
An Overview of Reactive Oxygen Species Damage Occurring during In Vitro Bovine Oocyte and Embryo Development and the Efficacy of Antioxidant Use to Limit These Adverse Effects

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
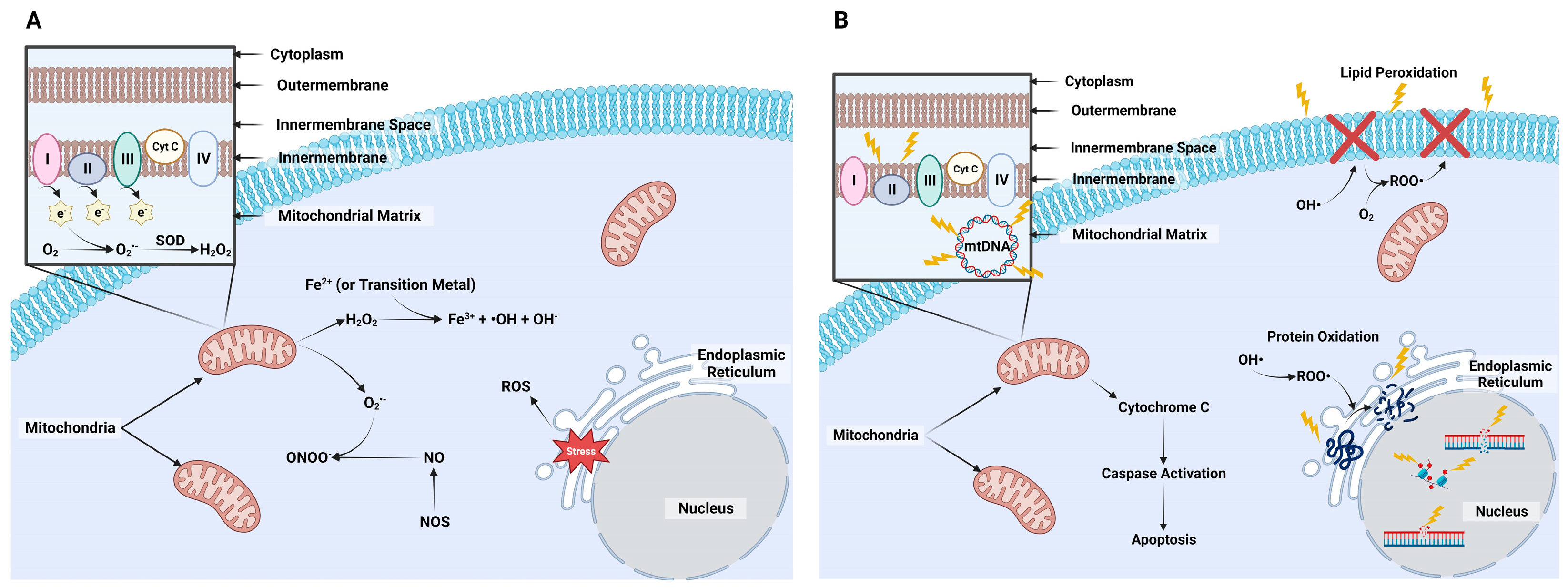
2. Formation of Reactive Species
2.1. Reactive Oxygen Species
2.2. Nitrogen Species
2.3. Detrimental Effects
2.4. Accumulation of Reactive Species with In Vitro Production
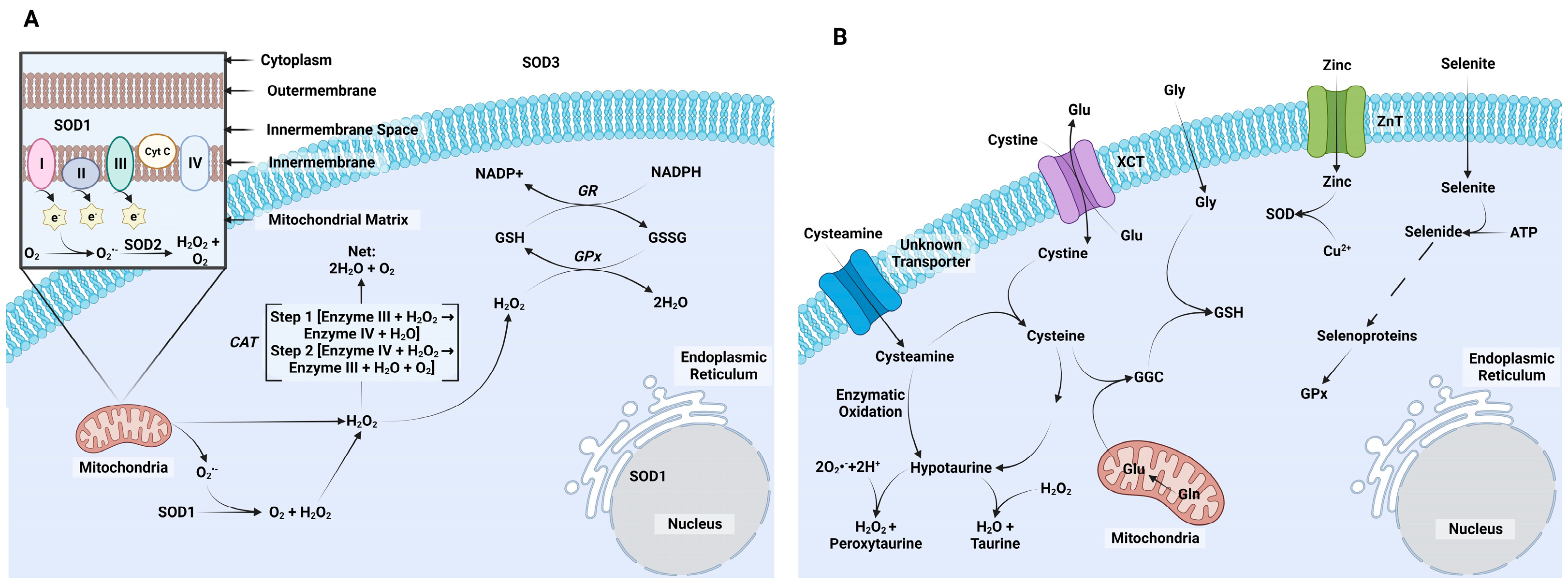
3. Intrinsic ROS Mitigation Systems
4. Antioxidants Examined in Bovine Embryos
4.1. Cysteine
4.2. Cysteamine
4.3. Selenium
4.4. Hypotaurine
4.5. Zinc
5. Antioxidant Potential to Improve Bovine IVP Systems
5.1. IVM
5.2. IVF
5.3. IVC
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nemcova, L.; Rosenbaum Bartkova, A.; Kinterova, V.; Toralova, T. Importance of Supplementation during In Vitro Production of Livestock Animals. In Theriogenology—Recent Advances in the Field; IntechOpen: London, UK, 2023. [Google Scholar]
- Kępka, K.; Wójcik, E.; Wysokińska, A. Assessment of the Genomic Stability of Calves Obtained from Artificial Insemination and OPU/IVP in Vitro Fertilization. Reprod. Domest. Anim. 2023, 58, 1289–1297. [Google Scholar] [CrossRef] [PubMed]
- Tšuiko, O.; Catteeuw, M.; Zamani Esteki, M.; Destouni, A.; Bogado Pascottini, O.; Besenfelder, U.; Havlicek, V.; Smits, K.; Kurg, A.; Salumets, A.; et al. Genome Stability of Bovine in Vivo-Conceived Cleavage-Stage Embryos Is Higher Compared to in Vitro-Produced Embryos. Hum. Reprod. 2017, 32, 2348–2357. [Google Scholar] [CrossRef] [PubMed]
- Hansen, P.J. The Incompletely Fulfilled Promise of Embryo Transfer in Cattle—Why Aren’t Pregnancy Rates Greater and What Can We Do about It? J. Anim. Sci. 2020, 98. [Google Scholar] [CrossRef] [PubMed]
- Ealy, A.D.; Wooldridge, L.K.; McCoski, S.R. BOARD INVITED REVIEW: Post-Transfer Consequences of in Vitro-Produced Embryos in Cattle. J. Anim. Sci. 2019, 97, 2555–2568. [Google Scholar] [CrossRef] [PubMed]
- dos Santos, É.C.; da Fonseca Junior, A.M.; de Lima, C.B.; Ispada, J.; da Silva, J.V.A.; Milazzotto, M.P. Less Is More: Reduced Nutrient Concentration during in Vitro Culture Improves Embryo Production Rates and Morphophysiology of Bovine Embryos. Theriogenology 2021, 173, 37–47. [Google Scholar] [CrossRef]
- Thompson, J.G.E.; Simpson, A.C.; Pugh, P.A.; Donnelly, P.E.; Tervit, H.R. Effect of Oxygen Concentration on In-Vitro Development of Preimplantation Sheep and Cattle Embryos. J. Reprod. Fertil. 1990, 89, 573–578. [Google Scholar] [CrossRef] [PubMed]
- Steeves, T.E.; Gardner, D.K. Temporal and Differential Effects of Amino Acids on Bovine Embryo Development in Culture1. Biol. Reprod. 1999, 61, 731–740. [Google Scholar] [CrossRef] [PubMed]
- Oyamada, T.; Fukui, Y. Oxygen Tension and Medium Supplements for In Vitro Maturation of Bovine Oocytes Cultured Individually in a Chemically Defined Medium. J. Reprod. Dev. 2004, 50, 107–117. [Google Scholar] [CrossRef]
- Krisher, R.L.; Lane, M.; Bavister, B.D. Developmental Competence and Metabolism of Bovine Embryos Cultured in Semi-Defined and Defined Culture Media. Biol. Reprod. 1999, 60, 1345–1352. [Google Scholar] [CrossRef]
- Phaniendra, A.; Jestadi, D.B.; Periyasamy, L. Free Radicals: Properties, Sources, Targets, and Their Implication in Various Diseases. Indian J. Clin. Biochem. 2015, 30, 11–26. [Google Scholar] [CrossRef]
- Nandi, A.; Yan, L.-J.; Jana, C.K.; Das, N. Role of Catalase in Oxidative Stress- and Age-Associated Degenerative Diseases. Oxid. Med. Cell. Longev. 2019, 2019, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B.; Whiteman, M. Measuring Reactive Species and Oxidative Damage in Vivo and in Cell Culture: How Should You Do It and What Do the Results Mean? Br. J. Pharmacol. 2009, 142, 231–255. [Google Scholar] [CrossRef] [PubMed]
- Di Meo, S.; Reed, T.T.; Venditti, P.; Victor, V.M. Role of ROS and RNS Sources in Physiological and Pathological Conditions. Oxid. Med. Cell. Longev. 2016, 2016, 1–44. [Google Scholar] [CrossRef] [PubMed]
- He, L.; He, T.; Farrar, S.; Ji, L.; Liu, T.; Ma, X. Antioxidants Maintain Cellular Redox Homeostasis by Elimination of Reactive Oxygen Species. Cell. Physiol. Biochem. 2017, 44, 532–553. [Google Scholar] [CrossRef] [PubMed]
- Rasouli, M.; Fattahi, R.; Nuoroozi, G.; Zarei-Behjani, Z.; Yaghoobi, M.; Hajmohammadi, Z.; Hosseinzadeh, S. The Role of Oxygen Tension in Cell Fate and Regenerative Medicine: Implications of Hypoxia/Hyperoxia and Free Radicals. Cell Tissue Bank. 2023. [Google Scholar] [CrossRef] [PubMed]
- Henzler, T.; Steudle, E. Transport and Metabolic Degradation of Hydrogen Peroxide in Chara Corallina: Model Calculations and Measurements with the Pressure Probe Suggest Transport of H2O2 across Water Channels. J. Exp. Bot. 2000, 51, 2053–2066. [Google Scholar] [CrossRef] [PubMed]
- Bienert, G.P.; Møller, A.L.B.; Kristiansen, K.A.; Schulz, A.; Møller, I.M.; Schjoerring, J.K.; Jahn, T.P. Specific Aquaporins Facilitate the Diffusion of Hydrogen Peroxide across Membranes. J. Biol. Chem. 2007, 282, 1183–1192. [Google Scholar] [CrossRef]
- Smith, R.A.J.; Hartley, R.C.; Cochemé, H.M.; Murphy, M.P. Mitochondrial Pharmacology. Trends Pharmacol. Sci. 2012, 33, 341–352. [Google Scholar] [CrossRef]
- Liemburg-Apers, D.C.; Willems, P.H.G.M.; Koopman, W.J.H.; Grefte, S. Interactions between Mitochondrial Reactive Oxygen Species and Cellular Glucose Metabolism. Arch. Toxicol. 2015, 89, 1209–1226. [Google Scholar] [CrossRef]
- Fenton, H.J.H. LXXIII.—Oxidation of Tartaric Acid in Presence of Iron. J. Chem. Soc. Trans. 1894, 65, 899–910. [Google Scholar] [CrossRef]
