Indirect Influence of African Swine Fever Outbreak on the Raven (Corvus corax) Population


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
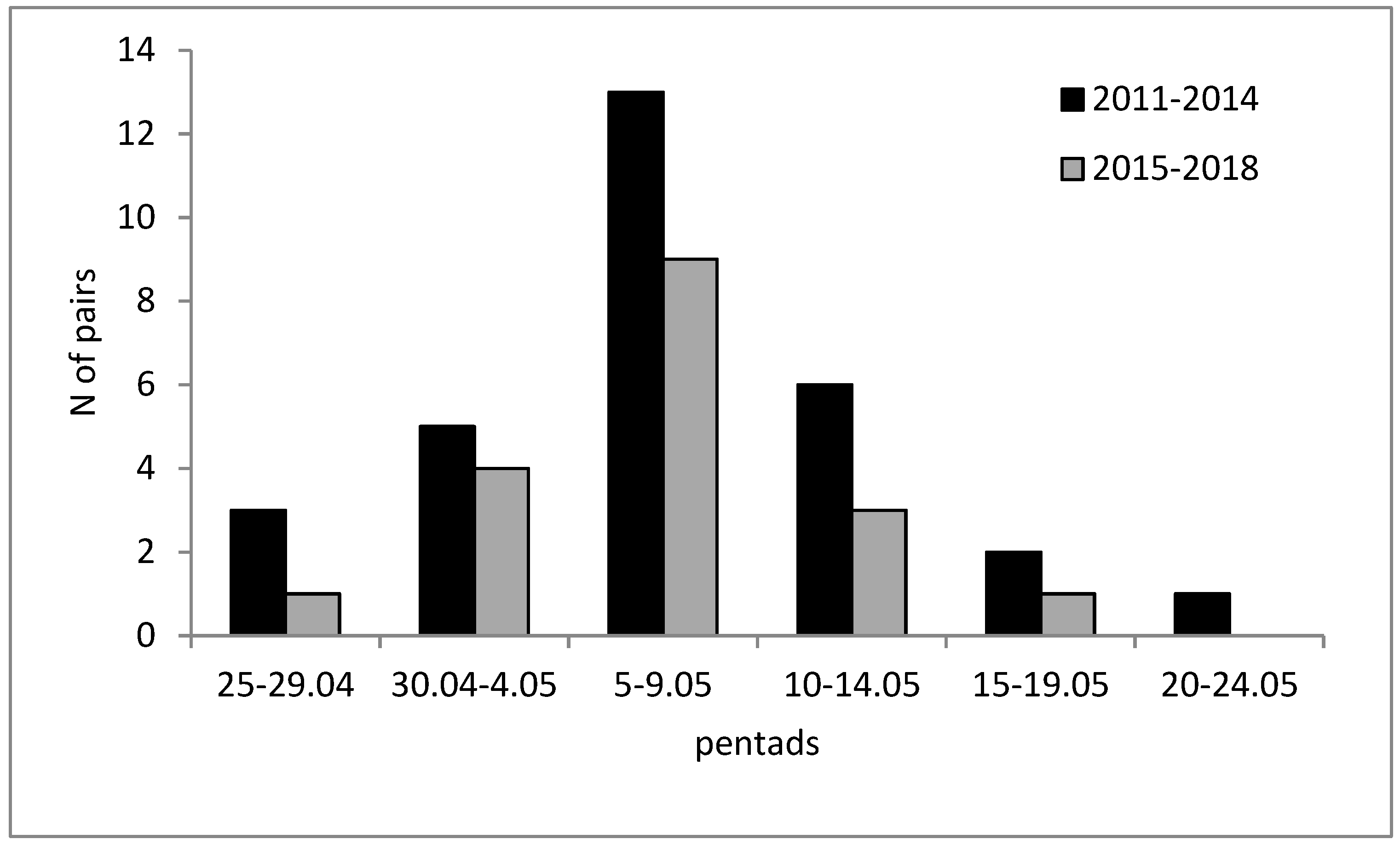
2.2. Raven Population in the Area
2.3. Methods
2.4. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Cramp, S.; Perrins, C.M. Handbook of the Birds of Europe, the Middle East. and North. Africa. The Birds of Western Palearctic. Vol. VIII Crows to Finches; Oxford University Press: Oxford, UK, 1994; ISBN 978-0198546795. [Google Scholar]
- Snow, D.W.; Perrins, C.M. The Birds of the Western Palearctic. Vol. 2; Oxford University Press: Oxford, UK, 2015; ISBN 978-3891047859. [Google Scholar]
- Glandt, D. Kolkrabe & Co.: Verhalten und Strategien Intelligenter Lebenskünstler; Aula-Verlag: Wiebelsheim, Germany, 2003; ISBN 978-3891047606. [Google Scholar]
- Zawadzka, D. Raven; Wydawnictwo Klubu Przyrodników: Świebodzin, Poland, 2006; ISBN 83-87846-88-0. [Google Scholar]
- Chylarecki, P.; Chodkiewicz, T.; Neubauer, G.; Sikora, A.; Meissner, W.; Woźniak, B.; Wylegała, P.; Ławicki, Ł.; Marchowski, D.; Betleja, J.; et al. Trendy Liczebności Ptaków w Polsce; Główny Inspektorat Ochrony Środowiska: Warszawa, Poland, 2018; ISBN 78-83-950881-0-0.
