X.; Tan, C.; Zou, H.; Peng, Q.; Xue, B.; Wang, L.; Dong, G. Effects of dietary energy level on lipid metabolism-related gene expression in subcutaneous adipose tissue of Yellow breed × Simmental cattle. Anim. Sci. J. 2015, 86, 392–400. [Google Scholar] [CrossRef]
- Sørensen, M.T.; Chaudhuri, S.; Louveau, I.; Coleman, M.E.; Etherton, T.D. Growth hormone binding proteins in pig adipose tissue: Number, size and effects of pGH treatment on pGH and bGH binding. Domest. Anim. Endocrinol. 1992, 9, 13–24. [Google Scholar] [CrossRef]
- Baumann, G. Growth hormone-binding proteins: State of the art. J. Endocrinol. 1994, 141, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Renaville, R.; Hammadi, M.; Portetelle, D. Role of the somatotropic axis in the mammalian metabolism. Domest. Anim. Endocrinol. 2002, 23, 351–360. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.; Xu, X.R.; Ren, H.Y.; Zhang, Y.H.; Xv, S.Z. The effects of the GH, IGF-I and IGFIBP-3 gene on growth and development traits of nanyang cattle in different growth period. Hereditas 2006, 28, 927–932. [Google Scholar] [PubMed]
- Othman, O.E.; Alam, S.S.; El-Aziem, S.H.A. Single nucleotide polymorphism in Egyptian cattle insulin-like growth factor binding protein-3 gene. J. Genet. Eng. Biotechnol. 2014, 12, 143–147. [Google Scholar] [CrossRef] [Green Version]
- Ayuk, J.; Sheppard, M.C. Growth hormone and its disorders. Postgrad. Med. J. 2006, 82, 24–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akis, I.; Oztabak, K.; Gonulalp, I.; Meng, A.; Un, C. IGF-1 and IGF-1R gene polymorphisms in East Anatolian Red and South Anatolian Red cattle breeds. Russ. J. Genet. 2010, 46, 439–442. [Google Scholar] [CrossRef]
- Shao, B.; Long, R.; Ding, Y.; Wang, J.; Ding, L.; Wang, H. Morphological adaptations of yak (Bos grunniens) tongue to the foraging environment of the Qinghai-Tibetan Plateau. J. Anim. Sci. 2010, 88, 2594–2603. [Google Scholar] [CrossRef]
- Qiu, Q.; Zhang, G.; Ma, T.; Qian, W.; Wang, J.; Ye, Z.; Cao, C.; Hu, Q.; Kim, J.; Larkin, D.M.; et al. The yak genome and adaptation to life at high altitude. Nat. Genet. 2012, 44, 946–949. [Google Scholar] [CrossRef]
- Zhang, Z.; Xu, D.; Li, W.; Hao, J.; Wang, J.; Xin, Z.; Wang, W.; Qiu, Q.; Huang, X.; Zhou, J.; et al. Convergent evolution of rumen microbiomes in high-altitude mammals. Curr. Biol. 2016, 26, 1873–1879. [Google Scholar] [CrossRef]
- Feng, Y.L.; Wang, J.Q.; Yang, H.J.; Mo, F.; Wei, H.Y.; Huang, Y.X.; Feng, D.Y.; Wang, Z.H.; Gong, Y.S.; Li, S.C. Feed Standard of Beef Cattle; Ministry of Agrivulture of The People’s Republic of China: Beijing, China, 2004.