- Haber, F.; Weiss, J. The Catalytic Decomposition of Hydrogen Peroxide by Iron Salts. Proc. R. Soc. London Ser. A-Math. Phys. Sci. 1934, 147, 332–351. [Google Scholar] [CrossRef]
- Cao, S.S.; Kaufman, R.J. Endoplasmic Reticulum Stress and Oxidative Stress in Cell Fate Decision and Human Disease. Antioxid. Redox Signal. 2014, 21, 396–413. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Simmen, T. Mechanistic Connections between Endoplasmic Reticulum (ER) Redox Control and Mitochondrial Metabolism. Cells 2019, 8, 1071. [Google Scholar] [CrossRef] [PubMed]
- Bhandary, B.; Marahatta, A.; Kim, H.-R.; Chae, H.-J. An Involvement of Oxidative Stress in Endoplasmic Reticulum Stress and Its Associated Diseases. Int. J. Mol. Sci. 2012, 14, 434–456. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Zhu, Y.; Basang, W.; Wang, X.; Li, C.; Zhou, X. Roles of Nitric Oxide in the Regulation of Reproduction: A Review. Front. Endocrinol. 2021, 12, 752410. [Google Scholar] [CrossRef] [PubMed]
- Ignarro, L.J.; Buga, G.M.; Wood, K.S.; Byrns, R.E.; Chaudhuri, G. Endothelium-Derived Relaxing Factor Produced and Released from Artery and Vein Is Nitric Oxide. Proc. Natl. Acad. Sci. USA 1987, 84, 9265–9269. [Google Scholar] [CrossRef] [PubMed]
- Pandey, A.N.; Tripathi, A.; PremKumar, K.V.; Shrivastav, T.G.; Chaube, S.K. Reactive Oxygen and Nitrogen Species during Meiotic Resumption from Diplotene Arrest in Mammalian Oocytes. J. Cell. Biochem. 2010, 111, 521–528. [Google Scholar] [CrossRef]
- Radi, R.; Cassina, A.; Hodara, R.; Quijano, C.; Castro, L. Peroxynitrite Reactions and Formation in Mitochondria. Free Radic. Biol. Med. 2002, 33, 1451–1464. [Google Scholar] [CrossRef]
- Sturmey, R.G.; Hawkhead, J.A.; Barker, E.A.; Leese, H.J. DNA Damage and Metabolic Activity in the Preimplantation Embryo. Hum. Reprod. 2008, 24, 81–91. [Google Scholar] [CrossRef]
- Haghdoost, S.; Czene, S.; Näslund, I.; Skog, S.; Harms-Ringdahl, M. Extracellular 8-Oxo-DG as a Sensitive Parameter for Oxidative Stress in Vivo and in Vitro. Free Radic. Res. 2005, 39, 153–162. [Google Scholar] [CrossRef]
- Gros, L.; Saparbaev, M.K.; Laval, J. Enzymology of the Repair of Free Radicals-Induced DNA Damage. Oncogene 2002, 21, 8905–8925. [Google Scholar] [CrossRef] [PubMed]
- Slupphaug, G.; Kavli, B.; Krokan, H. The Interacting Pathways for Prevention and Repair of Oxidative DNA Damage. Mutat. Res. Mol. Mech. Mutagen. 2003, 531, 231–251. [Google Scholar] [CrossRef] [PubMed]
- Ames, B.N.; Shigenaga, M.K.; Hagen, T.M. Oxidants, Antioxidants, and the Degenerative Diseases of Aging. Proc. Natl. Acad. Sci. USA 1993, 90, 7915–7922. [Google Scholar] [CrossRef] [PubMed]
- Malott, K.F.; Reshel, S.; Ortiz, L.; Luderer, U. Glutathione Deficiency Decreases Lipid Droplet Stores and Increases Reactive Oxygen Species in Mouse Oocytes. Biol. Reprod. 2022, 106, 1218–1231. [Google Scholar] [CrossRef] [PubMed]
- Bomfim, M.M.; Andrade, G.M.; del Collado, M.; Sangalli, J.R.; Fontes, P.K.; Nogueira, M.F.G.; Meirelles, F.V.; da Silveira, J.C.; Perecin, F. Antioxidant Responses and Deregulation of Epigenetic Writers and Erasers Link Oxidative Stress and DNA Methylation in Bovine Blastocysts. Mol. Reprod. Dev. 2017, 84, 1296–1305. [Google Scholar] [CrossRef] [PubMed]
- Kietzmann, T.; Petry, A.; Shvetsova, A.; Gerhold, J.M.; Görlach, A. The Epigenetic Landscape Related to Reactive Oxygen Species Formation in the Cardiovascular System. Br. J. Pharmacol. 2017, 174, 1533–1554. [Google Scholar] [CrossRef] [PubMed]
- Zheng, P.; Schramm, R.D.; Latham, K.E. Developmental Regulation and In Vitro Culture Effects on Expression of DNA Repair and Cell Cycle Checkpoint Control Genes in Rhesus Monkey Oocytes and Embryos1. Biol. Reprod. 2005, 72, 1359–1369. [Google Scholar] [CrossRef]
- Pratt, D.A.; Tallman, K.A.; Porter, N.A. Free Radical Oxidation of Polyunsaturated Lipids: New Mechanistic Insights and the Development of Peroxyl Radical Clocks. Acc. Chem. Res. 2011, 44, 458–467. [Google Scholar] [CrossRef]
- Juan, C.A.; Pérez de la Lastra, J.M.; Plou, F.J.; Pérez-Lebeña, E. The Chemistry of Reactive Oxygen Species (ROS) Revisited: Outlining Their Role in Biological Macromolecules (DNA, Lipids and Proteins) and Induced Pathologies. Int. J. Mol. Sci. 2021, 22, 4642. [Google Scholar] [CrossRef]
- Marnett, L.J. Lipid Peroxidation—DNA Damage by Malondialdehyde. Mutat. Res. Mol. Mech. Mutagen. 1999, 424, 83–95. [Google Scholar] [CrossRef]
- Wang, L.; Wang, D.; Zou, X.; Xu, C. Mitochondrial Functions on Oocytes and Preimplantation Embryos. J. Zhejiang Univ. Sci. B 2009, 10, 483–492. [Google Scholar] [CrossRef] [PubMed]
- Dean, R.T.; Fu, S.; Stocker, R.; Davies, M.J. Biochemistry and Pathology of Radical-Mediated Protein Oxidation. Biochem. J. 1997, 324, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Butterfield, D.A.; Koppal, T.; Howard, B.; Subramaniam, R.; Hall, N.; Hensley, K.; Yatin, S.; Allen, K.; Aksenov, M.; Aksenova, M.; et al. Structural and Functional Changes in Proteins Induced by Free Radical-Mediated Oxidative Stress and Protective Action of the Antioxidants N-Tert-Butyl-Alpha-Phenylnitrone and Vitamin Ea. Ann. N. Y. Acad. Sci. 1998, 854, 448–462. [Google Scholar] [CrossRef] [PubMed]
- Gardner, D.K.; Pool, T.B.; Lane, M. Embryo Nutrition and Energy Metabolism and Its Relationship to Embryo Growth, Differentiation, and Viability. Semin. Reprod. Med. 2000, 18, 205–218. [Google Scholar] [CrossRef] [PubMed]
- Leese, H.J. Metabolism of the Preimplantation Embryo: 40 Years On. Reproduction 2012, 143, 417–427. [Google Scholar] [CrossRef] [PubMed]
