Effects of Dietary Supplementation of Alpha-Ketoglutarate in a Low-Protein Diet on Fatty Acid Composition and Lipid Metabolism Related Gene Expression in Muscles of Growing Pigs

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals, Experimental Design, and Diets
2.2. Sample Collection and Preparation
2.3. Measurement of Serum Lipid Related Substances
2.4. Muscle Fatty Acid Composition
2.5. RNA Extraction, Complementary DNA Synthesis, and Quantitative RT-PCR
2.6. Statistical Analysis
3. Results
3.1. Serum Biochemical Parameters
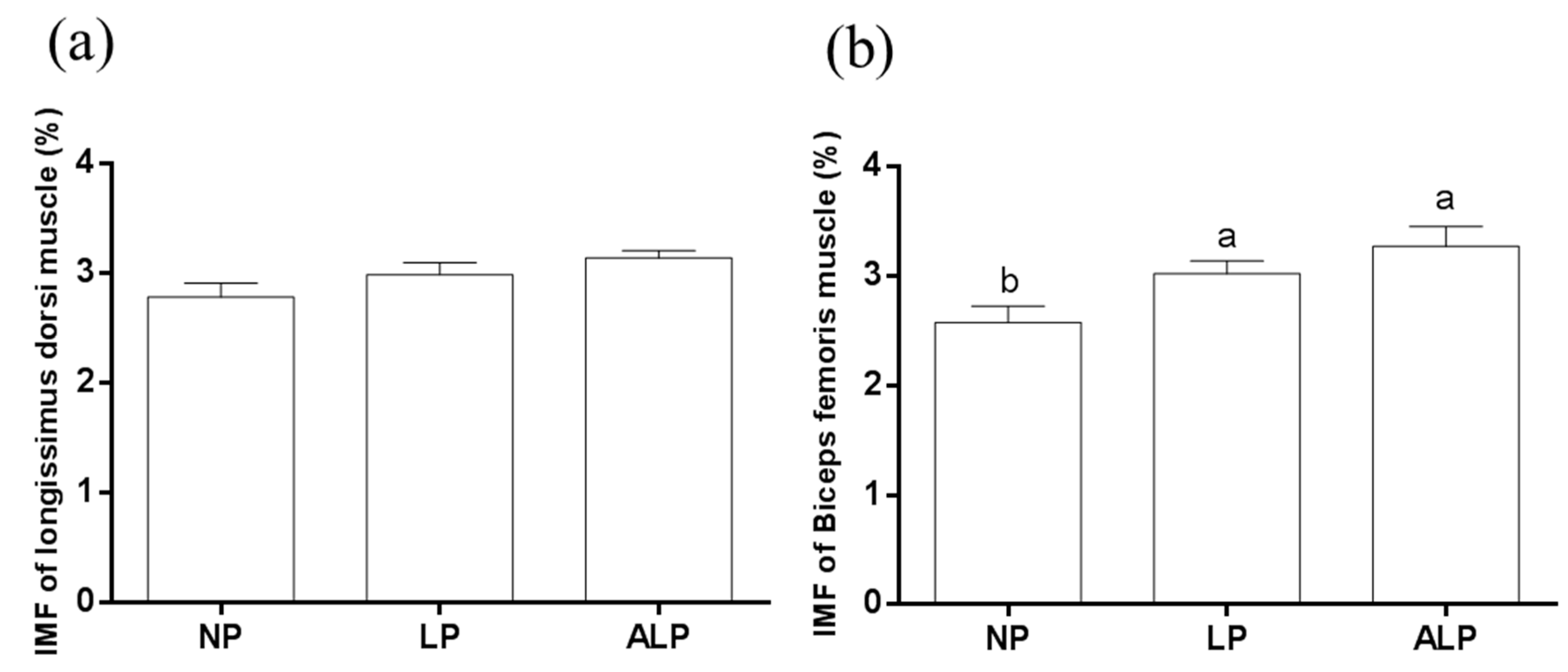
3.2. Intramuscular Fat and Fatty Acid Profile of Muscle
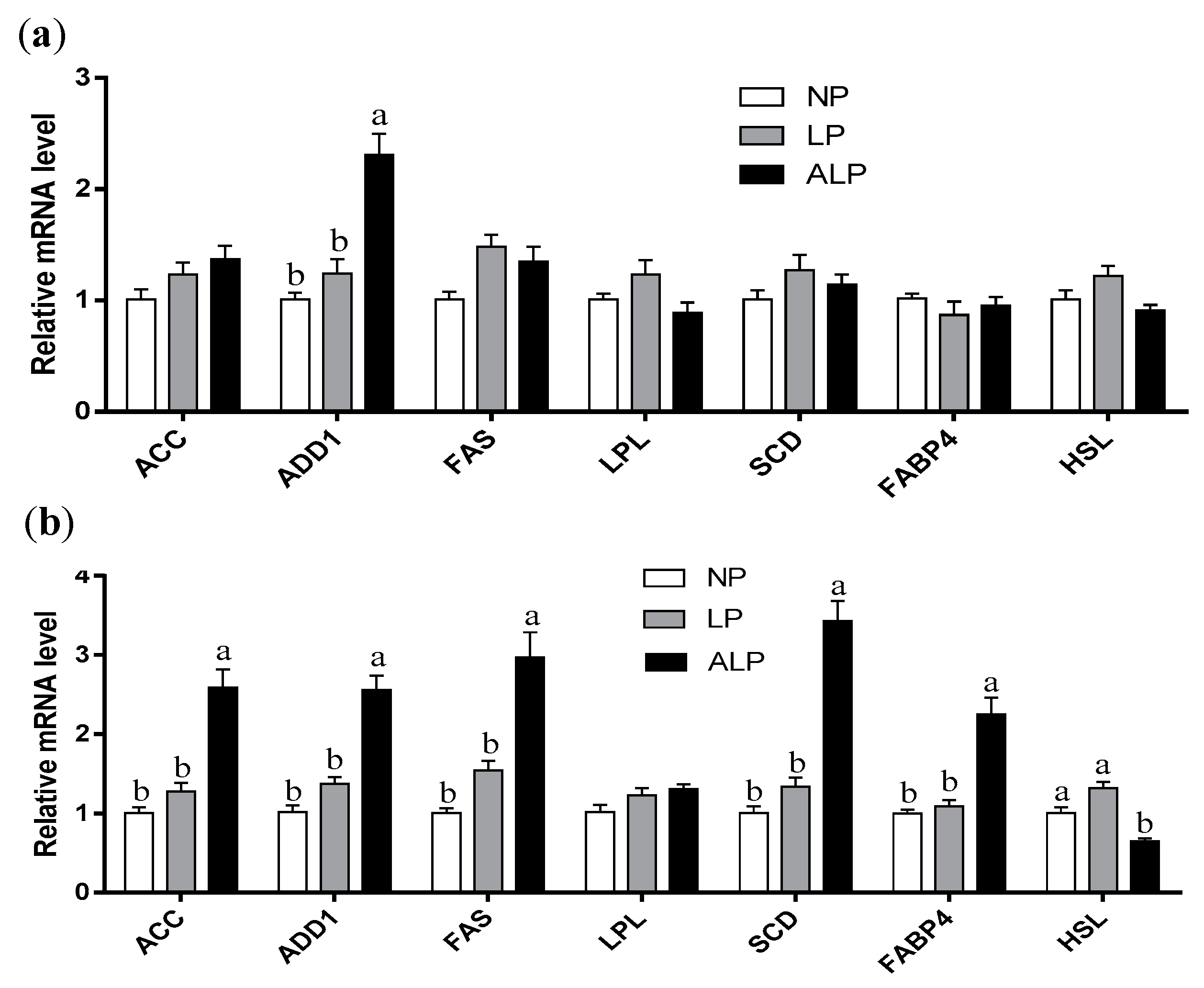
3.3. Gene mRNA Expression Levels in Muscle
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Dannenberger, D.; Nuernberg, K.; Nuernberg, G.; Priepke, A. Impact of dietary protein level and source of polyunsaturated fatty acids on lipid metabolism-related protein expression and fatty acid concentrations in porcine tissues. J. Agric. Food Chem. 2014, 62, 12453–12461. [Google Scholar] [CrossRef] [PubMed]
- Kouba, M.; Bonneau, M. Compared development of intermuscular and subcutaneous fat in carcass and primal cuts of growing pigs from 30 to 140 kg body weight. Meat Sci. 2009, 81, 270–274. [Google Scholar] [CrossRef] [PubMed]
- Wood, J.D.; Nute, G.R.; Richardson, R.I.; Whittington, F.M.; Southwood, O.; Plastow, G.; Mansbridge, R.; da Costa, N.; Chang, K.C. Effects of breed, diet and muscle on fat deposition and eating quality in pigs. Meat Sci. 2004, 67, 651–667. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, X.; Monin, G.; Talmant, A.; Mourot, J.; Lebret, B. Influence of intramuscular fat content on the quality of pig meat-1. Composition of the lipid fraction and sensory characteristics of m. longissimus lumborum. Meat Sci. 1999, 53, 59–65. [Google Scholar] [CrossRef]
- Ye, C.; Zeng, X.; Zhu, J.; Liu, Y.; Ye, Q.; Qiao, S.; Zeng, X. Dietary N-Carbamylglutamate Supplementation in a Reduced Protein Diet Affects Carcass Traits and the Profile of Muscle Amino Acids and Fatty Acids in Finishing Pigs. J. Agric. Food Chem. 2017, 65, 5751–5758. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.; Li, F.; Guo, Q.; Wang, W.; Zhang, L.; Wen, C.; Yin, Y. Branched-chain amino acid ratios modulate lipid metabolism in adipose tissues of growing pigs. J. Funct. Foods 2018, 40, 614–624. [Google Scholar] [CrossRef]