- Ratciffe, D. The Raven; Poyser: London, UK, 1997; ISBN 978-0856610905. [Google Scholar]
- Zawadzka, D.; Zawadzki, G. Synantropisation and synurbisation of raven Corvus corax in Poland: A review. Int. Stud. Sparrows 2014, 38, 11–16. [Google Scholar] [CrossRef]
- Engel, K.A.; Young, L.S. Spatial and temporal patterns in the diet of Common Ravens in southwestern Idaho. Condor 1989, 91, 372–378. [Google Scholar] [CrossRef]
- Chevallier, C.; Lai, S.; Berteaux, D. Predation of arctic fox Vulpes lagopus pups by common ravens Corvus corax. Pol. Biol. 2016, 39, 1335–1341. [Google Scholar] [CrossRef]
- Nogales, M.; Hernandez, E.C. Interinsular variations in the spring and summer diet of the Raven Corvus corax in the Canary Island. Ibis 1994, 136, 441–447. [Google Scholar] [CrossRef]
- Nogales, M.; Hernández, E.C.; Valdés, F. Seed dispersal by common ravens Corvus corax among island habitats (Canarian Archipelago). Ecoscience 1999, 6, 56–61. [Google Scholar] [CrossRef]
- Kristan, W.B.; Boarman, W.I. Spatial pattern of risk of Common Raven predation on desert tortoises. Ecology 2003, 84, 2432–2443. [Google Scholar] [CrossRef]
- Marquiss, M.; Booth, C.J. The diet of Ravens Corvus corax in Orkney. Bird Study 1986, 33, 190–195. [Google Scholar] [CrossRef]
- Tome, D.; Krofel, M.; Mihelič, T. The diet of the raven Corvus corax in south-west Slovenia. Annal. Ser. Hist. Nat. 2009, 19, 161–166. [Google Scholar]
- Ewins, P.J.; Dymond, J.N.; Marquiss, M. The distribution, breeding and diet of Ravens Corvus corax in Shetland. Bird Study 1986, 33, 110–116. [Google Scholar] [CrossRef]
- Dare, P.J. Raven Corvus corax population in two upland regions of north Wales. Bird Study 1986, 33, 179–189. [Google Scholar] [CrossRef]
- Zając, K. The diet of Raven Corvus corax in the fish pond area near Jelenia Góra (SW Poland). In Corvids of Poland; Jerzak, L., Kavanagh, B.P., Tryjanowski, P., Eds.; Bogucki Wydawnictwo Naukowe: Poznań, Poland, 2005; pp. 367–372. ISBN 83-89290-92-82005. [Google Scholar]
- Rösner, S.; Selva, N.; Muller, T.; Pugacewicz, E.; Laudet, F. Raven Corvus corax ecology in a primeval temperate forest. In Corvids of Poland; Jerzak, L., Kavanagh, B.P., Tryjanowski, P., Eds.; Bogucki Wydawnictwo Naukowe: Poznań, Poland, 2005; pp. 385–405. ISBN 83-89290-92-82005. [Google Scholar]
- Selva, N.; Jędrzejewska, B.; Jędrzejewski, W.; Wajrak, A. Factors affecting carcass use by a guild of scavengers on European temperate woodland. Can. J. Zool. 2005, 83, 1590–1601. [Google Scholar] [CrossRef]
- General Veterinary Inspectorate. Available online: https://bip.wetgiw.gov.pl/asf/mapa (accessed on 9 September 2018).
