Sperm Morphology in Neotropical Primates

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Analyzed Specimens
2.2. Ethical Note
2.3. Semen Collection
2.4. Sperm Morphometric Analysis
2.5. Statistical Analysis
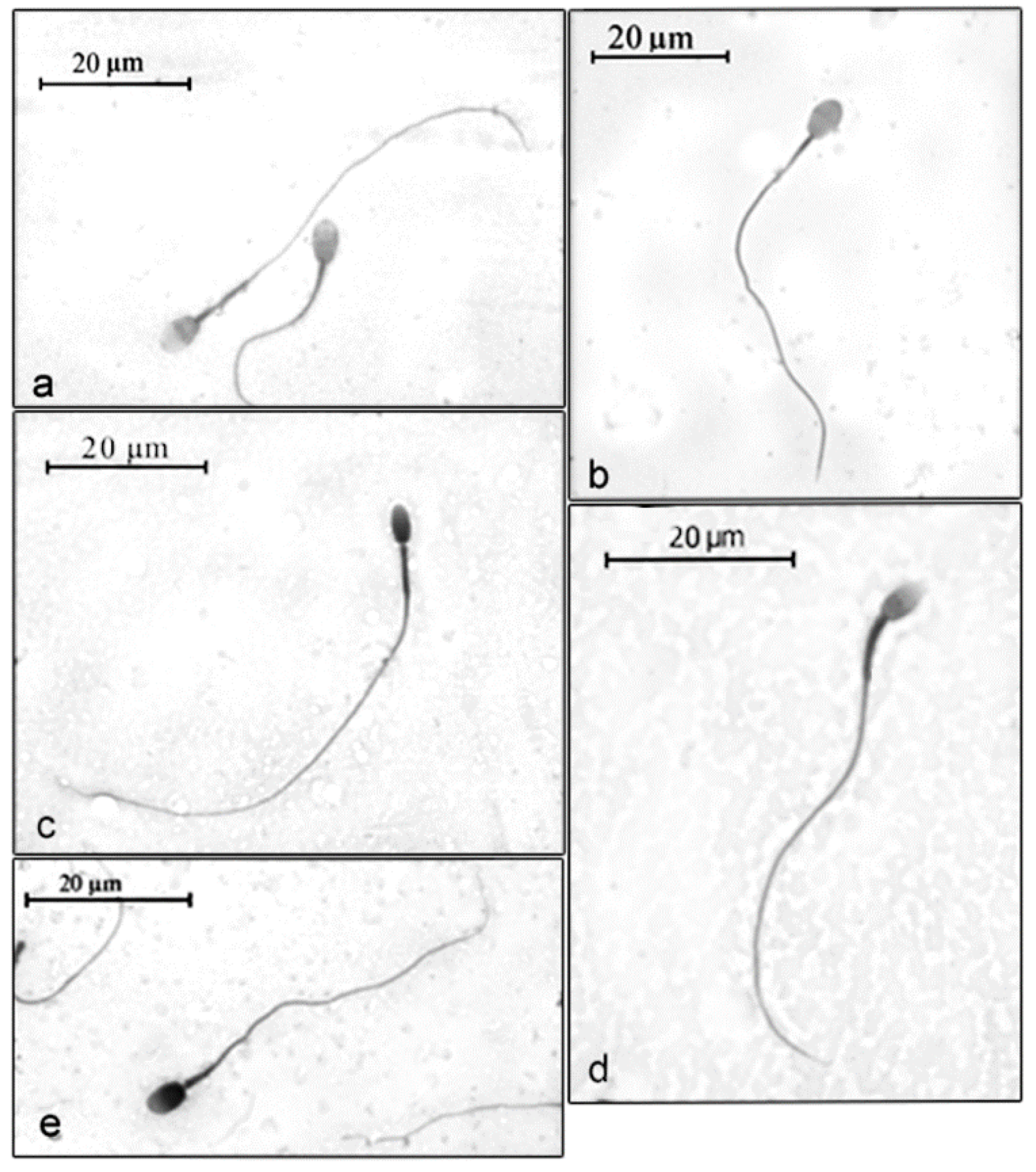
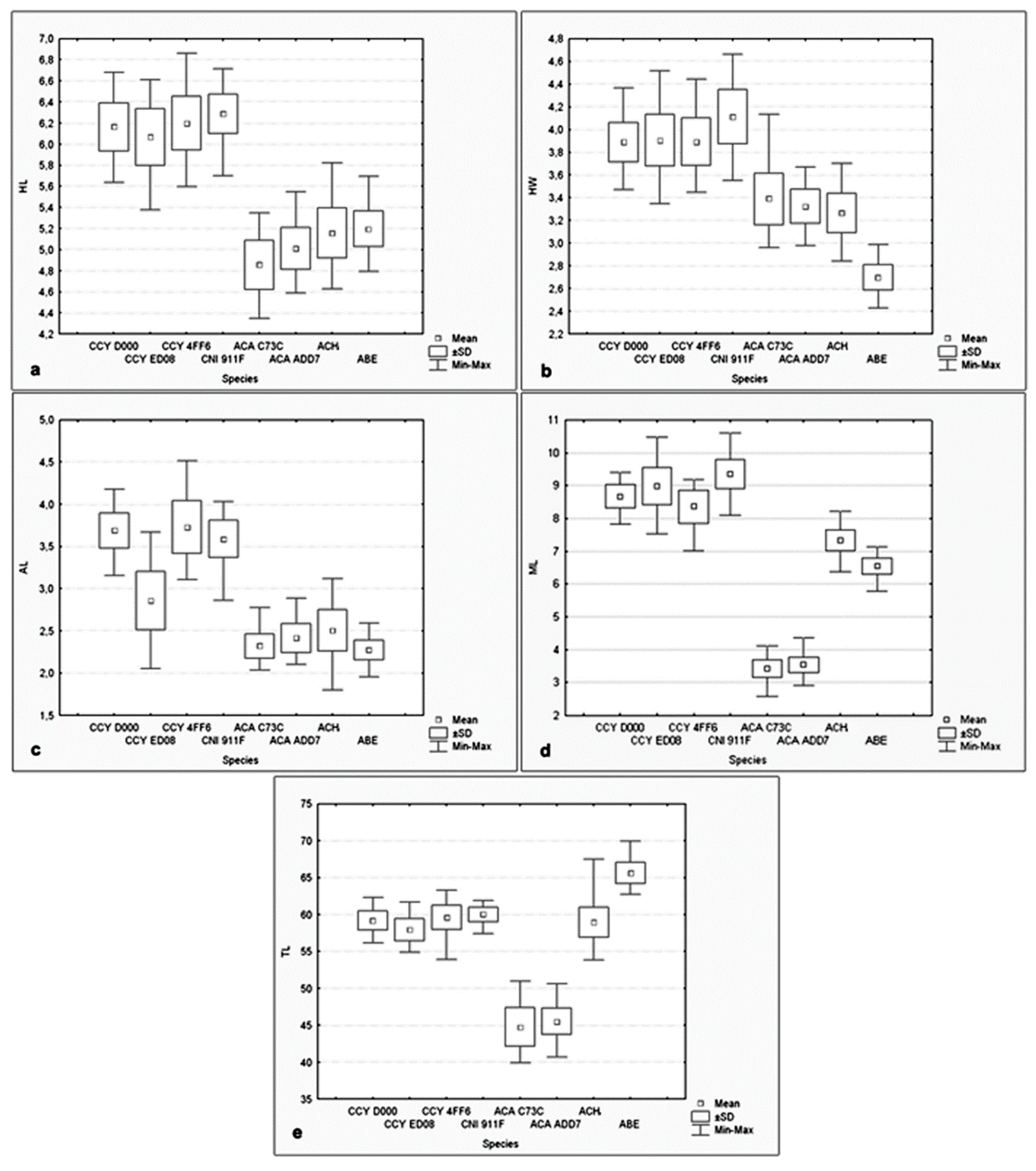
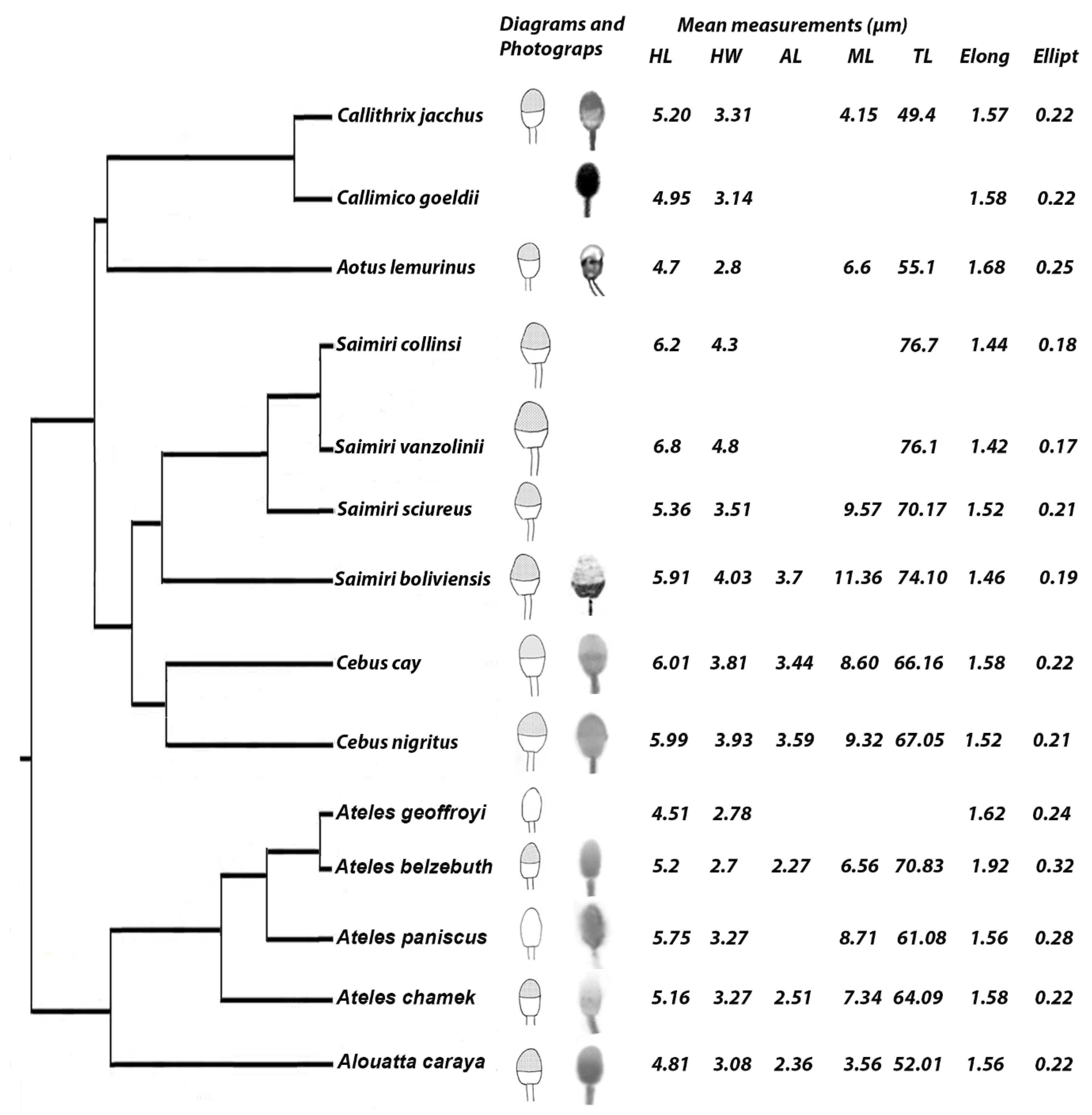
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fawcett, D.W. The mammalian spermatozoon. Dev. Biol. 1975, 44, 394–436. [Google Scholar] [CrossRef]
- Cummins, J.M.; Woodall, P.F. On mammalian sperm dimensions. J. Reprod. Fertil. 1985, 75, 153–175. [Google Scholar] [CrossRef] [PubMed]
- Roldan, E.R.S.; Gomendio, M.; Vitullo, A.D. The Evolution of Eutherian Spermatozoa and Underlying Selective Forces: Female Selection and Sperm Competition. Biol. Rev. 1992, 67, 551–593. [Google Scholar] [CrossRef] [PubMed]
- Downing Meisner, A.; Klaus, A.V.; O’Leary, M.A. Sperm head morphology in 36 species of artiodactylans, perissodactylans, and cetaceans (Mammalia). J. Morphol. 2005, 263, 179–202. [Google Scholar] [CrossRef] [PubMed]
- Cetica, P.; Solari, A.; Merani, M.; De Rosas, J.; Burgos, M. Evolutionary sperm morphology and morphometry in armadillos. J. Submicrosc. Cytol. Pathol. 1998, 30, 309–314. [Google Scholar]
- Birkhead, T.R.; Hosken, D.J.; Pitnick, S.S. Sperm Biology: An Evolutionary Perspective; Academic Press: Burlington, MA, USA, 2008; ISBN 978-0-08-091987-4. [Google Scholar]
- Breed, W.; Aplin, K. Sperm Morphology of Murid Rodents from New-Guinea and the Solomon-Islands—Phylogenetic Implications. Aust. J. Zool. 1995, 43, 17–30. [Google Scholar] [CrossRef]
- Gallardo, M.; Mondaca, F.; Ojeda, R.; Kohler, N.; Garrido, O. Morphological Diversity in the Sperms of Caviomorph Rodents. Mastozool. Neotrop. 2002, 9, 159–170. [Google Scholar]
- Gage, M.J.G. Mammalian Sperm Morphometry. Proc. Biol. Sci. 1998, 265, 97–103. [Google Scholar] [CrossRef]
- Anderson, M.J.; Dixson, A.F. Sperm competition: Motility and the midpiece in primates. Nature 2002, 416, 496. [Google Scholar] [CrossRef]
- Anderson, M.J.; Nyholt, J.; Dixson, A.F. Sperm competition and the evolution of sperm midpiece volume in mammals. J. Zool. 2005, 267, 135–142. [Google Scholar] [CrossRef]
- Gomendio, M.; Roldan, E.R.S. Implications of diversity in sperm size and function for sperm competition and fertility. Int. J. Dev. Biol. 2008, 52, 439–447. [Google Scholar] [CrossRef] [PubMed]
- Tourmente, M.; Gomendio, M.; Roldan, E.R. Sperm competition and the evolution of sperm design in mammals. BMC Evol. Biol. 2011, 11, 12. [Google Scholar] [CrossRef] [PubMed]