- Apple, J.K.; Maxwell, C.V.; Bass, B.E.; Yancey, J.W.S.; Payne, R.L.; Thomson, J. Effects of reducing dietary crude protein levels and replacement with crystalline amino acids on growth performance, carcass composition, and fresh pork quality of finishing pigs fed ractopamine hydrochloride. J. Anim. Sci. 2017, 95, 4971–4985. [Google Scholar] [CrossRef] [Green Version]
- Radzki, R.P.; Bienko, M.; Pierzynowski, S.G. Effect of dietary alpha-ketoglutarate on blood lipid profile during hypercholesterolaemia in rats. Scand. J. Clin. Lab. Investig. 2009, 69, 175–180. [Google Scholar] [CrossRef]
- Selvaraj, V.; Dakshayani, K.B.; Perumal, S. Effects of alpha-ketoglutarate on antioxidants and lipid peroxidation products in rats treated with ammonium acetate. Nutrition 2002, 18, 747–750. [Google Scholar]
- Junghans, P.; Derno, M.; Pierzynowski, S.; Hennig, U.; Eberhard Rudolph, P.; Souffrant, W.B. Intraduodenal infusion of alpha-ketoglutarate decreases whole body energy expenditure in growing pigs. Clin. Nutr. 2006, 25, 489–496. [Google Scholar] [CrossRef]
- Hou, Y.; Yao, K.; Wang, L.; Ding, B.; Fu, D.; Liu, Y.; Zhu, H.; Liu, J.; Li, Y.; Kang, P.; et al. Effects of α-ketoglutarate on energy status in the intestinal mucosa of weaned piglets chronically challenged with lipopolysaccharide. Br. J. Nutr. 2011, 106, 357–363. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Yi, D.; Hou, Y.; Ding, B.; Li, K.; Li, B.; Zhu, H.; Liu, Y.; Wu, G. Dietary Supplementation with alpha-Ketoglutarate Activates mTOR Signaling and Enhances Energy Status in Skeletal Muscle of Lipopolysaccharide-Challenged Piglets. J. Nutr. 2016, 146, 1514–1520. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Hou, Y.; Yi, D.; Li, Y.; Ding, B.; Zhu, H.; Liu, J.; Xiao, H.; Wu, G. Dietary supplementation with glutamate precursor α-ketoglutarate attenuates lipopolysaccharide-induced liver injury in young pigs. Amino Acids 2015, 47, 1309–1318. [Google Scholar] [CrossRef] [PubMed]
- Yao, K.; Xiong, X.; Fu, C.; Dahanayaka, S.; Lei, J.; Lu, W.; Wu, G.; Liao, P.; Yin, Y. Dietary alpha-ketoglutarate supplementation may increase muscle gain through mTOR signaling pathway in diet-induced obese rat. J. Food Agric. Environ. 2013, 11, 151–154. [Google Scholar]