- Leese, H.J.; Guerif, F.; Allgar, V.; Brison, D.R.; Lundin, K.; Sturmey, R.G. Biological Optimization, the Goldilocks Principle, and How Much Is Lagom in the Preimplantation Embryo. Mol. Reprod. Dev. 2016, 83, 748–754. [Google Scholar] [CrossRef] [PubMed]
- Gardner, D.K.; Lane, M. Culture of Viable Human Blastocysts in Defined Sequential Serum-Free Media. Hum. Reprod. 1998, 13, 148–159. [Google Scholar] [CrossRef]
- de Lima, C.B.; dos Santos, É.C.; Ispada, J.; Fontes, P.K.; Nogueira, M.F.G.; dos Santos, C.M.D.; Milazzotto, M.P. The Dynamics between in Vitro Culture and Metabolism: Embryonic Adaptation to Environmental Changes. Sci. Rep. 2020, 10, 15672. [Google Scholar] [CrossRef]
- Gimeno, I.; García-Manrique, P.; Carrocera, S.; López-Hidalgo, C.; Valledor, L.; Martín-González, D.; Gómez, E. The Metabolic Signature of In Vitro Produced Bovine Embryos Helps Predict Pregnancy and Birth after Embryo Transfer. Metabolites 2021, 11, 484. [Google Scholar] [CrossRef]
- Fischer, B.; Bavister, B.D. Oxygen Tension in the Oviduct and Uterus of Rhesus Monkeys, Hamsters and Rabbits. J. Reprod. Fertil. 1993, 99, 673–679. [Google Scholar] [CrossRef]
- Takahashi, M.; Keicho, K.; Takahashi, H.; Ogawa, H.; Schulte, R.M.; Okano, A. Effect of Oxidative Stress on Development and DNA Damage in In-Vitro Cultured Bovine Embryos by Comet Assay. Theriogenology 2000, 54, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Kitagawa, Y.; Suzuki, K.; Yoneda, A.; Watanabe, T. Effects of Oxygen Concentration and Antioxidants on the in Vitro Developmental Ability, Production of Reactive Oxygen Species (ROS), and DNA Fragmentation in Porcine Embryos. Theriogenology 2004, 62, 1186–1197. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, M.; Nagai, T.; Okamura, N.; Takahashi, H.; Okano, A. Promoting Effect of β-Mercaptoethanol on In Vitro Development under Oxidative Stress and Cystine Uptake of Bovine Embryos1. Biol. Reprod. 2002, 66, 562–567. [Google Scholar] [CrossRef] [PubMed]
- Voelkel, S.A.; Hu, Y.X. Effect of Gas Atmosphere on the Development of One-Cell Bovine Embryos in Two Culture Systems. Theriogenology 1992, 37, 1117–1131. [Google Scholar] [CrossRef] [PubMed]
- Whitty, A.; Kind, K.L.; Dunning, K.R.; Thompson, J.G. Effect of Oxygen and Glucose Availability during in Vitro Maturation of Bovine Oocytes on Development and Gene Expression. J. Assist. Reprod. Genet. 2021, 38, 1349–1362. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, S.; Minami, N.; Takakura, R.; Yamada, M.; Imai, H.; Kashima, N. Low Oxygen Tension during in Vitro Maturation Is Beneficial for Supporting the Subsequent Development of Bovine Cumulus-Oocyte Complexes. Mol. Reprod. Dev. 2000, 57, 353–360. [Google Scholar] [CrossRef] [PubMed]
- Pinyopummintr, T.; Bavister, B.D. Optimum Gas Atmosphere for in Vitro Maturation and in Vitro Fertilization of Bovine Oocytes. Theriogenology 1995, 44, 471–477. [Google Scholar] [CrossRef]
- Báez, F.; de Brun, V.; Rodríguez-Osorio, N.; Viñoles, C. Low Oxygen Tension during in Vitro Oocyte Maturation and Fertilisation Improves Cryotolerance of Bovine Blastocysts Produced in Vitro. Reprod. Fertil. Dev. 2021, 34, 251. [Google Scholar] [CrossRef]
- Takahashi, M.; Saka, N.; Takahashi, H.; Kanai, Y.; Schultz, R.M.; Okano, A. Assessment of DNA Damage in Individual Hamster Embryos by Comet Assay. Mol. Reprod. Dev. 1999, 54, 1–7. [Google Scholar] [CrossRef]
- Kielbassa, C.; Roza, L.; Epe, B. Wavelength Dependence of Oxidative DNA Damage Induced by UV and Visible Light. Carcinogenesis 1997, 18, 811–816. [Google Scholar] [CrossRef]
- Oh, S.J.; Gong, S.P.; Lee, S.T.; Lee, E.J.; Lim, J.M. Light Intensity and Wavelength during Embryo Manipulation Are Important Factors for Maintaining Viability of Preimplantation Embryos in Vitro. Fertil. Steril. 2007, 88, 1150–1157. [Google Scholar] [CrossRef] [PubMed]
- Takenaka, M.; Horiuchi, T.; Yanagimachi, R. Effects of Light on Development of Mammalian Zygotes. Proc. Natl. Acad. Sci. USA 2007, 104, 14289–14293. [Google Scholar] [CrossRef] [PubMed]
- Korhonen, K.; Sjovall, S.; Viitanen, J.; Ketoja, E.; Makarevich, A.; Peippo, J. Viability of Bovine Embryos Following Exposure to the Green Filtered or Wider Bandwidth Light during in Vitro Embryo Production. Hum. Reprod. 2009, 24, 308–314. [Google Scholar] [CrossRef] [PubMed]
- FitzHarris, G.; Baltz, J.M. Granulosa Cells Regulate Intracellular PH of the Murine Growing Oocyte via Gap Junctions: Development of Independent Homeostasis during Oocyte Growth. Development 2006, 133, 591–599. [Google Scholar] [CrossRef] [PubMed]
- Erdogan, S.; FitzHarris, G.; Tartia, A.P.; Baltz, J.M. Mechanisms Regulating Intracellular PH Are Activated during Growth of the Mouse Oocyte Coincident with Acquisition of Meiotic Competence. Dev. Biol. 2005, 286, 352–360. [Google Scholar] [CrossRef] [PubMed]
- Steel, T.; Conaghan, J. PH Equilibration Dynamics of Culture Medium Under Oil. Fertil. Steril. 2008, 89, S27. [Google Scholar] [CrossRef]
- Scarica, C.; Monaco, A.; Borini, A.; Pontemezzo, E.; Bonanni, V.; De Santis, L.; Zacà, C.; Coticchio, G. Use of Mineral Oil in IVF Culture Systems: Physico-Chemical Aspects, Management, and Safety. J. Assist. Reprod. Genet. 2022, 39, 883–892. [Google Scholar] [CrossRef]
- Agarwal, A.; Maldonado Rosas, I.; Anagnostopoulou, C.; Cannarella, R.; Boitrelle, F.; Munoz, L.V.; Finelli, R.; Durairajanayagam, D.; Henkel, R.; Saleh, R. Oxidative Stress and Assisted Reproduction: A Comprehensive Review of Its Pathophysiological Role and Strategies for Optimizing Embryo Culture Environment. Antioxidants 2022, 11, 477. [Google Scholar] [CrossRef] [PubMed]
