
Effects of Dietary Plant Protein Replacement with Insect and Poultry By-Product Meals on the Liver Health and Serum Metabolites of Sea Bream (Sparus aurata) and Sea Bass (Dicentrarchus labrax)
 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Test Diets
2.2. Fish Husbandry and Sampling
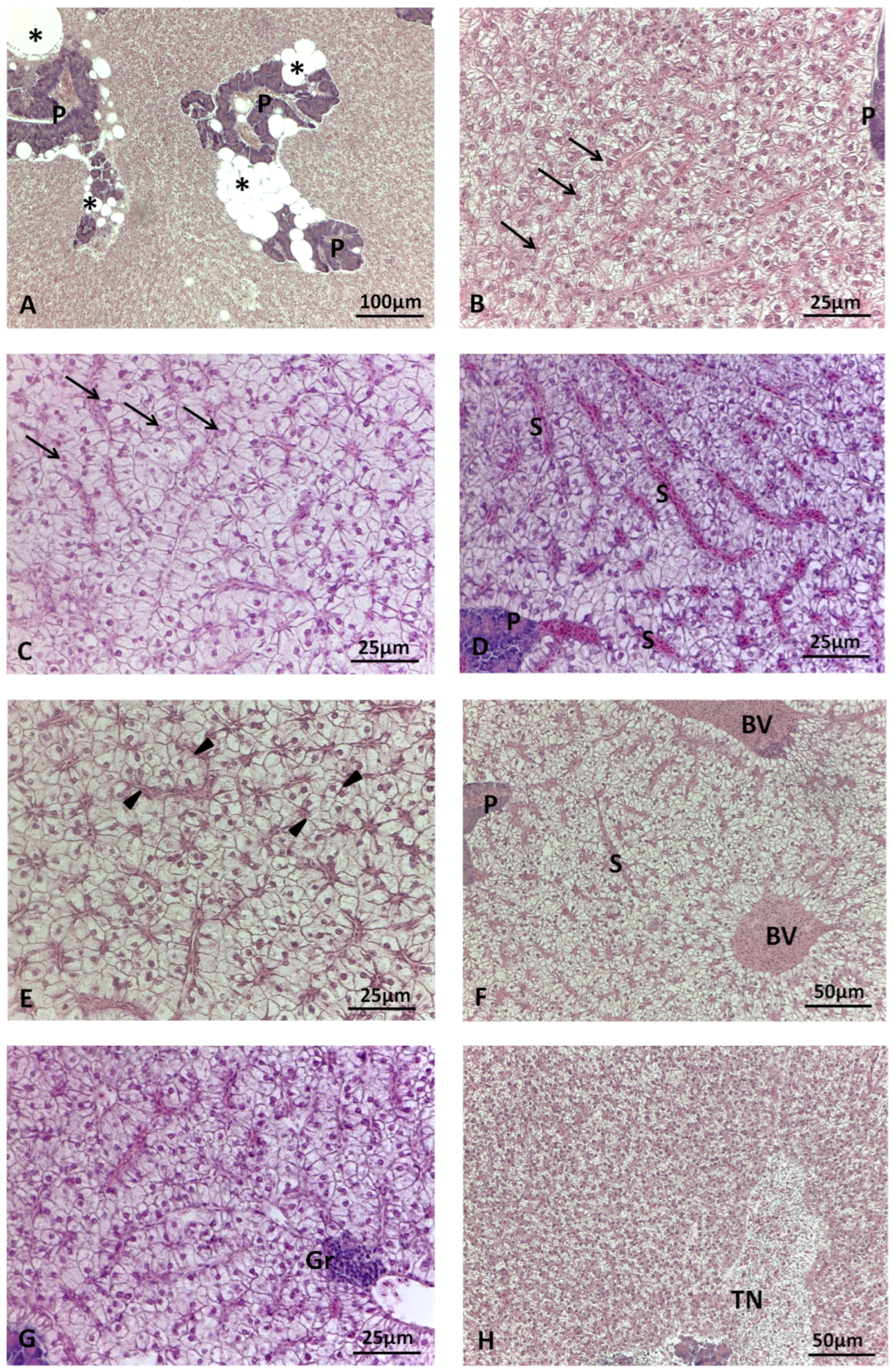
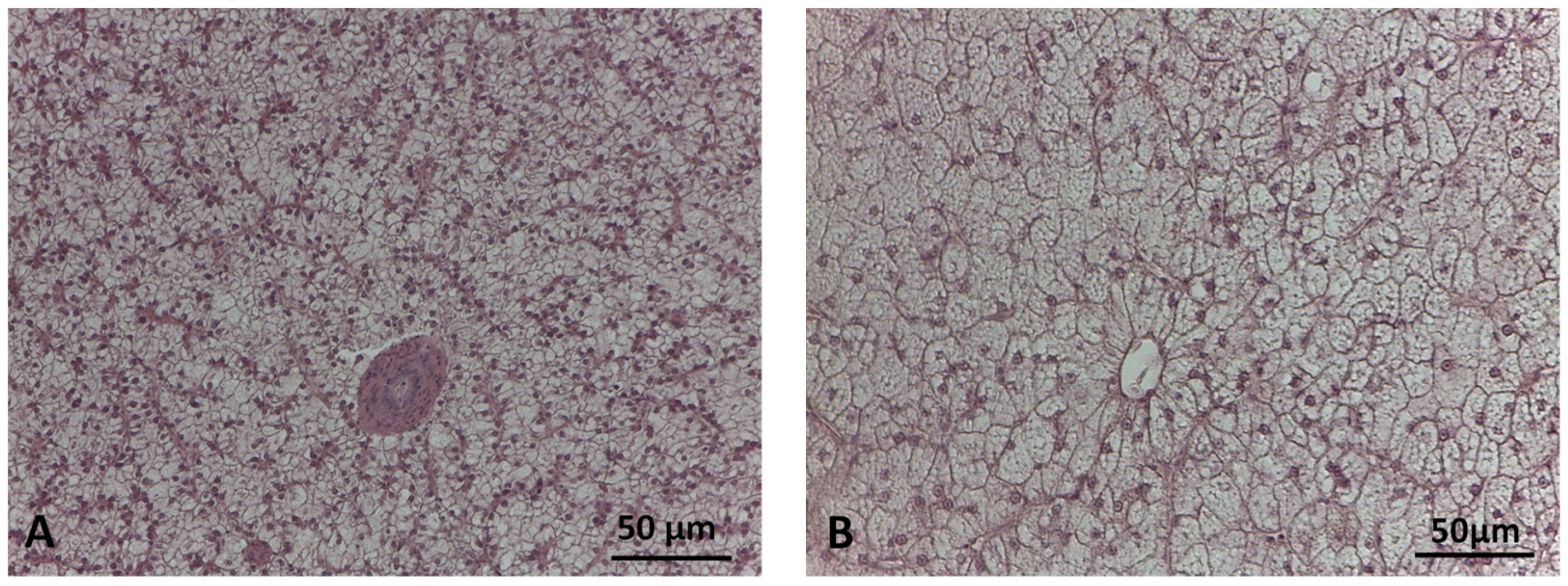
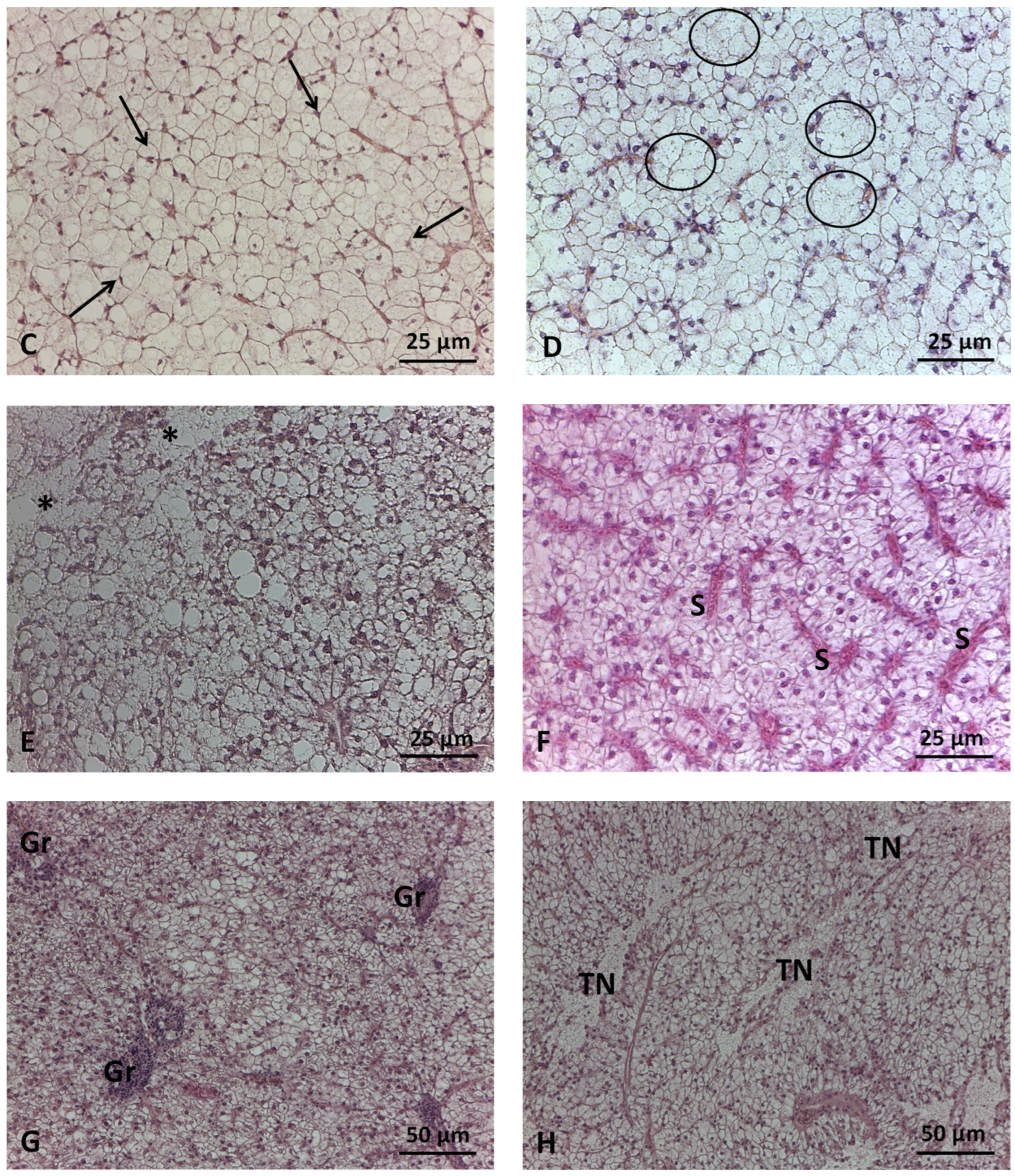
2.3. Liver Histological Analyses
2.3.1. Histomorphometric Analysis
2.3.2. Histopathological Evaluation
2.3.3. Liver Index Calculation
- i—ith fish;
- sj—the score assigned to each histological alteration (j) from 0 to 6, considering the frequency percentage within each field, as described above;
- wj—the importance factor, expressing the severity of the histological alteration (j);