- Association of Official Analytical Chemists International. Official Methods of Analysis, 15th ed.; AOAC International: Arlington, TX, USA, 1990. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for Dietary Fiber, Neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Diary Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Ding, L.M.; Long, R.J.; Yang, Y.; Guo, X.S.; Wang, H.C. Behavioural responses by yaks in different physiological states (lactating, dry or replacement heifers), when grazing natural pasture in the spring (dry and germinating) season on the Qinghai-Tibetan plateau. Appl. Anim. Behav. Sci. 2007, 108, 239–250. [Google Scholar] [CrossRef]
- Schoonmaker, J.P.; Fluharty, F.L.; Loerch, S.C. Effect of source and amount of energy and rate of growth in the growing phase on adipocyte cellularity and lipogenic enzyme activity in the intramuscular and subcutaneous fat depots of Holstein steers. J. Anim. Sci. 2004, 82, 137–148. [Google Scholar] [CrossRef] [PubMed]
- Kertz, A.F.; Barton, B.A.; Reutzel, L.F. Relative Efficiencies of wither height and body weight increase from birth until first calving in Holstein cattle. J. Dairy Sci. 1998, 81, 1479–1482. [Google Scholar] [CrossRef]
- Jobgen, W.S.; Fried, S.K.; Fu, W.J.; Meininger, C.J.; Wu, G. Regulatory role for the arginine–nitric oxide pathway in metabolism of energy substrates. J. Nutr. Biochem. 2006, 17, 571–588. [Google Scholar] [CrossRef] [PubMed]
- Jaster, E.H.; Mccoy, G.C.; Spanski, N.; Tomkins, T. Effect of extra energy as fat or milk replacer solids in diets of young dairy calves on growth during cold weather. J. Dairy Sci. 1992, 75, 2524–2531. [Google Scholar] [CrossRef]
- Schwartzkopfgenswein, K.S.; Beauchemin, K.A.; Gibb, D.J.; Crews, D.H.; Hickman, D.D.; Streeter, M.; McAllister, T.A. Effect of bunk management on feeding behavior, ruminal acidosis and performance of feedlot cattle: A review. J. Anim. Sci. 2003, 25, 571–572. [Google Scholar]
- Curi, R.A.; de Oliveira, H.N.; Silveira, A.C.; Lopes, C.R. Association between IGF-I, IGF-IR and GHRH gene polymorphisms and growth and carcass traits in beef cattle. Livest. Prod. Sci. 2005, 94, 159–167. [Google Scholar] [CrossRef]
- Purchas, R.W.; Macmillan, K.L.; Hafs, H.D. Pituitary and plasma growth hormone levels in bulls from birth to one year of age. J. Anim. Sci. 1970, 31, 358–363. [Google Scholar] [CrossRef]
- Dong, L.F.; Zhang, W.B.; Zhang, N.F.; Tu, Y.; Diao, Q.Y. Feeding different dietary protein to energy ratios to Holstein heifers: Effects on growth performance, blood metabolites and rumen fermentation parameters. J. Anim. Physiol. Anim. Nutr. 2016, 101, 30–37. [Google Scholar] [CrossRef]
- Sejrsen, K.; Huber, J.T.; Tucker, H.A. Influence of Amount Fed on Hormone Concentrations and Their Relationship to Mammary Growth in Heifers. J. Dairy Sci. 1983, 66, 845–855. [Google Scholar] [CrossRef]
- Yambayamba, E.S.; Price, M.A.; Foxcroft, G.R. Hormonal status, metabolic changes, and resting metabolic rate in beef heifers undergoing compensatory growth. J. Anim. Sci. 1996, 74, 57–69. [Google Scholar] [CrossRef] [PubMed]
- Ford, J.A.; Park, C.S. Nutritionally directed compensatory growth enhances heifer development and lactation potential. J. Dairy Sci. 2001, 84, 1669–1678. [Google Scholar] [CrossRef]
