
Mitochonic Acid 5 Increases Ram Sperm Quality by Improving Mitochondrial Function during Storage at 4 °C
 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Ethical Approval
2.3. Ram Semen Collection and Processing
2.4. Evaluation of Sperm Motility Using a Computer-Assisted Sperm Analysis (CASA) System
2.5. Evaluation of Sperm Membrane Integrity and Acrosome Integrity
2.6. Mitochondrial Activity
2.7. Sperm Mitochondrial ROS Level
2.8. Sperm ATP Contents
2.9. Western Blotting
2.10. Statistical Analysis
3. Results
3.1. Effect of Mitochonic Acid 5 on Sperm Motility Parameters
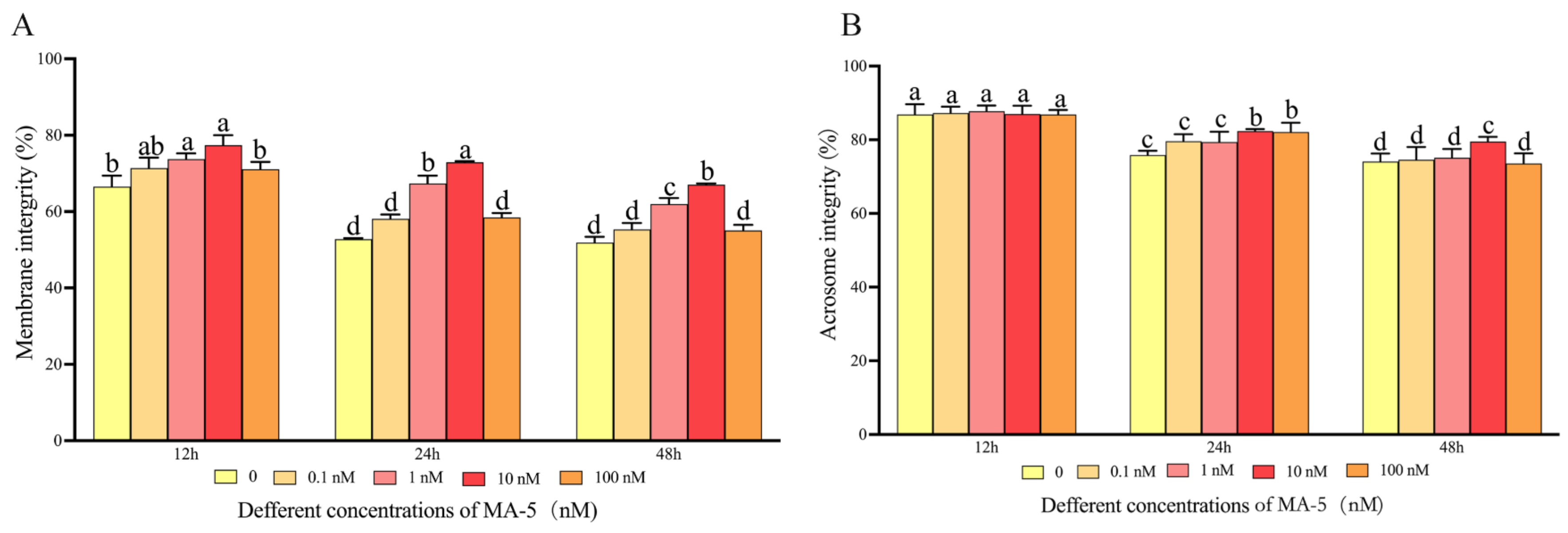
3.2. Mitochonic Acid 5 Affected Ram Sperm Membrane Integrity and Acrosome Integrity
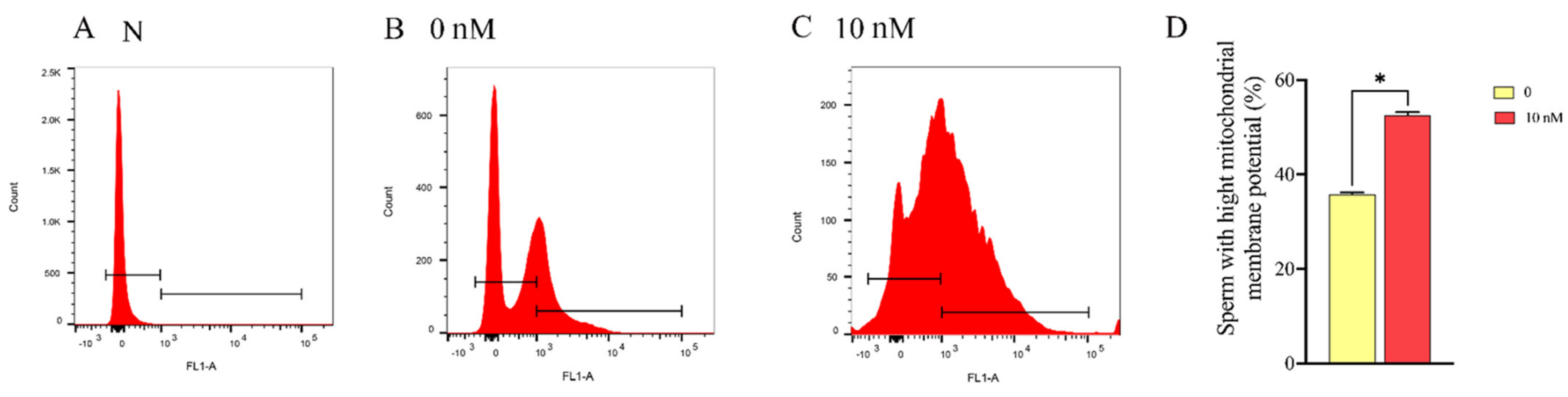
3.3. Mitochonic Acid 5 Increased Sperm Mitochondrial Membrane Potential and ATP Content, Decreasing the ROS Level
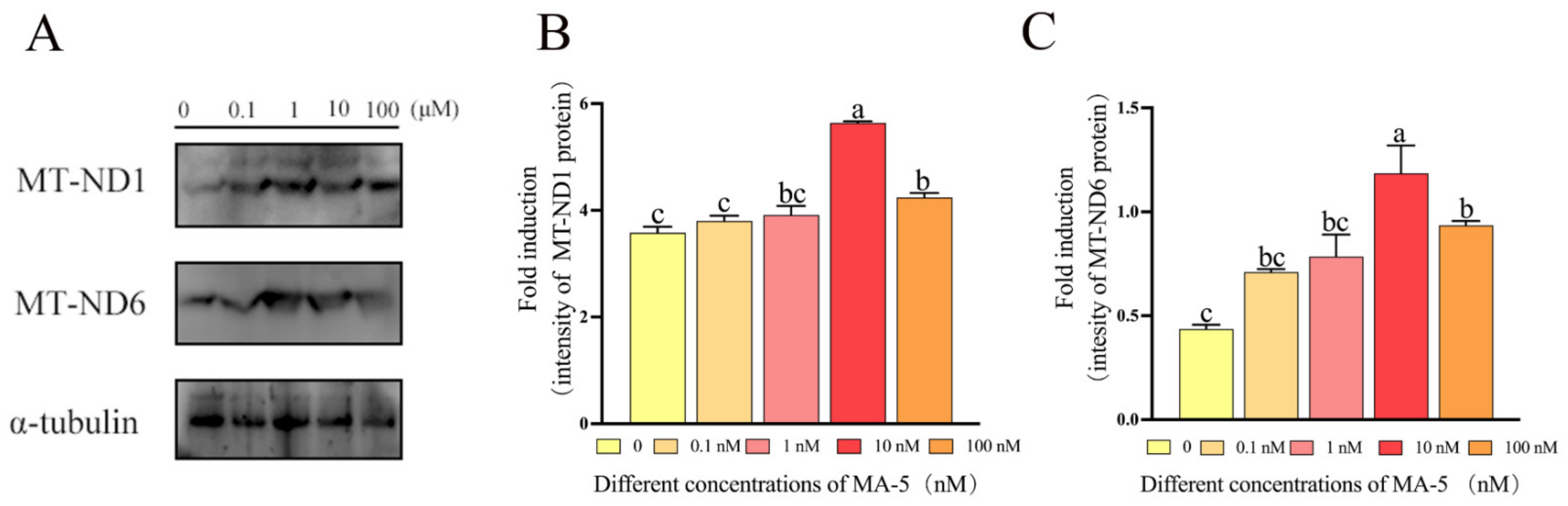
3.4. Mitochonic Acid 5 Increased the Expression of MT-ND1 and MT-ND6 Proteins in Ram Sperm after 48 h of Preservation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Salamon, S.; Maxwell, W.M. Storage of ram semen. Anim. Reprod. Sci. 2000, 62, 77–111. [Google Scholar] [CrossRef] [PubMed]
- Kameni, S.L.; Meutchieye, F.; Ngoula, F. Liquid Storage of Ram Semen: Associated Damages and Improvement. Open J. Anim. Sci. 2021, 11, 473–500. [Google Scholar] [CrossRef]
- Rizkallah, N.; Chambers, C.G.; de Graaf, S.P.; Rickard, J.P. Factors Affecting the Survival of Ram Spermatozoa during Liquid Storage and Options for Improvement. Animals 2022, 12, 244. [Google Scholar] [CrossRef] [PubMed]
- Azimi, G.; Farshad, A.; Farzinpour, A.; Rostamzadeh, J.; Sharafi, M. Evaluation of used Purslane extracts in Tris extenders on cryopreserved goat sperm. Cryobiology 2020, 94, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Salmon, V.M.; Leclerc, P.; Bailey, J.L. Cholesterol-Loaded Cyclodextrin Increases the Cholesterol Content of Goat Sperm to Improve Cold and Osmotic Resistance and Maintain Sperm Function after Cryopreservation. Biol. Reprod. 2016, 94, 85. [Google Scholar] [CrossRef] [PubMed]