- Conaghan, J. PH Control in the Embryo Culture Environment. In Culture Media, Solutions, and Systems in Human ART; Cambridge University Press: Cambridge, UK, 2014; pp. 142–154. [Google Scholar]
- Lamber, A.J.; Brand, M.D. Superoxide Production by NADH:Ubiquinone Oxidoreductase (Complex I) Depends on the PH Gradient across the Mitochondrial Inner Membrane. Biochem. J. 2004, 382, 511–517. [Google Scholar] [CrossRef]
- Selivanov, V.A.; Zeak, J.A.; Roca, J.; Cascante, M.; Trucco, M.; Votyakova, T.V. The Role of External and Matrix PH in Mitochondrial Reactive Oxygen Species Generation. J. Biol. Chem. 2008, 283, 29292–29300. [Google Scholar] [CrossRef]
- Shi, D.; Avery, B.; Greve, T. Effects of Temperature Gradients on in Vitro Maturation of Bovine Oocytes. Theriogenology 1998, 50, 667–674. [Google Scholar] [CrossRef] [PubMed]
- Swain, J.E.; Pool, T.B. New PH-Buffering System for Media Utilized during Gamete and Embryo Manipulations for Assisted Reproduction. Reprod. Biomed. Online 2009, 18, 799–810. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.-F.; Wang, W.-H.; Keefe, D.L. Overheating Is Detrimental to Meiotic Spindles within in Vitro Matured Human Oocytes. Zygote 2004, 12, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.-H.; Meng, L.; Hackett, R.J.; Odenbourg, R.; Keefe, D.L. Limited Recovery of Meiotic Spindles in Living Human Oocytes after Cooling–Rewarming Observed Using Polarized Light Microscopy. Hum. Reprod. 2001, 16, 2374–2378. [Google Scholar] [CrossRef] [PubMed]
- Leese, H.J.; Baumann, C.G.; Brison, D.R.; McEvoy, T.G.; Sturmey, R.G. Metabolism of the Viable Mammalian Embryo: Quietness Revisited. Mol. Hum. Reprod. 2008, 14, 667–672. [Google Scholar] [CrossRef] [PubMed]
- Sen, U. Maturation of Bovine Oocytes under Low Culture Temperature Decreased Glutathione Peroxidase Activity of Both Oocytes and Blastocysts. Pol. J. Vet. Sci. 2021, 24, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Nabenishi, H.; Ohta, H.; Nishimoto, T.; Morita, T.; Ashizawa, K.; Tsuzuki, Y. The Effects of Cysteine Addition during in Vitro Maturation on the Developmental Competence, ROS, GSH and Apoptosis Level of Bovine Oocytes Exposed to Heat Stress. Zygote 2012, 20, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Len, J.S.; Koh, W.S.D.; Tan, S.-X. The Roles of Reactive Oxygen Species and Antioxidants in Cryopreservation. Biosci. Rep. 2019, 39, BSR20191601. [Google Scholar] [CrossRef]
- Estudillo, E.; Jiménez, A.; Bustamante-Nieves, P.E.; Palacios-Reyes, C.; Velasco, I.; López-Ornelas, A. Cryopreservation of Gametes and Embryos and Their Molecular Changes. Int. J. Mol. Sci. 2021, 22, 10864. [Google Scholar] [CrossRef]
- Bilodeau, J.-F.; Chatterjee, S.; Sirard, M.-A.; Gagnon, C. Levels of Antioxidant Defenses Are Decreased in Bovine Spermatozoa after a Cycle of Freezing and Thawing. Mol. Reprod. Dev. 2000, 55, 282–288. [Google Scholar] [CrossRef]
- Sies, H.; Jones, D.P. Reactive Oxygen Species (ROS) as Pleiotropic Physiological Signalling Agents. Nat. Rev. Mol. Cell Biol. 2020, 21, 363–383. [Google Scholar] [CrossRef] [PubMed]
- Eleutherio, E.C.A.; Silva Magalhães, R.S.; de Araújo Brasil, A.; Monteiro Neto, J.R.; de Holanda Paranhos, L. SOD1, More than Just an Antioxidant. Arch. Biochem. Biophys. 2021, 697, 108701. [Google Scholar] [CrossRef] [PubMed]
- Ganini, D.; Santos, J.H.; Bonini, M.G.; Mason, R.P. Switch of Mitochondrial Superoxide Dismutase into a Prooxidant Peroxidase in Manganese-Deficient Cells and Mice. Cell Chem. Biol. 2019, 25, 413–425.e6. [Google Scholar] [CrossRef] [PubMed]
- Franklin, C.C.; Backos, D.S.; Mohar, I.; White, C.C.; Forman, H.J.; Kavanagh, T.J. Structure, Function, and Post-Translational Regulation of the Catalytic and Modifier Subunits of Glutamate Cysteine Ligase. Mol. Aspects Med. 2009, 30, 86–98. [Google Scholar] [CrossRef] [PubMed]
- de Matos, D.G.; Herrera, C.; Cortvrindt, R.; Smitz, J.; Van Soom, A.; Nogueira, D.; Pasqualini, R.S. Cysteamine Supplementation during in Vitro Maturation and Embryo Culture: A Useful Tool for Increasing the Efficiency of Bovine in Vitro Embryo Production. Mol. Reprod. Dev. 2002, 62, 203–209. [Google Scholar] [CrossRef] [PubMed]
- Couto, N.; Wood, J.; Barber, J. The Role of Glutathione Reductase and Related Enzymes on Cellular Redox Homoeostasis Network. Free Radic. Biol. Med. 2016, 95, 27–42. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.M.; Liou, S.S.; Hansel, W. Intracytoplasmic Glutathione Concentration and the Role of β-Mercaptoethanol in Preimplantation Development of Bovine Embryos. Theriogenology 1996, 46, 429–439. [Google Scholar] [CrossRef]
- Merton, J.S.; Knijn, H.M.; Flapper, H.; Dotinga, F.; Roelen, B.A.J.; Vos, P.L.A.M.; Mullaart, E. Cysteamine Supplementation during in Vitro Maturation of Slaughterhouse- and Opu-Derived Bovine Oocytes Improves Embryonic Development without Affecting Cryotolerance, Pregnancy Rate, and Calf Characteristics. Theriogenology 2013, 80, 365–371. [Google Scholar] [CrossRef]
- Sutovsky, P.; Schatten, G. Depletion of Glutathione during Bovine Oocyte Maturation Reversibly Blocks the Decondensation of the Male Pronucleus and Pronuclear Apposition during Fertilization1. Biol. Reprod. 1997, 56, 1503–1512. [Google Scholar] [CrossRef]
- de Matos, D.G.; Furnus, C.C. The Importance of Having High Glutathione (GSH) Level after Bovine in Vitro Maturation on Embryo Development: Effect of β-Mercaptoethanol, Cysteine and Cystine. Theriogenology 2000, 53, 761–771. [Google Scholar] [CrossRef]