- Houseknecht, K.L.; Boggs, D.L.; Campion, D.R.; Sartin, J.L.; Kiser, T.E.; Rampacek, G.B.; Amos, H.E. Effect of dietary energy source and level on serum growth hormone, insulin-like growth factor 1, growth and body composition in beef heifers. J. Anim. Sci. 1988, 66, 2916–2923. [Google Scholar] [CrossRef]
- Roberts, A.J.; Klindt, J.; Jenkins, T.G. Effects of varying energy intake and sire breed on duration of postpartum anestrus, insulin like growth factor-1, and growth hormone in mature crossbred cows. J. Anim. Sci. 2005, 83, 1705–1709. [Google Scholar] [CrossRef] [PubMed]
- Dauncey, M.J.; Burton, K.A.; White, P.; Harrison, A.P.; Gilmour, R.S.; Duchamp, C.; Cattaneo, D. Nutritional regulation of growth hormone receptor gene expression. FASEB J. 1994, 8, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Combes, S.; Louveau, I.; Bonneau, M. Moderate food restriction affects skeletal muscle and liver growth hormone receptors differently in pigs. J. Nutr. 1997, 127, 1944–1999. [Google Scholar] [CrossRef] [PubMed]
- Thissen, J.; Ketelslegers, J. Underwood LE. Nutritional regulation of the insulin-like growth factors. Endocr. Rev. 1994, 15, 80–101. [Google Scholar] [PubMed]
- Lucy, M.C.; Jiang, H.; Kobayashi, Y. Changes in the somatotrophic axis associated with the initiation of lactation. J. Dairy Sci. 2001, 84, E113–E119. [Google Scholar] [CrossRef]
- Groenewegen, P.P.; Mcbride, B.W.; Burton, J.H.; Elsasser, T.H. Effect of bovine somatotropin on the growth rate, hormone profiles and carcass composition of holstein bull calves. Domest. Anim. Endocrinol. 1990, 7, 43–54. [Google Scholar] [CrossRef]
- Moriel, P.; Johnson, S.E.; Vendramini, J.M.; Mercadante, V.R.; Hersom, M.J.; Arthington, J.D. Effects of calf weaning age and subsequent management system on growth and reproductive performance of beef heifers. J. Anim. Sci. 2014, 92, 3096–3107. [Google Scholar] [CrossRef]
- Lucy, M.C.; Beck, J.; Staples, C.R.; Head, H.H.; Rl, D.L.S.; Thatcher, W.W. Follicular dynamics, plasma metabolites, hormones and insulin-like growth factor I (IGF-I) in lactating cows with positive or negative energy balance during the preovulatory period. Reprod. Nutr. Dev. 1992, 32, 331–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersen, J.B.; Friggens, N.C.; Larsen, T.; Vestergaard, M.; Ingvartsen, K.L. Effect of energy density in the diet and milking frequency on plasma metabolites and hormones in early lactation dairy cows. J. Vet. Med. 2004, 51, 52–57. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, D.G.; Mcevoy, T.G.; Baxter, G.; Robinson, J.J.; Hogg, C.O.; Woad, K.J.; Webb, R.; Sinclair, K.D. Effect of dietary energy and protein on bovine follicular dynamics and embryo production in vitro: Associations with the ovarian insulin-like growth factor system. Biol. Reprod. 2001, 64, 1624–1632. [Google Scholar] [CrossRef] [PubMed]
- Bishop, M.D.; Simmen, R.C.; Simmen, F.A.; Davis, M.E. The relationship of insulin-like growth factor-I with postweaning performance in Angus beef cattle. J. Anim. Sci. 1989, 67, 2872–2880. [Google Scholar] [CrossRef] [PubMed]
- Dawson, J.M.; Craigon, J.; Buttery, P.J.; Beever, D.E. Influence of diet and P-agonist administration on plasma concentrations of growth hormone and insulin-like growth factor-1 in young steers. Br. J. Nutr. 1993, 70, 93–102. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Item | Treatment 1 | ||