- Pejsak, Z.; Truszczyński, M.; Niemczuk, K.; Kozak, E.; Markowska-Daniel, I. Epidemiology of African Swine Fever in Poland since the detection of first case. Pol. J. Vet. Sci. 2014, 17, 665–672. [Google Scholar] [CrossRef] [PubMed]
- Śmietanka, K.; Woźniakowski, G.; Kozak, E.; Niemczuk, K.; Frączyk, M.; Bocian, Ł.; Kowalczyk, A.; Pejsak, Z. African Swine Fever epidemic, Poland, 2014–2015. Emerg. Infect. Dis. 2016, 22, 1201–1207. [Google Scholar] [CrossRef] [PubMed]
- Abrahantes, J.C.; Gogin, A.; Richardson, J.; Gervelmeyer, A. Epidemiological analyses on African swine fever in the Baltic countries and Poland. EFSA J. 2017, 15, 4732. [Google Scholar] [CrossRef]
- Galindo, I.; Alonso, C. African swine fever virus: A review. Viruses 2017, 9, 103. [Google Scholar] [CrossRef] [PubMed]
- Ministry of the Environment. Available online: www.mos.gov.pl access (accessed on 10 November 2018).
- Goszczyński, J.; Gryz, J.; Krauze, D. Fluctuations of a Common Buzzard Buteo buteo Population in Central Poland. Acta Ornithol. 2005, 40, 75–78. [Google Scholar] [CrossRef]
- Krauze-Gryz, D.; Gryz, J. Free-ranging domestic dogs (Canis familiaris) in central Poland: Density, penetration range and diet composition. Pol. J. Ecol. 2014, 62, 183–193. [Google Scholar] [CrossRef]
- Krauze-Gryz, D.; Żmihorski, M.; Jasińska, K.; Kwaśny, Ł.; Werka, J. Temporal pattern of wildlife-train collisions in Poland. J. Wildl. Manag. 2017, 82, 1513–1519. [Google Scholar] [CrossRef]
- Lagos, L.; Picos, J.; Valero, E. Temporal pattern of wild ungulate-related traffic accidents in northwest Spain. Eur. J. Wildl. Res. 2012, 58, 661–668. [Google Scholar] [CrossRef]
- Zaborowski, S.; Warsaw University of Life Sciences, Warsaw, Poland. Personal communication, 2001.
- Gryz, J.; Krauze-Gryz, D. The influence of raptors (Falconiformes) and raven (Corvus corax) on populations of game animals. Ann. Warsaw Univ. Life Sci. SGGW For. Wood Technol. 2014, 86, 114–124. [Google Scholar]
- Goszczyński, J.; Warsaw University of Life Sciences, Warsaw, Poland. Personal communication, 2003.