- Malo, A.F.; Gomendio, M.; Garde, J.; Lang-Lenton, B.; Soler, A.J.; Roldan, E.R.S. Sperm design and sperm function. Biol. Lett. 2006, 2, 246–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomendio, M.; Roldan, E.R.S. Sperm competition influences sperm size in mammals. Proc. R Soc. Lond. B 1991, 243, 181–185. [Google Scholar]
- Smith, R.L. Sperm Competition and the Evolution of Animal Mating Systems; Elsevier: Orlando, Florida, USA, 1984; ISBN 978-0-323-14313-4. [Google Scholar]
- Martin, D.E.; Gould, K.G.; Warner, H. Comparative morphology of primate spermatozoa using scanning electron microscopy. I. Families Hominidae, Pongidae, Cercopithecidae and Cebidae. J. Hum. Evol. 1975, 4, 287–292. [Google Scholar] [CrossRef]
- Nakazato, C.; Yoshizawa, M.; Isobe, K.; Kusakabe, K.T.; Kuraishi, T.; Hattori, S.; Matsumoto, H.; Fukui, E.; Kuwahata, A.; Ochi, M.; et al. Morphological Characterization of Spermatozoa of the Night Monkey. J. Mamm. Ova Res. 2015, 32, 37–40. [Google Scholar] [CrossRef]
- Sampaio, W.V.; Oliveira, K.G.; Leão, D.L.; Caldas-Bussiere, M.C.; Queiroz, H.L.; Paim, F.P.; Santos, R.R.; Domingues, S.F.S. Morphologic analysis of sperm from two neotropical primate species: Comparisons between the squirrel monkeys Saimiri collinsi and Saimiri vanzolinii. Zygote 2017, 25, 141–148. [Google Scholar] [CrossRef]
- Steinberg, E.R.; Nieves, M.; Ascunce, M.S.; Palermo, A.M.; Mudry, M.D. Morphological and Genetic Characterization of Saimiri boliviensis. Int. J. Primatol. 2009, 30, 29–31. [Google Scholar] [CrossRef]
- Swanson, W.F.; Valle, R.R.; Carvalho, F.M.; Arakaki, P.R.; Rodas-Martínez, A.Z.; Muniz, J.; García-Herreros, M. Sperm Morphology Assessment in Captive Neotropical Primates. Reprod. Domest. Anim. 2016, 51, 623–627. [Google Scholar] [CrossRef]
- Valle, R.R.; Arakaki, P.R.; Carvalho, F.M.; Muniz, J.A.P.C.; Leal, C.L.V.; García-Herreros, M. Identification of sperm head subpopulations with defined pleiomorphic characteristics in ejaculates of captive Goeldi’s monkeys (Callimico goeldii). Anim. Reprod. Sci. 2013, 137, 93–102. [Google Scholar] [CrossRef]
- Valle, R.R.; Carvalho, F.M.; Muniz, J.A.P.C.; Leal, C.L.V.; García-Herreros, M. Differential distribution of sperm subpopulations and incidence of pleiomorphisms in ejaculates of captive howling monkeys (Alouatta caraya). Naturwissenschaften 2013, 100, 923–933. [Google Scholar] [CrossRef] [PubMed]
- Valle, R.R.; Nayudu, P.L.; Leal, C.L.V.; García-Herreros, M. Sperm head morphometry in ejaculates of adult marmosets (Callithrix jacchus): A model for studying sperm subpopulations and among-donor variations. Theriogenology 2012, 78, 1152–1165. [Google Scholar] [CrossRef] [PubMed]