- Johnston, S.D.; Satake, N.; Zee, Y.; López-Fernández, C.; Holt, W.V.; Gosálvez, J. Osmotic stress and cryoinjury of koala sperm: An integrative study of the plasma membrane, chromatin stability and mitochondrial function. Reproduction 2012, 143, 787–797. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Li, Z.H.; Dzyuba, B.; Hulak, M.; Rodina, M.; Linhart, O. Evaluating the impacts of osmotic and oxidative stress on common carp (Cyprinus carpio, L.) sperm caused by cryopreservation techniques. Biol. Reprod. 2010, 83, 852–858. [Google Scholar] [CrossRef] [PubMed]
- Han, D.; Williams, E.; Cadenas, E. Mitochondrial respiratory chain-dependent generation of superoxide anion and its release into the intermembrane space. Biochem. J. 2001, 353, 411–416. [Google Scholar] [CrossRef]
- Guthrie, H.D.; Welch, G.R.; Long, J.A. Mitochondrial function and reactive oxygen species action in relation to boar motility. Theriogenology 2008, 70, 1209–1215. [Google Scholar] [CrossRef]
- Kim, S.; Lee, Y.J.; Kim, Y.J. Changes in sperm membrane and ROS following cryopreservation of liquid boar semen stored at 15 °C. Anim. Reprod. Sci. 2011, 124, 118–124. [Google Scholar] [CrossRef]
- Zhu, Z.; Kawai, T.; Umehara, T.; Hoque, S.A.M.; Zeng, W.; Shimada, M. Negative effects of ROS generated during linear sperm motility on gene expression and ATP generation in boar sperm mitochondria. Free. Radic. Biol. Med. 2019, 141, 159–171. [Google Scholar] [CrossRef] [PubMed]
- Ankel-Simons, F.; Cummins, J.M. Misconceptions about mitochondria and mammalian fertilization: Implications for theories on human evolution. Proc. Natl. Acad. Sci. USA 1996, 93, 13859–13863. [Google Scholar] [CrossRef] [PubMed]
- Brand, M.D.; Nicholls, D.G. Assessing mitochondrial dysfunction in cells. Biochem. J. 2011, 435, 297–312. [Google Scholar] [CrossRef] [PubMed]
- Davies, K.M.; Strauss, M.; Daum, B.; Kief, J.H.; Osiewacz, H.D.; Rycovska, A.; Zickermann, V.; Kühlbrandt, W. Macromolecular organization of ATP synthase and complex I in whole mitochondria. Proc. Natl. Acad. Sci. USA 2011, 108, 14121–14126. [Google Scholar] [CrossRef] [PubMed]
- Mukai, C.; Okuno, M. Glycolysis plays a major role for adenosine triphosphate supplementation in mouse sperm flagellar movement. Biol. Reprod. 2004, 71, 540–547. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, J.M.; Shi, L.Z.; Tam, J.; Chandsawangbhuwana, C.; Durrant, B.; Botvinick, E.L.; Berns, M.W. Comparison of glycolysis and oxidative phosphorylation as energy sources for mammalian sperm motility, using the combination of fluorescence imaging, laser tweezers, and real-time automated tracking and trapping. J. Cell. Physiol. 2008, 217, 745–751. [Google Scholar] [CrossRef] [PubMed]