- Chen, J.; Kang, B.; Jiang, Q.; Han, M.; Zhao, Y.; Long, L.; Fu, C.; Yao, K. Alpha-ketoglutarate in low-Protein diets for growing pigs: Effects on cecal microbial communities and parameters of microbial metabolism. Front. Microbiol. 2018, 9, 1057. [Google Scholar] [CrossRef]
- Chen, J.; Su, W.; Kang, B.; Jiang, Q.; Zhao, Y.; Fu, C.; Yao, K. Supplementation with alpha-ketoglutarate to a low-protein diet enhances amino acid synthesis in tissues and improves protein metabolism in the skeletal muscle of growing pigs. Amino Acids 2018, 50, 1525–1537. [Google Scholar] [CrossRef]
- Tan, B.; Yin, Y.; Liu, Z.; Tang, W.; Xu, H.; Kong, X.; Li, X.; Yao, K.; Gu, W.; Smith, S.B.; et al. Dietary L-arginine supplementation differentially regulates expression of lipid-metabolic genes in porcine adipose tissue and skeletal muscle. J. Nutr. Biochem. 2011, 22, 441–445. [Google Scholar] [CrossRef]
- National Research Council. Nutrient Requirements of Swine, 11th ed.; National Academic Press: Washington, DC, USA, 2012. [Google Scholar]
- Chen, C.; Chen, J.; Wang, S.; You, C.; Li, Y. Effects of different dietary ratios of linolenic to linoleic acids or docosahexaenoic to eicosapentaenoic acids on the growth and immune indices in grouper, Epinephelus coioides. Aquaculture 2017, 473, 153–160. [Google Scholar] [CrossRef]
- Li, Y.Y.; Hu, C.B.; Zheng, Y.J.; Xia, X.A.; Xu, W.J.; Wang, S.Q.; Chen, W.Z.; Sun, Z.W.; Huang, J.H. The effects of dietary fatty acids on liver fatty acid composition and Δ 6 -desaturase expression differ with ambient salinities in Siganus canaliculatus. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2008, 151, 183–190. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C (T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Luo, Y.; Zhang, X.; Zhu, Z.; Jiao, N.; Qiu, K.; Yin, J. Surplus dietary isoleucine intake enhanced monounsaturated fatty acid synthesis and fat accumulation in skeletal muscle of finishing pigs. J. Anim. Sci. Biotechnol. 2018, 9, 88. [Google Scholar] [CrossRef] [PubMed]