- Kluge, A.G. A Concern for Evidence and a Phylogenetic Hypothesis of Relationships Among Epicrates (Boidae, Serpentes). Syst. Zool. 1989, 38, 7–25. [Google Scholar] [CrossRef]
- Hernández-López, L.; Parra, G.C.; Cerda-Molina, A.L.; Pérez-Bolaños, S.C.; Sánchez, V.D.; Mondragón-Ceballos, R. Sperm quality differences between the rainy and dry seasons in captive black-handed spider monkeys (Ateles geoffroyi). Am. J. Primatol. 2002, 57, 35–41. [Google Scholar] [CrossRef]
- Moreland, R.B.; Richardson, M.E.; Lamberski, N.; Long, J.A. Characterizing the Reproductive Physiology of the Male Southern Black Howler Monkey, Alouatta caraya. J. Androl. 2001, 22, 395–403. [Google Scholar]
- Flores-Herrera, H.; Acuña-Hernández, D.G.; Rivera-Rebolledo, J.A.; González-Jiménez, M.A.; Rodas-Martínez, A.Z.; Swanson, W.F. Effect of increasing trypsin concentrations on seminal coagulum dissolution and sperm parameters in spider monkeys (Ateles geoffroyi). Theriogenology 2012, 78, 612–619. [Google Scholar] [CrossRef]
- Yu, J.-F.; Lai, Y.-H.; Wang, T.-E.; Wei, Y.-S.; Chang, Y.-J.; Li, S.-H.; Chin, S.-C.; Joshi, R.; Chang, H.-W.; Tsai, P.-S. The effects of type I collagenase on the degelification of chimpanzee (Pan troglodytes) semen plug and sperm quality. BMC Vet. Res. 2018, 14, 58. [Google Scholar] [CrossRef]
- Björndahl, L.; Mortimer, D.; Barratt, C.L.R.; Castilla, J.A.; Menkveld, R.; Kvist, U.; Alvarez, J.G.; Haugen, T.B. A Practical Guide to Basic Laboratory Andrology; Cambridge University Press: Cambridge, UK, 2010; ISBN 978-1-139-48249-3. [Google Scholar]
- Watson, P.F. Use of a Giemsa stain to detect changes in acrosomes of frozen ram spermatozoa. Vet. Rec. 1975, 97, 12–15. [Google Scholar] [CrossRef]
- Tamuli, M.K.; Watson, P.F. Use of a simple staining technique to distinguish acrosomal changes in the live sperm sub-population. Anim. Reprod. Sci. 1994, 35, 247–254. [Google Scholar] [CrossRef]
- Dumas, F.; Mazzoleni, S. Neotropical primate evolution and phylogenetic reconstruction using chromosomal data. Eur. Zool. J. 2017, 84, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Lima, M.G.M.; Silva-Júnior, J.; Černý, D.; Buckner, J.C.; Aleixo, A.; Chang, J.; Zheng, J.; Alfaro, M.E.; Martins, A.; Di Fiore, A.; et al. A phylogenomic perspective on the robust capuchin monkey (Sapajus) radiation: First evidence for extensive population admixture across South America. Mol. Phylogenet. Evol. 2018, 124, 137–150. [Google Scholar] [CrossRef] [PubMed]