- Travis, A.J.; Foster, J.A.; Rosenbaum, N.A.; Visconti, P.E.; Gerton, G.L.; Kopf, G.S.; Moss, S.B. Targeting of a germ cell-specific type 1 hexokinase lacking a porin-binding domain to the mitochondria as well as to the head and fibrous sheath of murine spermatozoa. Mol. Biol. Cell 1998, 9, 263–276. [Google Scholar] [CrossRef] [PubMed]
- Matsuhashi, T.; Sato, T.; Kanno, S.-i.; Suzuki, T.; Matsuo, A.; Oba, Y.; Kikusato, M.; Ogasawara, E.; Kudo, T.; Suzuki, K.; et al. Mitochonic Acid 5 (MA-5) Facilitates ATP Synthase Oligomerization and Cell Survival in Various Mitochondrial Diseases. EBioMedicine 2017, 20, 27–38. [Google Scholar] [CrossRef]
- Suzuki, T.; Yamaguchi, H.; Kikusato, M.; Matsuhashi, T.; Matsuo, A.; Sato, T.; Oba, Y.; Watanabe, S.; Minaki, D.; Saigusa, D.; et al. Mitochonic Acid 5 (MA-5), a Derivative of the Plant Hormone Indole-3-Acetic Acid, Improves Survival of Fibroblasts from Patients with Mitochondrial Diseases. Tohoku J. Exp. Med. 2015, 236, 225–232. [Google Scholar] [CrossRef]
- Lowell, B.B.; Shulman, G.I. Mitochondrial dysfunction and type 2 diabetes. Science 2005, 307, 384–387. [Google Scholar] [CrossRef]
- Suzuki, T.; Yamaguchi, H.; Kikusato, M.; Hashizume, O.; Nagatoishi, S.; Matsuo, A.; Sato, T.; Kudo, T.; Matsuhashi, T.; Murayama, K.; et al. Mitochonic Acid 5 Binds Mitochondria and Ameliorates Renal Tubular and Cardiac Myocyte Damage. J. Am. Soc. Nephrol. 2016, 27, 1925–1932. [Google Scholar] [CrossRef]
- Umehara, T.; Richards, J.S.; Shimada, M. The stromal fibrosis in aging ovary. Aging 2018, 10, 9–10. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Min, L.; Li, Y.; Lang, Y.; Hoque, S.A.M.; Adetunji, A.O.; Zhu, Z. Beneficial Effect of Proline Supplementation on Goat Spermatozoa Quality during Cryopreservation. Animals 2022, 12, 2626. [Google Scholar] [CrossRef]
- Zhu, Z.; Zeng, Y.; Zeng, W. Cysteine improves boar sperm quality via glutathione biosynthesis during the liquid storage. Anim. Biosci. 2022, 35, 166–176. [Google Scholar] [CrossRef] [PubMed]