- Franco, D.; Vazquez, J.A.; Lorenzo, J.M. Growth performance, carcass and meat quality of the Celta pig crossbred with Duroc and Landrance genotypes. Meat Sci. 2014, 96, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Wood, J.; Enser, M.; Fisher, A.; Nute, G.; Sheard, P.; Richardson, R.I.; Hughes, S.; Whittington, F. Fat deposition, fatty acid composition and meat quality: A review. Meat Sci. 2008, 78, 343–358. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.; Liu, Z.; Zhou, Y.; Wei, H.; Zhang, X.; Xia, M.; Deng, Z.; Zou, Y.; Jiang, S.; Peng, J. Effect of oregano essential oil supplementation to a reduced-protein, amino acid-supplemented diet on meat quality, fatty acid composition, and oxidative stability of Longissimus thoracis muscle in growing-finishing pigs. Meat Sci. 2017, 133, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Wood, J.D.; Lambe, N.R.; Walling, G.A.; Whitney, H.; Jagger, S.; Fullarton, P.J.; Bayntun, J.; Hallett, K.; Bünger, L. Effects of low protein diets on pigs with a lean genotype. 1. Carcass composition measured by dissection and muscle fatty acid composition. Meat Sci. 2013, 95, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.; Duan, Y.; Li, F.; Li, Y.; Guo, Q.; Ji, Y.; Tan, B.; Li, T.; Yin, Y. Effects of supplementation with branched-chain amino acids to low-protein diets on expression of genes related to lipid metabolism in skeletal muscle of growing pigs. Amino Acids 2016, 48, 2131–2144. [Google Scholar] [CrossRef] [PubMed]
- Doran, O.; Moule, S.K.; Teye, G.A.; Whittington, F.M.; Hallett, K.G.; Wood, J.D. A reduced protein diet induces stearoyl-CoA desaturase protein expression in pig muscle but not in subcutaneous adipose tissue: Relationship with intramuscular lipid formation. Br. J. Nutr. 2006, 95, 609–617. [Google Scholar] [CrossRef]
- Bie, T.; Yin, Y.; Liu, Z.; Li, X.; Xu, H.; Kong, X.; Huang, R.; Tang, W.; Shinzato, I.; Smith, S.B. Dietaryl-arginine supplementation increases muscle gain and reduces body fat mass in growing-finishing pigs. Amino Acids 2009, 37, 169–175. [Google Scholar]
- Cameron, N.D.; Enser, M.; Nute, G.R.; Whittington, F.M.; Penman, J.C.; Fisken, A.C.; Perry, A.M.; Wood, J.D. Genotype with nutrition interaction on fatty acid composition of intramuscular fat and the relationship with flavour of pig meat. Meat Sci. 2000, 55, 187–195. [Google Scholar] [CrossRef]
- Realini, C.E.; Duran-Montge, P.; Lizardo, R.; Gispert, M.; Oliver, M.A.; Esteve-Garcia, E. Effect of source of dietary fat on pig performance, carcass characteristics and carcass fat content, distribution and fatty acid composition. Meat Sci. 2010, 85, 606–612. [Google Scholar] [CrossRef]