- Lynch Alfaro, J.W.; Boubli, J.P.; Paim, F.P.; Ribas, C.C.; da Silva, M.N.F.; Messias, M.R.; Röhe, F.; Mercês, M.P.; Silva Júnior, J.S.; Silva, C.R.; et al. Biogeography of squirrel monkeys (genus Saimiri): South-central Amazon origin and rapid pan-Amazonian diversification of a lowland primate. Mol. Phylogenet. Evol. 2015, 82, 436–454. [Google Scholar] [CrossRef] [PubMed]
- Nieves, M.; Ascunce, M.S.; Rahn, M.I.; Mudry, M.D. Phylogenetic relationships among some Ateles species: The use of chromosomic and molecular characters. Primates 2005, 46, 155–164. [Google Scholar] [CrossRef]
- Oliveira, E.H.C.; Neusser, M.; Müller, S. Chromosome Evolution in New World Monkeys (Platyrrhini). Cytogenet. Genome Res. 2012, 137, 259–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz-García, M.; Escobar-Armel, P.; Leguizamon, N.; Manzur, P.; Pinedo-Castro, M.; Shostell, J.M. Genetic characterization and structure of the endemic Colombian silvery brown bare-face tamarin, Saguinus leucopus (Callitrichinae, Cebidae, Primates). Primates 2014, 55, 415–435. [Google Scholar] [CrossRef]
- Steinberg, E.; Nieves, M.; Mudry, M.D. Multiple sex chromosome systems in howler monkeys (Platyrrhini, Alouatta). Comp. Cytogenet. 2014, 8, 43–69. [Google Scholar] [CrossRef]
- Steinberg, E.R.; Mudry, M.D. Células germinales en primates: Una mirada evolutiva. In Proceedings of the Actas de las XII Jornadas Nacionales de Antropología Biológica, Corrientes, Provincia de Corrientes, Argentina, 22–25 September 2015. [Google Scholar]
- Mudry, M.D. Cytogenetic variability within and across populations of Cebus apella in Argentina. Folia Primatol. (Basel) 1990, 54, 206–216. [Google Scholar] [CrossRef] [PubMed]
- Nieves, M.; Mendez, G.; Ortiz, A.; Mühlmann, M.; Mudry, M.D. Karyological diagnosis of Cebus (Primates, Platyrrhini) in captivity: Detection of hybrids and management program applications. Anim. Reprod. Sci. 2008, 108, 66–78. [Google Scholar] [CrossRef]
- Morrow, E.H.; Gage, M.J.G. Consistent significant variation between individual males in spermatozoal morphometry. J. Zool. 2001, 254, 147–153. [Google Scholar] [CrossRef]
- Gago, C.; Pérez-Sánchez, F.; Yeung, C.H.; Tablado, L.; Cooper, T.G.; Soler, C. Morphological characterization of ejaculated cynomolgus monkey (Macaca fascicularis) sperm. Am. J. Primatol. 1999, 47, 105–115. [Google Scholar] [CrossRef]
- Gravance, C.G.; Champion, Z.J.; Casey, P.J. Computer-assisted sperm head morphometry analysis (ASMA) of cryopreserved ram spermatozoa—ScienceDirect. Theriogenology 1998, 49, 1219–1230. [Google Scholar] [CrossRef]