- Pucek, Z. (Ed.) Keys to Vertebrates of Poland-Mammals; Polish Scientific Publishers: Warsaw, Poland, 1981; ISBN 978-8301025533. [Google Scholar]
- Engelmann, W.E.; Fritzsche, J.; Gunther, R.; Obst, F.J. Lurche und Kriechtiere Europas; Neuman Verlag: Leipzig, Germany, 1985; ISBN 3-423-03263-4. [Google Scholar]
- Moreno, E. Clave osteologica para la identification de los Passeriformes Ibericos. 1. Aegithalidae, Remizidae, Fringiliade, Alaudidae. Ardeola 1985, 32, 295–377. [Google Scholar]
- Moreno, E. Clave osteologica para la identification de los Passeriformes Ibericos. 2. Hirundinidae, Prunellidae, Sittidae, Cerothidae, Troglotytidae, Cinclidae, Lanidae, Oriolodae, Corvidae, Sturnidae, Motacillidae. Ardeola 1986, 33, 69–129. [Google Scholar]
- März, R. Gewöll—Und Rupfungskunde; Akademie-Verlang: Berlin, Germany, 1987; ISBN 978-3891047620. [Google Scholar]
- Ujhelyi, P. Identification of the Fringillidae of Europe on the basis of craniometric characteristics. Aquila 1992, 99, 99–110. [Google Scholar]
- Brown, R.; Ferguson, J.; Lawrence, M.; Lees, D. Tracks & Signs of the Birds of Britain and Europe an Identification Guide; A & C Black: London, UK, 1999; ISBN 978-0713653823. [Google Scholar]
- Dziurdzik, B. Key to the identification of hairs of mammals from Poland. Acta Zool. Crac. 1973, 18, 73–91. [Google Scholar]
- Brunner, H.; Coman, B. The Identification of Mammalian Hair; Intaka Press: Melbourne, Australia, 1974; ISBN 9780909605018. [Google Scholar]
- Dziurdzik, B. Badania nad identyfikacją gatunków ssaków na podstawie budowy histologicznej włosów. Prz. Zool. 1978, 22, 73–91. [Google Scholar]
- Teerink, B.J. Hair of West-European Mammals; Cambridge University Press: Cambridge, UK, 1991; ISBN 978-0521545778. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Dałkowski, D.; Rogów Experimental Forest Station, Rogów, Poland. Personal communication, 2017.
- Newton, I.; Davis, P.E.; Davis, J.E. Ravens and buzzards in relation to sheep farming and forestry in Wales. J. Appl. Ecol. 1982, 19, 681–706. [Google Scholar] [CrossRef]
- Marquiss, M.; Newton, I.; Ratcliffe, D.A. The decline of raven, Corvus corax, in relation to afforestation in southern Scotland and northern England. J. Appl. Ecol. 1978, 15, 129–144. [Google Scholar] [CrossRef]
- Pugacewicz, E. Breeding Birds of the Białoweża Forest; Północnopodlaskie Towarzystwo Ochrony Ptaków: Białystok, Poland, 1997; ISBN 83-903553-5-3. [Google Scholar]
- Chojnacka-Ożga, L.; Ożga, W. Air temperature anomalies in experimental forests in Rogów in 1924–2015. For. Res. Pap. 2018, 79, 37–44. [Google Scholar] [CrossRef]
- Gryz, J.; Chojnacka-Ożga, L.; Krauze-Gryz, D. A tawny owl (Strix aluco) population remains stable despite changing environmental conditions—A case study from central Poland. Pol. J. Ecol. 2019, in press. [Google Scholar]
- Woźniak, M.; Gawrysiak, K.; Woźniak, B. Numbers and breeding place characteristics of common raven (Corvus corax) in Sobibór Forest. Studia i Materiały CEPL w Rogowie 2018, 56, 74–84. [Google Scholar]
- Bednorz, J. Ravens Corvus corax Linnaeus 1758, nesting on electricity pylons in the Wielkopolska region. Acta Zool. Crac. 2000, 43, 177–184. [Google Scholar]