- Hu, F.B.; Stampfer, M.J.; Manson, J.E.; Ascherio, A.; Colditz, G.A.; Speizer, F.E.; Hennekens, C.H.; Willett, W.C. Dietary saturated fats and their food sources in relation to the risk of coronary heart disease in women. Am. J. Clin. Nutr. 1999, 70, 1001–1008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kristensen, N.B.; Jungvid, H.; Fernandez, J.A.; Pierzynowski, S.G.; Kristensen, N.B.; Jungvid, H.; Fernandez, J.A.; Pierzynowski, S.G. Absorption and metabolism of α-etoglutarate in growing pigs. J. Anim. Physiol. Anim. Nutr. 2002, 86, 239–245. [Google Scholar] [CrossRef] [PubMed]
- Kimball, S.R.; Jefferson, L.S. Signaling pathways and molecular mechanisms through which branched-chain amino acids mediate translational control of protein synthesis. J. Nutr. 2006, 136, 227–231. [Google Scholar] [CrossRef] [PubMed]
- Estany, J.; Ros-Freixedes, R.; Tor, M.; Pena, R.N. A functional variant in the stearoyl-CoA desaturase gene promoter enhances fatty acid desaturation in pork. PLoS ONE 2014, 9, e86177. [Google Scholar] [CrossRef]
- Ntambia, J.M.; Miyazakib, M. Regulation of stearoyl-CoA desaturases and role in metabolism. Prog. Lipid. Res. 2004, 43, 91–104. [Google Scholar] [CrossRef]
- Sul, H.S.; Wang, D. Nutritional and hormonal regulation of enzymes in fat synthesis: Studies of fatty acid synthase and mitochondrial glycerol-3-phosphate acyltransferase gene transcription. Annu. Rev. Nutr. 1998, 18, 331–351. [Google Scholar] [CrossRef]
- Moldes, M.; Boizard, M.; Liepvre, X.L.; Fève, B.; Dugail, I.; Pairault, J. Functional antagonism between inhibitor of DNA binding (Id) and adipocyte determination and differentiation factor 1/sterol regulatory element-binding protein-1c (ADD1/SREBP-1c) trans-factors for the regulation of fatty acid synthase promoter in adipocytes. Biochem. J. 1999, 344, 873–880. [Google Scholar] [CrossRef]
- Horton, J.D.; Goldstein, J.L.; Brown, M.S. SREBPs: Activators of the complete program of cholesterol and fatty acid synthesis in the liver. J. Clin. Investig. 2002, 109, 1125–1131. [Google Scholar] [CrossRef]
- Munday, M.R. Regulation of mammalian acetyl-CoA carboxylase. Biochem. Soc. Trans. 2002, 30, 1059–1064. [Google Scholar] [CrossRef]
- Holm, C. Molecular mechanisms regulating hormone-sensitive lipase and lipolysis. Biochem. Soc. Trans. 2003, 31, 1120–1124. [Google Scholar] [CrossRef]