- Hidalgo, M.; Rodríguez, I.; Dorado, J.; Soler, C. Morphometric classification of Spanish thoroughbred stallion sperm heads. Anim. Reprod. Sci. 2008, 103, 374–378. [Google Scholar] [CrossRef] [PubMed]
- Kahrl, A.F.; Cox, R.M. Consistent Differences in Sperm Morphology and Testis Size between Native and Introduced Populations of Three Anolis Lizard Species. J. Herpetol. 2017, 51, 532–537. [Google Scholar] [CrossRef]
- Maroto-Morales, A.; Ramón, M.; García-Álvarez, O.; Soler, A.J.; Esteso, M.C.; Martínez-Pastor, F.; Pérez-Guzmán, M.D.; Garde, J.J. Characterization of ram (Ovis aries) sperm head morphometry using the Sperm-Class Analyzer. Theriogenology 2010, 73, 437–448. [Google Scholar] [CrossRef]
- Rijsselaere, T.; Van Soom, A.; Hoflack, G.; Maes, D.; de Kruif, A. Automated sperm morphometry and morphology analysis of canine semen by the Hamilton-Thorne analyser. Theriogenology 2004, 62, 1292–1306. [Google Scholar] [CrossRef]
- Buendía, P.; Soler, C.; Paolicchi, F.; Gago, G.; Urquieta, B.; Pérez-Sánchez, F.; Bustos-Obregón, E. Morphometric characterization and classification of alpaca sperm heads using the Sperm-Class Analyzer® computer-assisted system. Theriogenology 2002, 57, 1207–1218. [Google Scholar] [CrossRef]
- Dixson, A.F. Sexual Selection and Evolution of the Seminal Vesicles in Primates. Folia Primatol. (Basel) 1998, 69, 300–306. [Google Scholar] [CrossRef]
- Dixson, A.F.; Anderson, M.J. Sexual Selection, Seminal Coagulation and Copulatory Plug Formation in Primates. Folia Primatol. (Basel) 2002, 73, 63–69. [Google Scholar] [CrossRef]
- Hernández-López, L.; Cerda-Molina, A.L.; Páez-Ponce, L.D.; Mondragón-Ceballos, R. Seasonal emission of seminal coagulum and in vivo sperm dynamics in the black-handed spider monkey (Ateles geoffroyi). Theriogenology 2008, 69, 466–472. [Google Scholar] [CrossRef]
- Silva, K.S.M. Avaliação Andrológica e Criopreservação de Sêmen de Ateles (Macaco-aranha) Mantidos em Cativeiro; Universidade Federal do Pará: Belem, Brazil, 2005. [Google Scholar]
- Dunham, A.E.; Rudolf, V.H.W. Evolution of sexual size monomorphism: The influence of passive mate guarding. J. Evol. Biol. 2009, 22, 1376–1386. [Google Scholar] [CrossRef]
- Campbell, C.J. The taxonomic status of spider monkeys in the twenty-first century. In Spider Monkeys; Cambridge Studies in Biological and Evolutionary Anthropology; Cambridge University Press: Cambridge, UK, 2008; ISBN 978-0-511-72191-5. [Google Scholar]
- Kowalewski, M.M.; Garber, P.A. Mating promiscuity and reproductive tactics in female black and gold howler monkeys (Alouatta caraya) inhabiting an island on the Parana river, Argentina. Am. J. Primatol. 2010, 72, 734–748. [Google Scholar] [CrossRef] [PubMed]