- Quelennec, T. Le grand corbeau Corvus corax en Bretange. Alauda 2001, 69, 19–24. [Google Scholar]
- Ruffino, L.; Salo, P.; Koivisto, E.; Banks, P.; Korpimäki, E. Reproductive responses of birds to experimental food supplementation: A meta-analysis. Front. Zool. 2014, 11, 80. [Google Scholar] [CrossRef] [PubMed]
- Kekkonen, J.; Kolunen, H.; Pietiäinen, H.; Karell, P.; Brommer, J.E. Tawny owl reproduction and offspring sex ratios under variable food conditions. J. Ornithol. 2008, 149, 59–66. [Google Scholar] [CrossRef]
- Roulin, A.; Ducret, B.; Ravussin, P.A.; Bize, P. Importance des mulots Apodemus sp. dans le succès de reproduction de la Choulette hulotte Strix aluco en Suisse romande. Nos Oiseaux 2009, 56, 19–25. [Google Scholar]
- Donázar, J.A.; Cortés-Avizanda, A.; Carrete, M. Dietary shifts in two vultures after the demise of supplementary feeding stations: Consequences of the EU sanitary legislation. Eur. J. Wildl. Res. 2010, 56, 613–621. [Google Scholar] [CrossRef]
- Margalida, A.; Donázar, J.A.; Carrete, M.; Sánchez-Zapata, J.A. Sanitary versus environmental policies: Fitting together two pieces of the puzzle of European vulture conservation. J. Appl. Ecol. 2010, 47, 931–935. [Google Scholar] [CrossRef]
- Margalida, A.; Colomer, M.À.; Oro, D. Man-induced activities modify demographic parameters in a long-lived species: Effects of poisoning and health policies. Ecol. Appl. 2014, 24, 436–444. [Google Scholar] [CrossRef]
- Gangoso, L.; Agudo, R.; Anadón, J.D.; de la Riva, M.; Suleyman, A.S.; Porter, R.; Donázar, J.A. Reinventing mutualism between humans and wild fauna: Insights from vultures as ecosystem services providers. Conserv. Lett. 2013, 6, 172–179. [Google Scholar] [CrossRef]
- Markandya, A.; Taylor, T.; Longo, A.; Murty, M.N.; Murty, S.; Dhavala, K. Counting the cost of vulture decline—An appraisal of the human health and other benefits of vultures in India. Ecol. Econ. 2008, 67, 194–204. [Google Scholar] [CrossRef] [Green Version]
- Whelan, C.J.; Wenny, D.G.; Marquis, R.J. Ecosystem services provided by birds. Ann. N. Y. Acad. Sci. 2008, 1134, 25–60. [Google Scholar] [CrossRef] [PubMed]
- Devault, T.L.; Beasley, J.C.; Olson, Z.H.; Moleón, M.; Carrete, M.; Margalida, A.; Sánchez-Zapata, J.A. Ecosystem Services Provided by Avian Scavengers. In Why Birds Matter: Avian Ecological Function and Ecosystem Services; Sekercioglu, Ç.H., Wenny, D.G., Whelan, C.J., Eds.; University of Chicago Press: Chicago, IL, USA, 2016; pp. 235–270. [Google Scholar]
- Krauze-Gryz, D.; Gryz, J.; Goszczyński, J.; Chylarecki, P.; Żmihorski, M. The good, the bad, and the ugly: Space use and intraguild interactions among three opportunistic predators—Cat (Felis catus), dog (Canis lupus familiaris), and red fox (Vulpes vulpes)—Under human pressure. Can. J. Zool. 2012, 90, 1402–1413. [Google Scholar] [CrossRef]
- Zawadzka, D. Distribution, habitat selection, food and reproduction of the raven (Corvus corax) in the National Park of Lake Wigry. Notatki Ornitol. 1996, 37, 225–245. [Google Scholar]
- Zawadzka, D.; Drozdowski, S.; Zawadzki, G.; Zawadzki, J. Predator or opportunist? Feeding ecology of raven Corvus corax in Augustów Forest. Studia Materiały CEPL Rogowie 2013, 36, 210–220. [Google Scholar]