- Berraondo, B.; Martínez, J.A. Free fatty acids are involved in the inverse relationship between hormone-sensitive lipase (HSL) activity and expression in adipose tissue after high-fat feeding or β 3-adrenergic stimulation. Obes. Res. 2012, 8, 255–261. [Google Scholar] [CrossRef] [PubMed]
- Hocquette, J.F.; Gondret, F.; Baéza, E.; Médale, F.; Jurie, C.; Pethick, D.W. Intramuscular fat content in meat-producing animals: Development, genetic and nutritional control, and identification of putative markers. Animal 2010, 4, 303–319. [Google Scholar] [CrossRef] [PubMed]
- Madeira, M.S.; Pires, V.M.R.; Alfaia, C.M.; Costa, A.S.H.; Luxton, R.; Doran, O.; Bessa, R.J.B.; Prates, J.A.M. Differential effects of reduced protein diets on fatty acid composition and gene expression in muscle and subcutaneous adipose tissue of Alentejana purebred and Large White×Landrace×Pietrain crossbred pigs. Br. J. Nutr. 2013, 110, 216–229. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Items | Dietary Treatment a | ||
|---|---|---|---|
| NP | LP | ALP | |
| Ingredient (%) | |||
| Corn | 63.64 | 66.50 | 65.50 |
| Soybean meal | 19.80 | 18.80 | 18.80 |
| Dried whey | 4.30 | 4.30 | 4.30 |
| Fish meal | 9.00 | 4.00 | 4.00 |
| Soybean oil | 0.80 | 2.60 | 2.60 |
| AKG b | 0.00 | 0.00 | 1.00 |
| Limestone | 0.50 | 0.60 | 0.60 |
| Monocalcium phosphate | 0.00 | 0.74 | 0.74 |
| L-lysine-HCl | 0.41 | 0.65 | 0.65 |
| L-threonine | 0.11 | 0.25 | 0.25 |
| DL-methionine | 0.13 | 0.20 | 0.20 |
| L-tryptophan | 0.01 | 0.06 | 0.06 |
| Sodium chloride | 0.30 | 0.30 | 0.30 |
| Premix c | 1.00 | 1.00 | 1.00 |
| Total | 100.00 | 100.00 | 100.00 |
| Nutrient level (%) | |||
| Digestible energy (MJ/kg) d | 14.60 | 14.60 | 14.60 |
| Crude protein | 20.19 | 17.21 | 17.20 |
| Lysine | 1.25 | 1.24 | 1.23 |
| Methionine | 0.38 | 0.37 | 0.37 |
| Methionine + cysteine | 0.62 | 0.65 | 0.63 |
| Threonine | 0.76 | 0.73 | 0.74 |
| Tryptophan | 0.22 | 0.22 | 0.21 |
| Total calcium | 0.72 | 0.72 | 0.71 |
| Total phosphorus | 0.65 | 0.64 | 0.63 |
| Gene | Primer Sequence (5′-3′) | Size (bp) | Accession No. |
|---|---|---|---|
| ACC | F: GGCCATCAAGGACTTCAACC | 120 | NM_001114269 |
| R: ACGATGTAAGCGCCGAACTT | |||
| ADD1 | F: GCGACGGTGCCTCTGGTAGT | 218 | XM_021066226.1 |
| R: CGCAAGACGGCGGATTTA | |||
| FAS | F: GCCTAACTCCTCGCTGCAAT | 195 | NM_001099930.1 |
| R: TCCTTGGAACCGTCTGTGTTC | |||