- Belle, S.V.; Bicca-Marques, J.C. Insights into Reproductive Strategies and Sexual Selection in Howler Monkeys. In Howler Monkeys; Kowalewski, M.M., Garber, P.A., Cortés-Ortiz, L., Urbani, B., Youlatos, D., Eds.; Developments in Primatology: Progress and Prospects; Springer: New York, NY, USA, 2015; pp. 57–84. ISBN 978-1-4939-1959-8. [Google Scholar]
- Symington, M.M. Demography, ranging patterns, and activity budgets of black spider monkeys (Ateles paniscus chamek) in the Manu National Park, Peru. Am. J. Primatol. 1988, 15, 45–67. [Google Scholar] [CrossRef]
- Agostini, I.; Visalberghi, E. Social influences on the acquisition of sex-typical foraging patterns by juveniles in a group of wild tufted capuchin monkeys (Cebus nigritus). Am. J. Primatol. 2005, 65, 335–351. [Google Scholar] [CrossRef]
- Bitetti, M.S.D.; Janson, C.H. Reproductive Socioecology of Tufted Capuchins (Cebus apella nigritus) in Northeastern Argentina. Int. J. Primatol. 2001, 22, 127–142. [Google Scholar] [CrossRef]
- Izar, P.; Verderane, M.P.; Peternelli-dos-Santos, L.; Mendonça-Furtado, O.; Presotto, A.; Tokuda, M.; Visalberghi, E.; Fragaszy, D. Flexible and conservative features of social systems in tufted capuchin monkeys: Comparing the socioecology of Sapajus libidinosus and Sapajus nigritus. Am. J. Primatol. 2012, 74, 315–331. [Google Scholar] [CrossRef]
- Digby, L.J. Sexual behavior and extragroup copulations in a wild population of common marmosets (Callithrix jacchus). Folia Primatol. Basel 1999, 70, 136–145. [Google Scholar] [CrossRef] [PubMed]
- Boinski, S. The social organizations of squirrel monkeys: Implications for ecological models of social evolution. Evol. Anthropol. Issues News Rev. 1999, 8, 101–112. [Google Scholar] [CrossRef]
- Wright, P.C. Ecological correlates of monogamy in Aotus and Callicebus. In Primate Ecology and Conservation; Cambridge University Press: Cambridge, UK, 1986; pp. 159–170. [Google Scholar]
- Wright, P.C. The behaviour and ecology of the owl monkey. In Aotus: The Owl Monkey; Academic Press: San Diego, California, USA, 1994; pp. 97–112. [Google Scholar]
- Lüpold, S.; Pitnick, S. Sperm form and function: What do we know about the role of sexual selection? Reproduction 2018, 155, R229–R243. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Specimen Identification | Head Length (Mean ± SD) | Head Width (Mean ± SD) | Acrosome Length (Mean ± SD) | Midpiece Length (Mean ± SD) | Midpiece Width (Mean ± SD) | Tail Length (Mean ± SD) | Total Length (Mean ± SD) | Midpiece Volume (Mean ± SD) |
|---|---|---|---|---|---|---|---|---|