- Zuelke, K.A.; Jeffay, S.C.; Zucker, R.M.; Perreault, S.D. Glutathione (GSH) Concentrations Vary with the Cell Cycle in Maturing Hamster Oocytes, Zygotes, and Pre-Implantation Stage Embryos. Mol. Reprod. Dev. 2003, 64, 106–112. [Google Scholar] [CrossRef] [PubMed]
- García-Martínez, T.; Vendrell-Flotats, M.; Martínez-Rodero, I.; Ordóñez-León, E.A.; Álvarez-Rodríguez, M.; López-Béjar, M.; Yeste, M.; Mogas, T. Glutathione Ethyl Ester Protects In Vitro-Maturing Bovine Oocytes against Oxidative Stress Induced by Subsequent Vitrification/Warming. Int. J. Mol. Sci. 2020, 21, 7547. [Google Scholar] [CrossRef] [PubMed]
- Shi, F.; Li, H.; Wang, E.; Chen, Z.; Zhang, C. Melatonin Reduces Two-cell Block via Nonreceptor Pathway in Mice. J. Cell. Biochem. 2018, 119, 9380–9393. [Google Scholar] [CrossRef] [PubMed]
- Gardiner, C.S.; Reed, D.J. Synthesis of Glutathione in the Preimplantation Mouse Embryo. Arch. Biochem. Biophys. 1995, 318, 30–36. [Google Scholar] [CrossRef] [PubMed]
- de Matos, D.G.; Furnus, C.C.; Moses, D.F. Glutathione Synthesis During in Vitro Maturation of Bovine Oocytes: Role of Cumulus Cells1. Biol. Reprod. 1997, 57, 1420–1425. [Google Scholar] [CrossRef] [PubMed]
- Luberda, Z. The Role of Glutathione in Mammalian Gametes. Reprod. Biol. 2005, 5, 5–17. [Google Scholar]
- Negrón-Pérez, V.M.; Fausnacht, D.W.; Rhoads, M.L. Invited Review: Management Strategies Capable of Improving the Reproductive Performance of Heat-Stressed Dairy Cattle. J. Dairy Sci. 2019, 102, 10695–10710. [Google Scholar] [CrossRef] [PubMed]
- Tutt, D.A.R.; Guven-Ates, G.; Kwong, W.Y.; Simmons, R.; Sang, F.; Silvestri, G.; Canedo-Ribeiro, C.; Handyside, A.H.; Labrecque, R.; Sirard, M.-A.; et al. Developmental, Cytogenetic and Epigenetic Consequences of Removing Complex Proteins and Adding Melatonin during in Vitro Maturation of Bovine Oocytes. Front. Endocrinol. 2023, 14. [Google Scholar] [CrossRef]
- Pasquariello, R.; Verdile, N.; Brevini, T.A.L.; Gandolfi, F.; Boiti, C.; Zerani, M.; Maranesi, M. The Role of Resveratrol in Mammalian Reproduction. Molecules 2020, 25, 4554. [Google Scholar] [CrossRef]
- Rodríguez-Varela, C.; Labarta, E. Clinical Application of Antioxidants to Improve Human Oocyte Mitochondrial Function: A Review. Antioxidants 2020, 9, 1197. [Google Scholar] [CrossRef]
- Van Soom, A.; Yuan, Y.; Peelman, L.; de Matos, D.; Dewulf, J.; Laevens, H.; de Kruif, A. Prevalence of Apoptosis and Inner Cell Allocation in Bovine Embryos Cultured under Different Oxygen Tensions with or without Cysteine Addition. Theriogenology 2002, 57, 1453–1465. [Google Scholar] [CrossRef] [PubMed]
- Caamaño, J.N.; Ryoo, Z.Y.; Thomas, J.A.; Youngs, C.R. β-Mercaptoethanol Enhances Blastocyst Formation Rate of Bovine in Vitro-Matured/in Vitro-Fertilized Embryos1. Biol. Reprod. 1996, 55, 1179–1184. [Google Scholar] [CrossRef] [PubMed]
- Barrozo, L.G.; Paulino, L.R.F.M.; Silva, B.R.; Barbalho, E.C.; Nascimento, D.R.; Neto, M.F.L.; Silva, J.R.V. N-Acetyl-Cysteine and the Control of Oxidative Stress during in Vitro Ovarian Follicle Growth, Oocyte Maturation, Embryo Development and Cryopreservation. Anim. Reprod. Sci. 2021, 231, 106801. [Google Scholar] [CrossRef] [PubMed]
- Ezeriņa, D.; Takano, Y.; Hanaoka, K.; Urano, Y.; Dick, T.P. N-Acetyl Cysteine Functions as a Fast-Acting Antioxidant by Triggering Intracellular H2S and Sulfane Sulfur Production. Cell Chem. Biol. 2018, 25, 447–459.e4. [Google Scholar] [CrossRef] [PubMed]
- Meier, T.; Issels, R.D. Promotion of Cyst(e)Ine Uptake. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1995; pp. 103–112. [Google Scholar]
- Deleuze, S.; Goudet, G. Cysteamine Supplementation of In Vitro Maturation Media: A Review. Reprod. Domest. Anim. 2010, 45, e476–e482. [Google Scholar] [CrossRef] [PubMed]
- Ho, E. Zinc Deficiency, DNA Damage and Cancer Risk. J. Nutr. Biochem. 2004, 15, 572–578. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Cohen, H.J. Selenium-Dependent Glutathione Peroxidase Protein and Activity: Immunological Investigations on Cellular and Plasma Enzymes. Blood 1986, 68, 640–645. [Google Scholar] [CrossRef]
- Basini, G.; Tamanini, C. Selenium Stimulates Estradiol Production in Bovine Granulosa Cells: Possible Involvement of Nitric Oxide. Domest. Anim. Endocrinol. 2000, 18, 1–17. [Google Scholar] [CrossRef]
- Mistry, H.D.; Broughton Pipkin, F.; Redman, C.W.G.; Poston, L. Selenium in Reproductive Health. Am. J. Obstet. Gynecol. 2012, 206, 21–30. [Google Scholar] [CrossRef]
- Mihailovič, M.; Cvetkovč, M.; Ljubič, A.; Kosanovič, M.; Nedeljkovič, S.; Jovanovič, I.; Pešut, O. Selenium and Malondialdehyde Content and Glutathione Peroxidase Activity in Maternal and Umbilical Cord Blood and Amniotic Fluid. Biol. Trace Elem. Res. 2000, 73, 47–54. [Google Scholar] [CrossRef]
- Cerny, K.L.; Anderson, L.; Burris, W.R.; Rhoads, M.; Matthews, J.C.; Bridges, P.J. Form of Supplemental Selenium Fed to Cycling Cows Affects Systemic Concentrations of Progesterone but Not Those of Estradiol. Theriogenology 2016, 85, 800–806. [Google Scholar] [CrossRef] [PubMed]
- da Silva, J.S.; Rosa, A.F.; Moncau, C.T.; Silva-Vignato, B.; Pugine, S.M.P.; de Melo, M.P.; Sanchez, J.M.D.; Zanetti, M.A. Effect of Different Selenium Sources and Concentrations on Glutathione Peroxidase Activity and Cholesterol Metabolism of Beef Cattle. J. Anim. Sci. 2021, 99. [Google Scholar] [CrossRef] [PubMed]
- Pieczyńska, J.; Grajeta, H. The Role of Selenium in Human Conception and Pregnancy. J. Trace Elem. Med. Biol. 2015, 29, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.T. Metabolomics of Selenium: Se Metabolites Based on Speciation Studies. J. Heal. Sci. 2005, 51, 107–114. [Google Scholar] [CrossRef]