- Birben, E.; Sahiner, U.M.; Sackesen, C.; Erzurum, S.; Kalayci, O. Oxidative stress and antioxidant defense. World Allergy Organ. J. 2012, 5, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Allai, L.; Druart, X.; Öztürk, M.; BenMoula, A.; Nasser, B.; El Amiri, B. Protective effects of Opuntia ficus-indica extract on ram sperm quality, lipid peroxidation and DNA fragmentation during liquid storage. Anim. Reprod. Sci. 2016, 175, 1–9. [Google Scholar] [CrossRef]
- Coyan, K.; Baspinar, N.; Bucak, M.N.; Akalin, P.P.; Ataman, M.B.; Omur, A.D.; Gungor, S.; Kucukgunay, S.; Ozkalp, B.; Sariozkan, S. Influence of methionine and dithioerythritol on sperm motility, lipid peroxidation and antioxidant capacities during liquid storage of ram semen. Res. Vet. Sci. 2010, 89, 426–431. [Google Scholar] [CrossRef] [PubMed]
- Tzameli, I. The evolving role of mitochondria in metabolism. Trends Endocrinol. Metab. 2012, 23, 417–419. [Google Scholar] [CrossRef]
- Pieczenik, S.R.; Neustadt, J. Mitochondrial dysfunction and molecular pathways of disease. Exp. Mol. Pathol. 2007, 83, 84–92. [Google Scholar] [CrossRef]
- Aitken, R.J.; Drevet, J.R. The Importance of Oxidative Stress in Determining the Functionality of Mammalian Spermatozoa: A Two-Edged Sword. Antioxidants 2020, 9, 111. [Google Scholar] [CrossRef]
- Friedman, J.R.; Nunnari, J. Mitochondrial form and function. Nature 2014, 505, 335–343. [Google Scholar] [CrossRef]
- Ozimic, S.; Ban-Frangez, H.; Stimpfel, M. Sperm Cryopreservation Today: Approaches, Efficiency, and Pitfalls. Curr. Issues Mol. Biol. 2023, 45, 4716–4734. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; Chen, S.X.; Lei, Q.Y.; Yi, S.Q.; Wu, N.; Wang, Y.L.; Xiao, Z.J.; Wu, H. Mitochonic acid 5 regulates mitofusin 2 to protect microglia. Neural Regen. Res 2021, 16, 1813–1820. [Google Scholar] [CrossRef] [PubMed]
- Strauss, M.; Hofhaus, G.; Schröder, R.R.; Kühlbrandt, W. Dimer ribbons of ATP synthase shape the inner mitochondrial membrane. EMBO J. 2008, 27, 1154–1160. [Google Scholar] [CrossRef] [PubMed]
- Galantino-Homer, H.L.; Zeng, W.X.; Megee, S.O.; Dallmeyer, M.; Voelkl, D.; Dobrinski, I. Effects of 2-hydroxypropyl-beta-cyclodextrin and cholesterol on porcine sperm viability and capacitation status following cold shock or incubation. Mol. Reprod. Dev. 2006, 73, 638–650. [Google Scholar] [CrossRef] [PubMed]