| CCY D000 | 6.16 ± 0.23 | 3.89 ± 0.17 | 3.69 ± 0.21 | 8.67 ± 0.36 | 0.86 ± 0.05 | 59.19 ± 1.32 | 65.36 ± 1.37 | 5.05 ± 0.67 |
| CCY ED08 | 6.07± 0.27 | 3.91 ± 0.23 | 2.89 ± 0.34 | 8.98 ± 0.56 | 0.78 ± 0.08 | 57.94 ± 1.5 | 64.01 ± 1.58 | 4.31 ± 0.97 |
| CCY 4FF6 | 6.20 ± 0.25 | 3.89 ± 0.21 | 3.73 ± 0.31 | 8.36 ± 0.49 | 0.78 ± 0.06 | 59.60 ± 1.63 | 65.80 ± 1.70 | 4.03 ± 0.71 |
| CNI 911F | 6.29 ± 0.19 | 4.11 ± 0.24 | 3.59 ± 0.22 | 9.35 ± 0.44 | 0.84 ± 0.08 | 59.98 ± 0.97 | 66.27 ± 0.10 | 5.20 ± 1.05 |
| ACA C73C | 4.86 ± 0.23 | 3.39 ± 0.23 | 2.32 ± 0.14 | 3.43 ± 0.28 | 0.99 ± 0.09 | 44.78 ± 2.63 | 49.64 ± 2.70 | 2.65 ± 0.54 |
| ACA ADD7 | 5.01 ± 0.20 | 3.32 ± 0.15 | 2.41 ± 0.17 | 3.54 ± 0.24 | 0.90 ± 0.08 | 45.54 ± 1.76 | 50.55 ± 1.80 | 2.29 ± 0.45 |
| ACH 44AF | 5.16 ± 0.24 | 3.27 ± 0.17 | 2.51 ± 0.25 | 7.34 ± 0.33 | 0.97 ± 0.09 | 58.93 ± 2.01 | 64.09 ± 2.03 | 5.42 ± 1.08 |
| ABE C97C | 5.20 ± 0.17 | 2.70 ± 0.11 | 2.27 ± 0.12 | 6.56 ± 0.24 | 0.83 ± 0.06 | 65.63 ± 1.45 | 70.83 ± 1.47 | 3.53 ± 0.55 |
| Species | Head Length (µm) | Head Width (µm) | Acrosome Length (µm) | Midpiece Length (µm) | Total Length (µm) | Midpiece Volume (µm3) | Ellipticity (HL/HW) | Elongation (HL − HW)/(HL + HW) | Number of Specimens Analyzed | References |
|---|---|---|---|---|---|---|---|---|---|---|
| Cebidae familiy | ||||||||||
| Callithrix jacchus | 5 | 4.4 | 50 | 1 | [9] | |||||
| C. jacchus | 5 | 3 | 50 | 1.67 | 0.25 | 1 | [2] | |||
| C. jacchus | 5.6 | 3.9 | 48.2 | 0.6 | [11] | |||||
| C. jacchus | 5.19 | 3.46 | 1. 5 | 0.2 | 5 | [21] | ||||
| C. jacchus | 5.19 | 3.46 | 1.5 | 0.2 | 4 | [24] | ||||
| Callithrix jacchus Mean * | 5.20 | 3.31 | 4.15 | 49.4 | 1.58 | 0.22 | ||||
| C. pygmaea | 4.6 | 8.5 | 45.5 | 3.3 | 1 | [11] | ||||
| Callimico goeldii | 4.95 | 3.14 | 1.58 | 0.22 | 5 | [22] | ||||
| C. goeldii | 4.95 | 3.14 | 1.58 | 0.22 | 5 | [21] | ||||
| Callimico goeldii Mean * | 4.95 | 3.14 | 1.58 | 0.22 | ||||||
| Saguinus midas | 5.3 | 9.1 | 45.5 | 10.2 | 1 | [11] | ||||
| S. oedipus | 4.7 | 1.2 | 43.6 | 8.9 | 1 | [11] | ||||
| Aotus lemurinus | 4.7 | 2.8 | 6.6 | 55.1 | 1.68 | 0.25 | 3 | [18] | ||
| Saimiri boliviensis | 6.1 | 4.3 | 10.5 | 76.8 | 1.42 | 0.17 | 1 | [18] | ||
| S. boliviensis | 5.71 | 3.76 | 3.7 | 12.21 | 71.39 | 1.52 | 0.21 | 2 | [20] | |
| Saimiri boliviensis Mean * | 5.91 | 4.03 | 3.70 | 11.36 | 74.10 | 1.46 | 0.19 | |||
| S. sciureus | 5.6 | 10.1 | 71.1 | 6.3 | 1 | [11] | ||||
| S. sciureus | 5.11 | 3.51 | 9.03 | 69.24 | 1.45 | 0.19 | 1 | [17] | ||
| Saimiri sciureus Mean * | 5.36 | 3.51 | 9.57 | 70.17 | 1.52 | 0.21 | ||||