- Wang, Z.; Kong, L.; Zhu, L.; Hu, X.; Su, P.; Song, Z. The Mixed Application of Organic and Inorganic Selenium Shows Better Effects on Incubation and Progeny Parameters. Poult. Sci. 2021, 100, 1132–1141. [Google Scholar] [CrossRef] [PubMed]
- Grove, R.Q.; Karpowicz, S.J. Reaction of Hypotaurine or Taurine with Superoxide Produces the Organic Peroxysulfonic Acid Peroxytaurine. Free Radic. Biol. Med. 2017, 108, 575–584. [Google Scholar] [CrossRef] [PubMed]
- Aruoma, O.I.; Halliwell, B.; Hoey, B.M.; Butler, J. The Antioxidant Action of Taurine, Hypotaurine and Their Metabolic Precursors. Biochem. J. 1988, 256, 251–255. [Google Scholar] [CrossRef] [PubMed]
- Guérin, P.; El Mouatassim, S.; Ménézo, Y. Oxidative Stress and Protection against Reactive Oxygen Species in the Pre-Implantation Embryo and Its Surroundings. Hum. Reprod. Update 2001, 7, 175–189. [Google Scholar] [CrossRef]
- Guérin, P.; Guillaud, J.; Ménézo, Y. Andrology: Hypotaurine in Spermatozoa and Genital Secretions and Its Production by Oviduct Epithelial Cells in Vitro. Hum. Reprod. 1995, 10, 866–872. [Google Scholar] [CrossRef]
- McCall, K.A.; Huang, C.; Fierke, C.A. Function and Mechanism of Zinc Metalloenzymes. J. Nutr. 2000, 130, 1437S–1446S. [Google Scholar] [CrossRef]
- Powell, S.R. The Antioxidant Properties of Zinc. J. Nutr. 2000, 130, 1447S–1454S. [Google Scholar] [CrossRef] [PubMed]
- Duffy, R.; Yin, M.; Redding, L.E. A Review of the Impact of Dietary Zinc on Livestock Health. J. Trace Elem. Miner. 2023, 5, 100085. [Google Scholar] [CrossRef]
- Kumar, N.; Verma, R.P.; Singh, L.P.; Varshney, V.P.; Dass, R.S. Effect of Different Levels and Sources of Zinc Supplementation on Quantitative and Qualitative Semen Attributes and Serum Testosterone Level in Crossbred Cattle (Bos Indicus × Bos Taurus) Bulls. Reprod. Nutr. Dev. 2006, 46, 663–675. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Diaz, F.J. Zinc Depletion Causes Multiple Defects in Ovarian Function during the Periovulatory Period in Mice. Endocrinology 2012, 153, 873–886. [Google Scholar] [CrossRef] [PubMed]
- Galarza, E.M.; Lizarraga, R.M.; Anchordoquy, J.P.; Farnetano, N.A.; Furnus, C.C.; Fazzio, L.E.; Anchordoquy, J.M. Zinc Supplementation within the Reference Ranges for Zinc Status in Cattle Improves Sperm Quality without Modifying in Vitro Fertilization Performance. Anim. Reprod. Sci. 2020, 221, 106595. [Google Scholar] [CrossRef] [PubMed]
- Pham, A.N.; Xing, G.; Miller, C.J.; Waite, T.D. Fenton-like Copper Redox Chemistry Revisited: Hydrogen Peroxide and Superoxide Mediation of Copper-Catalyzed Oxidant Production. J. Catal. 2013, 301, 54–64. [Google Scholar] [CrossRef]
- Garner, M.A.; Ricart, K.C.; Roberts, B.R.; Bomben, V.C.; Basso, M.; Ye, Y.; Sahawneh, J.; Franco, M.C.; Beckman, J.S.; Estévez, A.G. Cu,Zn-Superoxide Dismutase Increases Toxicity of Mutant and Zinc-Deficient Superoxide Dismutase by Enhancing Protein Stability*. J. Biol. Chem. 2010, 285, 33885–33897. [Google Scholar] [CrossRef]
- Iakovidis, I.; Delimaris, I.; Piperakis, S.M. Copper and Its Complexes in Medicine: A Biochemical Approach. Mol. Biol. Int. 2011, 2011, 1–13. [Google Scholar] [CrossRef]
- Tarai, S.K.; Mandal, S.; Tarai, A.; Som, I.; Pan, A.; Bagchi, A.; Biswas, A.; Moi, S.C. Biophysical Study on DNA and BSA Binding Activity of Cu(II) Complex: Synthesis, Molecular Docking, Cytotoxic Activity, and Theoretical Approach. Appl. Organomet. Chem. 2023, 37. [Google Scholar] [CrossRef]
- Mizushima, S.; Fukui, Y. Fertilizability and Developmental Capacity of Bovine Oocytes Cultured Individually in a Chemically Defined Maturation Medium. Theriogenology 2001, 55, 1431–1445. [Google Scholar] [CrossRef]
- Elgebaly, M.M.; Hazaa, A.B.M.; Amer, H.A.; Mesalam, A. L-Cysteine Improves Bovine Oocyte Developmental Competence in Vitro via Activation of Oocyte-derived Growth Factors BMP-15 and GDF-9. Reprod. Domest. Anim. 2022, 57, 734–742. [Google Scholar] [CrossRef] [PubMed]
- Ambrogi, M.; Dall’Acqua, P.; Rocha-Frigoni, N.; Leão, B.; Mingoti, G. Transporting Bovine Oocytes in a Medium Supplemented with Different Macromolecules and Antioxidants: Effects on Nuclear and Cytoplasmic Maturation and Embryonic Development in Vitro. Reprod. Domest. Anim. 2017, 52, 409–421. [Google Scholar] [CrossRef] [PubMed]
- Lott, W.; Anchamparuthy, V.; McGilliard, M.; Mullarky, I.; Gwazdauskas, F. Influence of Cysteine in Conjunction with Growth Factors on the Development of In Vitro-Produced Bovine Embryos. Reprod. Domest. Anim. 2011, 46, 585–594. [Google Scholar] [CrossRef] [PubMed]
- Furnus, C.C.; de Matos, D.G.; Picco, S.; García, P.P.; Inda, A.M.; Mattioli, G.; Errecalde, A.L. Metabolic Requirements Associated with GSH Synthesis during in Vitro Maturation of Cattle Oocytes. Anim. Reprod. Sci. 2008, 109, 88–99. [Google Scholar] [CrossRef] [PubMed]
- Rocha-Frigoni, N.A.S.; Leão, B.C.S.; Dall’Acqua, P.C.; Mingoti, G.Z. Improving the Cytoplasmic Maturation of Bovine Oocytes Matured in Vitro with Intracellular and/or Extracellular Antioxidants Is Not Associated with Increased Rates of Embryo Development. Theriogenology 2016, 86, 1897–1905. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.A.; Bilodeau, J.F.; Sirard, M.A. Antioxidant Requirements for Bovine Oocytes Varies during in Vitro Maturation, Fertilization and Development. Theriogenology 2003, 59, 939–949. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.-S.; Jang, H.; Park, M.-R.; Oh, K.B.; Lee, H.; Hwang, S.; Xu, L.-J.; Hwang, I.-S.; Lee, J.-W. N-Acetyl-L-Cysteine Improves the Developmental Competence of Bovine Oocytes and Embryos Cultured In Vitro by Attenuating Oxidative Damage and Apoptosis. Antioxidants 2021, 10, 860. [Google Scholar] [CrossRef]