|---|---|---|---|
| LE (%) | ME (%) | HE (%) | |
| Ingredient, % of DM | |||
| Corn | 32.00 | 44.00 | 56.00 |
| Corn germ | 28.00 | 20.00 | 12.00 |
| Wheat bran | 4.80 | 4.80 | 4.80 |
| DDGS 2 | 9.00 | 5.00 | 3.00 |
| Prickly ash seed | 4.00 | 4.00 | 4.00 |
| Cottonseed meal | 12.00 | 12.00 | 10.00 |
| Soybean meal | 5.30 | 5.30 | 5.30 |
| White stone powder | 2.00 | 2.00 | 2.00 |
| Dicalcium phosphate | 0.60 | 0.60 | 0.60 |
| Urea | 0.50 | 0.50 | 0.50 |
| Sodium bicarbonate | 1.00 | 1.00 | 1.00 |
| Premix 3 | 0.80 | 0.80 | 0.80 |
| Nutrient composition, % of DM | |||
| Crude protein | 16.53 | 16.74 | 17.21 |
| Crude fat | 3.73 | 4.18 | 5.57 |
| Acid detergent fiber | 4.54 | 4.14 | 3.72 |
| Neutral detergent fiber | 15.93 | 13.15 | 12.32 |
| Phosphorus | 0.31 | 0.34 | 0.36 |
| Calcium | 0.64 | 0.84 | 0.75 |
| NEg 4 (MJ/kg) | 5.5 | 6.2 | 6.9 |
| Gene Name 1 | Primer Sequence (5′→3′) | Accession | Product Size | |
|---|---|---|---|---|
| β-actin | F | ACCATCGGCAATGAGCG | DQ838049 | 150bp |
| R | CACCGTGTTGGCGTAGAG | |||
| GHR | F | AATGTGGTCCTTTCCC | NM176608 | 116bp |
| R | CAGAAGTAAGCGTTGTCC | |||
| IGF-1 | F | ATGCCCATCACATCCTCC | NM001077828 | 254bp |
| R | TACATCTCCAGCCTCCTCA | |||
| IGFBP-3 | F | ACAGACACCCAGAACTTCTCCTC | NM174556 | 236bp |
| R | GTTCAGGAACTTGAGGTGGTTC | |||
| Item 1 | Time 2 | Treatment 3 | SEM | p-Value | |||||
|---|---|---|---|---|---|---|---|---|---|
| LE | ME | HE | Treatment I 4 | Treatment II 5 | Time | Treatment × Time | |||
| BW, kg | 1~60 | 296.8 | 305.6 | 304.3 | 6.264 | 0.002 | <0.001 | 0.009 | |
| 1 | 276.0 | 277.0 | 275.2 | 2.555 | 0.785 | ||||
| 30 | 301.0 b | 310.2 a | 310.8 a | 3.427 | 0.037 | ||||
| 60 | 313.4 b | 330.0 a | 326.8 a | 4.357 | 0.012 | ||||
| ADG, kg/d | 1~60 | 0.624 b | 0.883 a | 0.860 a | 0.034 | <0.001 | <0.001 | 0.021 | |
| 1~30 | 0.833 b | 1.107 a | 1.186 a | 0.041 | <0.001 | ||||
| 30~60 | 0.413 b | 0.660 a | 0.533 ab | 0.062 | 0.013 | ||||
| ADFI, kg/d | 1~60 | 8.116 a | 7.958 | 7.780 b | 0.050 | 0.010 | |||
| Feed Conversion Ratio | 1~60 | 13.087 a | 9.028 b | 9.088 b | 0.537 | <0.001 | |||
| Item 1 | Time | BW | ADG | ||
|---|---|---|---|---|---|
| d 30 | d 60 | d 30 | d 60 | ||
| GH | d 30 | −0.175 | −0.493 | ||
| d 60 | −0.302 | −0.525 * | |||
| IGF-1 | d 30 | 0.200 | 0.606 * | ||
| d 60 | 0.338 | 0.674 ** | |||
| Item 1 | GHR | IGF-1 | IGFBP-3 |
|---|---|---|---|
| BW | −0.250 | 0.474 | 0.463 |
| ADG | −0.531 * | 0.431 | 0.715 ** |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, C.; Zhang, J.; Ahmad, A.A.; Bao, P.; Guo, X.; Long, R.; Ding, X.; Yan, P. Dietary Energy Levels Affect Growth Performance through Growth Hormone and Insulin-Like Growth Factor 1 in Yak (Bos grunniens). Animals 2019, 9, 39. https://doi.org/10.3390/ani9020039
Yang C, Zhang J, Ahmad AA, Bao P, Guo X, Long R, Ding X, Yan P. Dietary Energy Levels Affect Growth Performance through Growth Hormone and Insulin-Like Growth Factor 1 in Yak (Bos grunniens). Animals. 2019; 9(2):39. https://doi.org/10.3390/ani9020039
Chicago/Turabian StyleYang, Chao, Jianbo Zhang, Anum Ali Ahmad, Pengjia Bao, Xian Guo, Ruijun Long, Xuezhi Ding, and Ping Yan. 2019. "Dietary Energy Levels Affect Growth Performance through Growth Hormone and Insulin-Like Growth Factor 1 in Yak (Bos grunniens)" Animals 9, no. 2: 39. https://doi.org/10.3390/ani9020039