- Heinrih, B. Ravens in Winter; Vintage Books: New York, NY, USA, 1989; ISBN 978-0679732365. [Google Scholar]


{kind=link}
| Year/Period | N of Breeding Pairs | Breeding Success (%) | N juv. per | |
|---|---|---|---|---|
| Breeding Pair | Successful Pair | |||
| 2011 | 13 | 61 | 1.5 | 2.5 |
| 2012 | 13 | 61 | 1.5 | 2.5 |
| 2013 | 11 | 63 | 1.6 | 2.6 |
| 2014 | 12 | 75 | 1.7 | 1.7 |
| (SD) | 12.3 (1.0) | 65.0 | 1.6 (1.3) | 2.3 (0.7) |
| 2015 | 8 | 87 | 1.8 | 2.0 |
| 2016 | 6 | 33 | 0.8 | 2.5 |
| 2017 | 7 | 71 | 1.1 | 1.6 |
| 2018 | 9 | 55 | 0.9 | 1.6 |
| (SD) | 7.5 (1.3) | 61.5 | 1.2 (1.7) | 1.9 (0.7) |
| Food Category | 2011–2014 | 2015–2018 | ||
|---|---|---|---|---|
| Pellets | Pellets | |||
| N | % | N | % | |
| Domestic pig | 156 | 32.9 | 56 | 11.6 |
| Wild boar | 19 | 4.0 | 10 | 2.1 |
| Domestic cow | 5 | 1.1 | 0.0 | |
| Domestic dog | 14 | 3.0 | 39 | 8.1 |
| Domestic cat | 4 | 0.8 | 15 | 3.1 |
| Domestic rabbit | 2 | 0.4 | 0.0 | |
| Moose | 3 | 0.6 | 0.0 | |
| Roe deer | 27 | 5.7 | 49 | 10.1 |
| Racoon dog | 3 | 0.6 | 0.0 | |
| Red fox | 1 | 0.2 | 4 | 0.8 |
| Mustela sp. | 1 | 0.2 | 0.0 | |
| Brown hare | 1 | 0.2 | 0.0 | |
| Soricomorphs | 58 | 12.2 | 90 | 18.6 |
| Small rodent | 78 | 16.5 | 140 | 28.9 |
| Chicken | 95 | 20.0 | 116 | 24.0 |
| Domestic pigeon | 2 | 0.4 | 5 | 1.0 |
| Small bird | 43 | 9.1 | 82 | 16.9 |
| Eggs | 34 | 7.2 | 62 | 12.8 |
| Fish | 54 | 11.4 | 63 | 13.0 |
| Amphibians | 15 | 3.2 | 29 | 6.0 |
| Reptiles | 18 | 3.8 | 24 | 5.0 |
| Invertebrates | 258 | 54.4 | 329 | 68.0 |
| Grains | 268 | 56.5 | 300 | 62.0 |
| Fruits | 44 | 9.3 | 58 | 12.0 |
| Unrecognized plants | 135 | 28.5 | 169 | 34.9 |
| Inorganic elements | 139 | 29.3 | 183 | 37.8 |
| Total N of pellets | 474 | 484 | ||
| Food Category | 2011–2014 | 2015–2018 | ||
|---|---|---|---|---|
| Observations | Observations | |||
| N | % | N | % | |
| Domestic pig | 66 | 27.2 | 14 | 4.9 |
| Wild boar | 10 | 4.1 | 2 | 0.7 |
| Domestic dog | 2 | 0.8 | 6 | 2.1 |
| Domestic cat | 2 | 0.8 | 3 | 1.0 |
| Roe deer | 9 | 3.7 | 21 | 7.3 |
| Small rodent | 4 | 1.6 | 9 | 3.1 |
| Mole | 2 | 0.8 | 6 | 2.1 |
| Chicken | 54 | 22.2 | 85 | 29.5 |
| Small birds | 1 | 0.4 | 2 | 0.7 |
| Eggs | 2 | 0.8 | 1 | 0.3 |
| Fish | 3 | 1.2 | 6 | 2.1 |
| Insects | 6 | 2.5 | 10 | 3.5 |
| Earthworm | 4 | 1.6 | 8 | 2.8 |
| Plant food | 41 | 16.9 | 60 | 20.8 |
| Garbage | 20 | 8.2 | 34 | 11.8 |
| Industrial waste | 12 | 4.9 | 15 | 5.2 |
| Excrements | 5 | 2.1 | 6 | 2.1 |
| Total N of observations | 243 | 100 | 288 | 100 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gryz, J.; Krauze-Gryz, D. Indirect Influence of African Swine Fever Outbreak on the Raven (Corvus corax) Population. Animals 2019, 9, 41. https://doi.org/10.3390/ani9020041
Gryz J, Krauze-Gryz D. Indirect Influence of African Swine Fever Outbreak on the Raven (Corvus corax) Population. Animals. 2019; 9(2):41. https://doi.org/10.3390/ani9020041
Chicago/Turabian StyleGryz, Jakub, and Dagny Krauze-Gryz. 2019. "Indirect Influence of African Swine Fever Outbreak on the Raven (Corvus corax) Population" Animals 9, no. 2: 41. https://doi.org/10.3390/ani9020041