- Sovernigo, T.; Adona, P.; Monzani, P.; Guemra, S.; Barros, F.; Lopes, F.; Leal, C. Effects of Supplementation of Medium with Different Antioxidants during in Vitro Maturation of Bovine Oocytes on Subsequent Embryo Production. Reprod. Domest. Anim. 2017, 52, 561–569. [Google Scholar] [CrossRef]
- Lodde, V.; Luciano, A.M.; Musmeci, G.; Miclea, I.; Tessaro, I.; Aru, M.; Albertini, D.F.; Franciosi, F. A Nuclear and Cytoplasmic Characterization of Bovine Oocytes Reveals That Cysteamine Partially Rescues the Embryo Development in a Model of Low Ovarian Reserve. Animals 2021, 11, 1936. [Google Scholar] [CrossRef]
- Zhenwei, J.; Xianhua, Z. Pre-IVM Treatment with C-Type Natriuretic Peptide in the Presence of Cysteamine Enhances Bovine Oocytes Antioxidant Defense Ability and Developmental Competence in Vitro. Iran. J. Vet. Res. 2019, 20, 173–179. [Google Scholar]
- Canel, N.G.; Suvá, M.; Bevacqua, R.J.; Arias, M.E.; Felmer, R.; Salamone, D.F. Improved Embryo Development Using High Cysteamine Concentration during IVM and Sperm Co-Culture with COCs Previous to ICSI in Bovine. Theriogenology 2018, 117, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Balasubramanian, S.; Rho, G.-J. Effect of Cysteamine Supplementation of in Vitro Matured Bovine Oocytes on Chilling Sensitivity and Development of Embryos. Anim. Reprod. Sci. 2007, 98, 282–292. [Google Scholar] [CrossRef] [PubMed]
- Anchordoquy, J.M.P.; Balbi, M.; Farnetano, N.A.; Fabra, M.C.; Carranza-Martin, A.C.; Nikoloff, N.; Giovambattista, G.; Furnus, C.C.; Anchordoquy, J.M.P. Trace Mineral Mixture Supplemented to in Vitro Maturation Medium Improves Subsequent Embryo Development and Embryo Quality in Cattle. Vet. Res. Commun. 2022, 46, 1111–1119. [Google Scholar] [CrossRef] [PubMed]
- Lizarraga, R.M.; Anchordoquy, J.M.P.; Galarza, E.M.; Farnetano, N.A.; Carranza-Martin, A.; Furnus, C.C.; Mattioli, G.A.; Anchordoquy, J.M.P. Sodium Selenite Improves In Vitro Maturation of Bos Primigenius Taurus Oocytes. Biol. Trace Elem. Res. 2020, 197, 149–158. [Google Scholar] [CrossRef] [PubMed]
- Toosinia, S.; Davoodian, N.; Arabi, M.; Kadivar, A. Ameliorating Effect of Sodium Selenite on Developmental and Molecular Response of Bovine Cumulus-Oocyte Complexes Matured in Vitro Under Heat Stress Condition. Biol. Trace Elem. Res. 2024, 202, 161–174. [Google Scholar] [CrossRef] [PubMed]
- Picco, S.J.; Anchordoquy, J.M.; de Matos, D.G.; Anchordoquy, J.P.; Seoane, A.; Mattioli, G.A.; Errecalde, A.L.; Furnus, C.C. Effect of Increasing Zinc Sulphate Concentration during in Vitro Maturation of Bovine Oocytes. Theriogenology 2010, 74, 1141–1148. [Google Scholar] [CrossRef] [PubMed]
- Anchordoquy, J.; Anchordoquy, J.; Sirini, M.; Picco, S.; Peral-García, P.; Furnus, C. The Importance of Having Zinc During In Vitro Maturation of Cattle Cumulus-Oocyte Complex: Role of Cumulus Cells. Reprod. Domest. Anim. 2014, 49, 865–874. [Google Scholar] [CrossRef]
- Anchordoquy, J.M.; Picco, S.J.; Seoane, A.; Anchordoquy, J.P.; Ponzinibbio, M.V.; Mattioli, G.A.; Peral García, P.; Furnus, C.C. Analysis of Apoptosis and DNA Damage in Bovine Cumulus Cells after Exposure in Vitro to Different Zinc Concentrations. Cell Biol. Int. 2011, 35, 593–597. [Google Scholar] [CrossRef]
- Kandil, O.; Alhallag, K.; Shawky, S.; Abu-Alya, I.; Kandeel, S.; Ismali, E.; Fathalla, S. Effect of Zinc and Nano Zinc on Developmental Competence of Buffalo Oocytes. J. Adv. Vet. Res. 2023, 13, 474–478. [Google Scholar]
- Anchordoquy, J.P.M.P.; Anchordoquy, J.P.M.P.; Lizarraga, R.M.; Nikoloff, N.; Pascua, A.M.; Furnus, C.C. The Importance of Trace Minerals Copper, Manganese, Selenium and Zinc in Bovine Sperm–Zona Pellucida Binding. Zygote 2019, 27, 89–96. [Google Scholar] [CrossRef]
- Pons-Rejraji, H.; Vorilhon, S.; Difrane, A.; Dollet, S.; Bourgne, C.; Berger, M.; Chaput, L.; Pereira, B.; Bouche, C.; Drevet, J.R.; et al. Beneficial Effects of Hypotaurine Supplementation in Preparation and Freezing Media on Human Sperm Cryo-Capacitation and DNA Quality. Basic Clin. Androl. 2021, 31, 26. [Google Scholar] [CrossRef] [PubMed]
- Pavlok, A. D-Penicillamine and Granulosa Cells Can Effectively Extend the Fertile Life Span of Bovine Frozen-Thawed Spermatozoa in Vitro: Effect on Fertilization and Polyspermy. Theriogenology 2000, 53, 1135–1146. [Google Scholar] [CrossRef] [PubMed]
- Schuh, S.M.; Hille, B.; Babcock, D.F. Adenosine and Catecholamine Agonists Speed the Flagellar Beat of Mammalian Sperm by a Non-Receptor-Mediated Mechanism1. Biol. Reprod. 2007, 77, 960–969. [Google Scholar] [CrossRef] [PubMed]
- Miller, G.F.; Gliedt, D.W.; Rakes, J.M.; Rorie, R.W. Addition of Penicillamine, Hypotaurine and Epinephrine (PHE) or Bovine Oviductal Epithelial Cells (BOEC) Alone or in Combination to Bovine in Vitro Fertilization Medium Increases the Subsequent Embryo Cleavage Rate. Theriogenology 1994, 41, 689–696. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, F.S.; Barretto, L.S.S.; Arruda, R.P.; Perri, S.H.V.; Mingoti, G.Z. Heparin and Penicillamine–Hypotaurine–Epinephrine (PHE) Solution during Bovine in Vitro Fertilization Procedures Impair the Quality of Spermatozoa but Improve Normal Oocyte Fecundation and Early Embryonic Development. Vitr. Cell. Dev. Biol.-Anim. 2014, 50, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.-S.; Koyama, K.; Huang, W.; Yang, Y.; Yanagawa, Y.; Takahashi, Y.; Nagano, M. Addition of D-Penicillamine, Hypotaurine, and Epinephrine (PHE) Mixture to IVF Medium Maintains Motility and Longevity of Bovine Sperm and Enhances Stable Production of Blastocysts in Vitro. J. Reprod. Dev. 2015, 61, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Silva, N.C.; Leao, K.M.; Padua, J.T.; Marques, T.C.; Neto, F.A.; Dode, M.A.N.; Cunha, A.M. Effect of Different Cryopreservation Extenders Added with Antioxidants on Semen Quality and in Vitro Embryo Production Efficiency in Cattle. An. Acad. Bras. Cienc. 2021, 93. [Google Scholar] [CrossRef]