- Baumber, J.; Ball, B.A.; Gravance, C.G.; Medina, V.; Davies-Morel, M.C. The effect of reactive oxygen species on equine sperm motility, viability, acrosomal integrity, mitochondrial membrane potential, and membrane lipid peroxidation. J. Androl. 2000, 21, 895–902. [Google Scholar] [CrossRef] [PubMed]
- Habersetzer, J.; Ziani, W.; Larrieu, I.; Stines-Chaumeil, C.; Giraud, M.F.; Brèthes, D.; Dautant, A.; Paumard, P. ATP synthase oligomerization: From the enzyme models to the mitochondrial morphology. Int. J. Biochem. Cell Biol. 2013, 45, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Piomboni, P.; Focarelli, R.; Stendardi, A.; Ferramosca, A.; Zara, V. The role of mitochondria in energy production for human sperm motility. Int. J. Androl. 2012, 35, 109–124. [Google Scholar] [CrossRef]
- Mimaki, M.; Wang, X.; McKenzie, M.; Thorburn, D.R.; Ryan, M.T. Understanding mitochondrial complex I assembly in health and disease. Biochim. Biophys. Acta 2012, 1817, 851–862. [Google Scholar] [CrossRef]
- Yano, T.; Magnitsky, S.; Ohnishi, T. Characterization of the complex I-associated ubisemiquinone species: Toward the understanding of their functional roles in the electron/proton transfer reaction. Biochim. Biophys. Acta 2000, 1459, 299–304. [Google Scholar] [CrossRef]
- Maranzana, E.; Barbero, G.; Falasca, A.I.; Lenaz, G.; Genova, M.L. Mitochondrial respiratory supercomplex association limits production of reactive oxygen species from complex I. Antioxid. Redox Signal. 2013, 19, 1469–1480. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Different Concentrations of MA-5 (μM) | ||||||
|---|---|---|---|---|---|---|
| Sperm Parameters | Time (h) | 0 | 0.1 | 1 | 10 | 100 |
| No progression (%) | 12 | 39.01 ± 1.82 a | 30.3 ± 0.92 b | 33.58 ± 1.59 ab | 25.05 ± 0.71 c | 34.31 ± 1.71 ab |
| 24 | 59.98 ± 1.04 a | 61.55 ± 1.10 a | 59.44 ± 1.09 a | 49.07 ± 0.61 c | 55.59 ± 0.34 b | |
| 48 | 61.73 ± 2.21 a | 57.99 ± 3.7 a | 49.87 ± 7.91 c | 41.32 ± 11.13 d | 54.95 ± 1.79 b | |
| VSL (μm/s) | 12 | 42.1 ± 1.39 b | 43.42 ± 1.29 b | 42.12 ± 3.28 b | 52.13 ± 1.04 a | 43.22 ± 1.16 b |
| 24 | 23.34 ± 0.90 c | 22.58 ± 2.58 d | 22.56 ± 1.13 d | 33.63 ± 0.88 a | 30.53 ± 0.52 b | |
| 48 | 29.5 ± 4.3 b | 31.05 ± 3.82 b | 39.8 ± 2.36 a | 48.65 ± 4.42 a | 39.8 ± 1.81 a | |