| LPL | F: CTCGTGCTCAGATGCCCTAC | 148 | NM_214286.1 |
| R: GGCAGGGTGAAAGGGATGTT | |||
| SCD | F: GCCTACTATCTGCTGAGTGC | 152 | XM_021072070.1 |
| R: TCTCGGGCCCATTCATAAAC | |||
| FABP4 | F: TGGAAACTTGTCTCCAGTG | 147 | NM_001002817.1 |
| R: GGTACTTTCTGATCTAATGGTG | |||
| HSL | F: TCGGAGTGAACGGATTTG | 195 | - |
| R: TCCTCCTTGGTGCTAATCTCGT | |||
| GAPDH | F: TCGGAGTGAACGGATTTG | 219 | NM_001206359.1 |
| R: CCTGGAAGATGGTGATGG |
| Items | Dietary Treatment 1 | SEM | p-Value | ||
|---|---|---|---|---|---|
| NP | LP | ALP | |||
| Non-esterified fatty acid, μmol/L | 610.43 | 578.68 | 605.74 | 11.81 | 0.261 |
| HDL-cholesterol, mmol/L | 0.78 b | 0.84 a,b | 0.96 a | 0.12 | 0.032 |
| LDL-cholesterol, mmol/L | 1.47 | 1.36 | 1.32 | 0.03 | 0.074 |
| Triglyceride, mmol/L | 0.46 a | 0.38 a | 0.24 b | 0.04 | 0.028 |
| TC, mmol/L | 2.96 a | 2.88 a | 2.73 b | 0.05 | 0.039 |
| Items | Dietary Treatment 1 | SEM | p-Value | ||
|---|---|---|---|---|---|
| NP | LP | ALP | |||
| C12:0 | 0.06 | 0.07 | 0.06 | 0.02 | 0.265 |
| C13:0 | 0.19 | 0.17 | 0.16 | 0.06 | 0.704 |
| C14:0 | 0.93 | 0.90 | 0.84 | 0.11 | 0.586 |
| C15:0 | 0.22 | 0.19 | 0.17 | 0.02 | 0.779 |
| C15:1 | 0.16 | 0.16 | 0.15 | 0.03 | 0.925 |
| C16:0 | 22.14 | 21.97 | 21.13 | 1.71 | 0.553 |
| C16:1 | 1.89 | 1.84 | 2.29 | 0.20 | 0.095 |
| C17:0 | 1.15 | 1.05 | 0.86 | 0.11 | 0.086 |
| C18:0 | 12.44 | 11.61 | 11.34 | 0.26 | 0.812 |
| C18:1n-9 | 30.13 | 30.99 | 31.77 | 2.21 | 0.115 |
| C18:2n-6 | 16.33 | 16.04 | 15.59 | 1.13 | 0.269 |
| C20:0 | 0.19 | 0.18 | 0.20 | 0.03 | 0.662 |
| C18:3n-3 | 0.51 | 0.50 | 0.48 | 0.07 | 0.238 |
| C20:2n-6 | 0.59 | 0.61 | 0.59 | 0.14 | 0.146 |
| C20:3n-6 | 0.33 | 0.30 | 0.28 | 0.06 | 0.804 |
| C20:3n-3 | 0.15 | 0.14 | 0.12 | 0.05 | 0.173 |
| C20:4n-6 | 1.72 | 1.66 | 1.60 | 0.19 | 0.147 |
| C20:5n-3 | 0.16 | 0.15 | 0.14 | 0.10 | 0.216 |
| C22:6n-3 | 0.13 | 0.13 | 0.12 | 0.29 | 0.418 |
| SFA 2 | 37.30 | 36.15 | 34.76 | 2.75 | 0.352 |
| MUFA 3 | 32.17 b | 32.99 a,b | 34.21 a | 2.34 | 0.049 |
| PUFA 4 | 19.91 | 19.54 | 18.91 | 1.07 | 0.526 |
| ΣPUFA:SFA | 0.53 | 0.54 | 0.54 | 0.09 | 0.599 |
| Σn-3 PUFA 5 | 0.94 | 0.93 | 0.86 | 0.30 | 0.198 |
| Σn-6 PUFA 6 | 18.97 | 18.61 | 18.05 | 1.17 | 0.371 |
| Σn-6:n-3 PUFA | 20.19 | 20.05 | 21.00 | 0.84 | 0.642 |
| Items | Dietary Treatment 1 | SEM | p-Value | ||
|---|---|---|---|---|---|