- Jahanbin, R.; Yazdanshenas, P.; Rahimi, M.; Hajarizadeh, A.; Tvrda, E.; Nazari, S.A.; Mohammadi-Sangcheshmeh, A.; Ghanem, N. In Vivo and In Vitro Evaluation of Bull Semen Processed with Zinc (Zn) Nanoparticles. Biol. Trace Elem. Res. 2021, 199, 126–135. [Google Scholar] [CrossRef]
- Stephenson, J.L.; Brackett, B.G. Influences of Zinc on Fertilisation and Development of Bovine Oocytes in Vitro. Zygote 1999, 7, 195–201. [Google Scholar] [CrossRef]
- Duncan, F.E.; Que, E.L.; Zhang, N.; Feinberg, E.C.; O’Halloran, T.V.; Woodruff, T.K. The Zinc Spark Is an Inorganic Signature of Human Egg Activation. Sci. Rep. 2016, 6, 24737. [Google Scholar] [CrossRef]
- Kerns, K.; Zigo, M.; Drobnis, E.Z.; Sutovsky, M.; Sutovsky, P. Zinc Ion Flux during Mammalian Sperm Capacitation. Nat. Commun. 2018, 9, 2061. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, F.; Barretto, L.; Arruda, R.; Perri, S.; Mingoti, G. Effect of Antioxidants During Bovine In Vitro Fertilization Procedures on Spermatozoa and Embryo Development. Reprod. Domest. Anim. 2010, 45, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Hansen, P.J.; Drost, M.; Rivera, R.M.; Paula-Lopes, F.F.; Al-Katanani, Y.M.; Krininger, C.E.; Chase, C.C. Adverse Impact of Heat Stress on Embryo Production: Causes and Strategies for Mitigation. Theriogenology 2001, 55, 91–103. [Google Scholar] [CrossRef] [PubMed]
- Wydooghe, E.; Heras, S.; Dewulf, J.; Piepers, S.; Van den Abbeel, E.; De Sutter, P.; Vandaele, L.; Van Soom, A. Replacing Serum in Culture Medium with Albumin and Insulin, Transferrin and Selenium Is the Key to Successful Bovine Embryo Development in Individual Culture. Reprod. Fertil. Dev. 2014, 26, 717. [Google Scholar] [CrossRef] [PubMed]
- Guimarães, A.L.S.; Pereira, S.A.; Diógenes, M.N.; Dode, M.A.N. Effect of Insulin–Transferrin–Selenium (ITS) and l-Ascorbic Acid (AA) during in Vitro Maturation on in Vitro Bovine Embryo Development. Zygote 2016, 24, 890–899. [Google Scholar] [CrossRef] [PubMed]
- dos Santos Mendonça-Soares, A.; Guimarães, A.L.S.; Fidelis, A.A.G.; Franco, M.M.; Dode, M.A.N. The Use of Insulin-Transferrin-Selenium (ITS), and Folic Acid on Individual in Vitro Embryo Culture Systems in Cattle. Theriogenology 2022, 184, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Guimarães, A.L.S.; Pereira, S.A.; Kussano, N.R.; Dode, M.A.N. The Effect of Pre-Maturation Culture Using Phosphodiesterase Type 3 Inhibitor and Insulin, Transferrin and Selenium on Nuclear and Cytoplasmic Maturation of Bovine Oocytes. Zygote 2016, 24, 219–229. [Google Scholar] [CrossRef]
- Smits, A.; Leroy, J.L.M.R.; Bols, P.E.J.; De Bie, J.; Marei, W.F.A. Rescue Potential of Supportive Embryo Culture Conditions on Bovine Embryos Derived from Metabolically Compromised Oocytes. Int. J. Mol. Sci. 2020, 21, 8206. [Google Scholar] [CrossRef]
- Raghu, H.M.; Reddy, S.M.; Nandi, S. Effect of Insulin, Transferrin and Selenium and Epidermal Growth Factor on Development of Buffalo Oocytes to the Blastocyst Stage in Vitro in Serum-free, Semidefined Media. Vet. Rec. 2002, 151, 260–265. [Google Scholar] [CrossRef]
- Córdova, B.; Morató, R.; Izquierdo, D.; Paramio, T.; Mogas, T. Effect of the Addition of Insulin-Transferrin-Selenium and/or L-Ascorbic Acid to the in Vitro Maturation of Prepubertal Bovine Oocytes on Cytoplasmic Maturation and Embryo Development. Theriogenology 2010, 74, 1341–1348. [Google Scholar] [CrossRef]
- Wooldridge, L.K.; Nardi, M.E.; Ealy, A.D. Zinc Supplementation during in Vitro Embryo Culture Increases Inner Cell Mass and Total Cell Numbers in Bovine Blastocysts1. J. Anim. Sci. 2019, 97, 4946–4950. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.-J.; Pang, Y.-W.; Liu, Y.; Hao, H.-S.; Zhao, X.-M.; Qin, T.; Zhu, H.-B.; Du, W.-H. Exogenous Glutathione Supplementation in Culture Medium Improves the Bovine Embryo Development after in Vitro Fertilization. Theriogenology 2015, 84, 716–723. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Cui, L.; Yu, D.; Hao, H.; Liu, Y.; Zhao, X.; Pang, Y.; Zhu, H.; Du, W. Exogenous Glutathione Improves Intracellular Glutathione Synthesis via the Γ-glutamyl Cycle in Bovine Zygotes and Cleavage Embryos. J. Cell. Physiol. 2019, 234, 7384–7394. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Keane, J.A.; Ealy, A.D. An Overview of Reactive Oxygen Species Damage Occurring during In Vitro Bovine Oocyte and Embryo Development and the Efficacy of Antioxidant Use to Limit These Adverse Effects. Animals 2024, 14, 330. https://doi.org/10.3390/ani14020330
Keane JA, Ealy AD. An Overview of Reactive Oxygen Species Damage Occurring during In Vitro Bovine Oocyte and Embryo Development and the Efficacy of Antioxidant Use to Limit These Adverse Effects. Animals. 2024; 14(2):330. https://doi.org/10.3390/ani14020330
Chicago/Turabian StyleKeane, Jessica A., and Alan D. Ealy. 2024. "An Overview of Reactive Oxygen Species Damage Occurring during In Vitro Bovine Oocyte and Embryo Development and the Efficacy of Antioxidant Use to Limit These Adverse Effects" Animals 14, no. 2: 330. https://doi.org/10.3390/ani14020330