| VCL (μm/s) | 12 | 73.69 ± 1.23 b | 70.32 ± 1.04 b | 74.11 ± 3.30 b | 81.18 ± 1.70 a | 73.97 ± 1.00 a |
| 24 | 62.26 ± 1.23 c | 62.77 ± 0.59 c | 65.67 ± 1.97 c | 81.99 ± 7.30 a | 75.83 ± 2.21 b | |
| 48 | 60.97 ± 3.19 b | 62.67 ± 1.82 b | 72.07 ± 1.76 a | 70.2 ± 2.45 a | 72.08 ± 3.95 a | |
| VAP (μm/s) | 12 | 48.87 ± 1.30 c | 45.50 ± 2.12 c | 49.29 ± 1.30 b | 56.37 ± 2.43 a | 49.15 ± 1.23 bc |
| 24 | 37.44 ± 0.2 c | 37.95 ± 0.53 c | 48.41 ± 0.62 b | 57.17 ± 0.72 a | 51.06 ± 1.99 b | |
| 48 | 36.15 ± 0.88 b | 37.83 ± 4.35 a | 39.69 ± 0.42 a | 45.15 ± 1.15 a | 47.25 ± 0.9 a | |
| LIN (%) | 12 | 53.09 ± 1.37 b | 49.72 ± 1.19 b | 53.51 ± 2.32 b | 60.58 ± 1.78 a | 53.37 ± 0.9 b |
| 24 | 41.66 ± 1.3 d | 42.17 ± 4.43 d | 52.63 ± 3.10 c | 61.3 ± 0.72 a | 55.28 ± 1.07 b | |
| 48 | 40.37 ± 4.08 b | 42.01 ± 2.52 b | 43.91 ± 7.2 ab | 49.62 ± 1.15 a | 51.42 ± 4.47 a | |
| STR (%) | 12 | 78.50 ± 1.15 bc | 75.13 ± 5.23 c | 78.92 ± 2.29 b | 85.99 ± 1.24 a | 78.78 ± 1.21 b |
| 24 | 67.07 ± 0.40 e | 67.58 ± 0.36 d | 78.04 ± 0.51 c | 86.80 ± 7.40 a | 80.69 ± 4.49 b | |
| 48 | 65.78 ± 3.29 d | 67.46 ± 1.82 c | 69.32 ± 0.52 b | 75.04 ± 0.11 a | 76.89 ± 4.54 a | |
| WOB (%) | 12 | 60.23 ± 1.30 bc | 56.86 ± 1.17 c | 60.65 ± 3.41 b | 67.72 ± 1.75 a | 60.51 ± 0.89 b |
| 24 | 48.80 ± 3.10 c | 49.31 ± 0.47 c | 59.77 ± 3.40 b | 68.10 ± 2.57 a | 62.40 ± 0.7 b | |
| 48 | 47.51 ± 7.47 c | 49.19 ± 0.52 c | 51.05 ± 3.4 b | 56.71 ± 1.57 a | 58.62 ± 0.97 a | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, R.; Liu, L.; Min, L.; Adetunji, A.O.; Kou, X.; Zhou, K.; Zhu, Z. Mitochonic Acid 5 Increases Ram Sperm Quality by Improving Mitochondrial Function during Storage at 4 °C. Animals 2024, 14, 368. https://doi.org/10.3390/ani14030368
Wang R, Liu L, Min L, Adetunji AO, Kou X, Zhou K, Zhu Z. Mitochonic Acid 5 Increases Ram Sperm Quality by Improving Mitochondrial Function during Storage at 4 °C. Animals. 2024; 14(3):368. https://doi.org/10.3390/ani14030368
Chicago/Turabian StyleWang, Ruyuan, Luwei Liu, Lingjiang Min, Adedeji O. Adetunji, Xin Kou, Kaifeng Zhou, and Zhendong Zhu. 2024. "Mitochonic Acid 5 Increases Ram Sperm Quality by Improving Mitochondrial Function during Storage at 4 °C" Animals 14, no. 3: 368. https://doi.org/10.3390/ani14030368