| NP | LP | ALP | |||
| C12:0 | 0.06 | 0.06 | 0.07 | 0.02 | 0.667 |
| C13:0 | 0.26 | 0.29 | 0.22 | 0.09 | 0.460 |
| C14:0 | 0.82 | 0.80 | 0.78 | 0.08 | 0.851 |
| C15:0 | 0.22 | 0.23 | 0.30 | 0.08 | 0.636 |
| C15:1 | 0.16 | 0.18 | 0.13 | 0.06 | 0.502 |
| C16:0 | 21.36 | 20.75 | 20.52 | 0.60 | 0.563 |
| C16:1 | 1.91 | 1.66 | 2.00 | 0.21 | 0.097 |
| C17:0 | 1.01 | 1.08 | 0.98 | 0.36 | 0.495 |
| C18:0 | 10.65 | 11.37 | 11.84 | 0.99 | 0.077 |
| C18:1n-9 | 27.67 b | 28.54 b | 31.94 a | 0.65 | 0.046 |
| C18:2n-6 | 19.62 | 19.61 | 18.38 | 0.39 | 0.119 |
| C20:0 | 0.15 | 0.15 | 0.17 | 0.03 | 0.286 |
| C18:3n-3 | 1.02 | 0.94 | 0.83 | 0.11 | 0.256 |
| C20:2n-6 | 0.75 | 0.71 | 0.67 | 0.09 | 0.243 |
| C20:3n-6 | 0.38 a | 0.35 a,b | 0.24 b | 0.13 | 0.041 |
| C20:3n-3 | 0.15 | 0.14 | 0.13 | 0.05 | 0.203 |
| C20:4n-6 | 2.40 | 2.27 | 2.11 | 0.13 | 0.477 |
| C20:5n-3 | 0.24 | 0.23 | 0.20 | 0.05 | 0.219 |
| C22:6n-3 | 0.18 | 0.17 | 0.17 | 0.02 | 0.312 |
| SFA 2 | 34.54 | 34.74 | 34.89 | 2.30 | 0.165 |
| MUFA 3 | 29.73 b | 30.38 b | 34.07 a | 1.26 | 0.038 |
| PUFA 4 | 24.74 | 24.42 | 22.73 | 1.39 | 0.219 |
| ΣPUFA:SFA | 0.72 | 0.70 | 0.65 | 0.07 | 0.104 |
| Σn-3 PUFA 5 | 1.58 | 1.48 | 1.33 | 0.03 | 0.135 |
| Σn-6 PUFA 6 | 23.15 | 22.94 | 21.40 | 2.44 | 0.132 |
| Σn-6:n-3 PUFA | 14.61 b | 15.49 a,b | 16.08 a | 1.51 | 0.026 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, J.; Zhang, H.; Gao, H.; Kang, B.; Chen, F.; Li, Y.; Fu, C.; Yao, K. Effects of Dietary Supplementation of Alpha-Ketoglutarate in a Low-Protein Diet on Fatty Acid Composition and Lipid Metabolism Related Gene Expression in Muscles of Growing Pigs. Animals 2019, 9, 838. https://doi.org/10.3390/ani9100838
Chen J, Zhang H, Gao H, Kang B, Chen F, Li Y, Fu C, Yao K. Effects of Dietary Supplementation of Alpha-Ketoglutarate in a Low-Protein Diet on Fatty Acid Composition and Lipid Metabolism Related Gene Expression in Muscles of Growing Pigs. Animals. 2019; 9(10):838. https://doi.org/10.3390/ani9100838
Chicago/Turabian StyleChen, Jiashun, Haihan Zhang, Hu Gao, Baoju Kang, Fengming Chen, Yinghui Li, Chenxing Fu, and Kang Yao. 2019. "Effects of Dietary Supplementation of Alpha-Ketoglutarate in a Low-Protein Diet on Fatty Acid Composition and Lipid Metabolism Related Gene Expression in Muscles of Growing Pigs" Animals 9, no. 10: 838. https://doi.org/10.3390/ani9100838