| S. collinsi | 6.2 | 4.3 | 76.7 | 1.44 | 0.18 | 10 | [19] | |||
| S. vanzolinii | 6.8 | 4.8 | 76.1 | 1.42 | 0.17 | 2 | [19] | |||
| Saimiri sp. Mean * | 5.92 | 4.13 | 3.70 | 10.46 | 73.56 | 1.43 | 0.18 | |||
| Cebus nigritus | 5.68 | 3.75 | 9.29 | 67.83 | 1.51 | 0.21 | 1 | [20] | ||
| C. nigritus | 6.29 | 4.11 | 3.59 | 9.35 | 66.27 | 5.2 | 1.53 | 0.21 | 1 | This study |
| Cebus nigritus Mean * | 5.99 | 3.93 | 3.59 | 9.32 | 67.05 | 1.52 | 0.21 | |||
| Cebus cay (ex C. paraguayanus) | 5.87 | 3.72 | 8.53 | 67.26 | 1.58 | 0.22 | 1 | [20] | ||
| C. cay | 6.14 | 3.90 | 3.44 | 8.67 | 65.06 | 5.05 | 1.58 | 0.22 | 3 | This study |
| Cebus cay Mean * | 6.01 | 3.81 | 3.44 | 8.60 | 66.16 | 1.58 | 0.22 | |||
| C. albifrons | 9 | 13.2 | 67.2 | 8.4 | 1 | [11] | ||||
| C. apella | 8.5 | 11.6 | 72.7 | 4.5 | 1 | [11] | ||||
| Cebus sp. Mean * | 6.47 | 3.88 | 3.49 | 9.37 | 66.91 | 1.67 | 0.25 | |||
| Cebidae Mean * | 5.71 | 3.70 | 3.52 | 8.96 | 61.45 | 1.54 | 0.21 | |||
| Atelidae family | ||||||||||
| Alouatta caraya | 5 | 3.09 | 3.63 | 53.93 | 1.62 | 0.24 | 1 | [20] | ||
| A. caraya | 4.66 | 2.94 | 1.58 | 0.23 | 5 | [21] | ||||
| A. caraya | 4.93 | 3.35 | 2.36 | 3.48 | 50.09 | 2.65 | 1.47 | 0.19 | 2 | This study |
| A. caraya | 4.66 | 2.94 | 1.58 | 0.23 | 5 | [23] | ||||
| Alouatta caraya Mean * | 4.81 | 3.08 | 2.36 | 3.56 | 52.01 | 1.56 | 0.22 | |||
| Ateles paniscus | 5.11 | 3.27 | 7.22 | 67.26 | 1.56 | 0.22 | 1 | [20] | ||
| A. paniscus | 6.4 | 10.2 | 54.9 | 13.2 | 1 | [11] | ||||
| Ateles paniscus Mean * | 5.75 | 3.27 | 8.71 | 61.08 | 13.2 | 1.56 | 0.28 | |||
| A. chamek | 5.16 | 3.27 | 2.51 | 7.34 | 64.09 | 5.42 | 1.58 | 0.22 | 1 | This study |
| A. belzebuth | 5.2 | 2.7 | 2.27 | 6.56 | 70.83 | 3.53 | 1.92 | 0.32 | 1 | This study |
| A. geoffroyi | 4.51 | 2.78 | 1.62 | 0.24 | 5 | [21] | ||||
| Ateles sp. Mean * | 5.00 | 3.01 | 2.39 | 7.04 | 67.39 | 1.66 | 0.27 | |||
| Atelidae Mean * | 4.91 | 3.08 | 2.38 | 5.29 | 59.38 | 1.65 | 0.25 | |||
| Total | 75 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Steinberg, E.R.; Sestelo, A.J.; Ceballos, M.B.; Wagner, V.; Palermo, A.M.; Mudry, M.D. Sperm Morphology in Neotropical Primates. Animals 2019, 9, 839. https://doi.org/10.3390/ani9100839
Steinberg ER, Sestelo AJ, Ceballos MB, Wagner V, Palermo AM, Mudry MD. Sperm Morphology in Neotropical Primates. Animals. 2019; 9(10):839. https://doi.org/10.3390/ani9100839
Chicago/Turabian StyleSteinberg, Eliana R., Adrián J. Sestelo, María B. Ceballos, Virginia Wagner, Ana M. Palermo, and Marta D. Mudry. 2019. "Sperm Morphology in Neotropical Primates" Animals 9, no. 10: 839. https://doi.org/10.3390/ani9100839