- a—the number of observed fields;
- sj—the maximum score attainable for each histological alteration.
2.4. Blood Biochemistry Analyses
2.5. Statistical Analysis
2.6. Ethical Approval
3. Results
3.1. Biometric Measurements and Fish Condition

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Test Diets | Kruskal–Wallis Test | |||||
|---|---|---|---|---|---|---|
| CV | H40 | P40 | H10P30 | |||
| Sea bream | TL(cm) | 21.9 ± 0.6 | 22.2 ± 0.8 | 22.3 ± 0.6 | 22.3 ± 0.3 | n.s. |
| BW final (g) | 175.2 ± 17.1 b | 188.8 ± 24.8 a | 190.3 ± 15.7 a | 192.6 ± 9.5 a | p < 0.05 | |
| K (g/cm3) | 1.67 ± 0.12 | 1.72 ± 0.13 | 1.70 ± 0.06 | 1.74 ± 0.07 | n.s. | |
| HSI (%) | 1.29 ± 0.27 ab | 1.34 ± 0.14 ab | 1.27 ± 0.18 b | 1.50 ± 0.19 a | p < 0.01 | |
| Sea bass | TL (cm) | 22.8 ± 1.02 | 23.3 ± 94 | 23.2 ± 1.38 | 23.5 ± 0.85 | n.s. |
| BW final (g) | 160.7 ± 28.2 | 159.3 ± 20.4 | 154.5 ± 27.9 | 171.2 ± 23.9 | n.s. | |
| K (g/cm3) | 1.31 ± 0.09 a | 1.26 ± 0.08 ab | 1.22 ± 0.04 b | 1.32 ± 0.06 a | p < 0.001 | |
| HSI (%) | 1.80 ± 0.29 | 1.60 ± 0.17 | 1.67 ± 0.32 | 1.52 ± 0.32 | n.s. | |
3.2. Liver Histology
3.2.1. Histomorphometry
| Lipid Accumulation | Kruskal–Wallis Test | ||||
|---|---|---|---|---|---|
| Mild | Moderate | Severe | |||
| Sea bream | Hepatocyte area (µm2) | 97.4 ± 13.4 a | 165.4 ± 37.9 b | - | p < 0.001 |
| Nucleus area (µm2) | 16.5 ± 1.8 a | 13.0 ± 1.2 b | - | p < 0.001 | |
| N/H ratio | 0.17 ± 0.03 a | 0.08 ± 0.02 b | - | p < 0.001 | |
| Sea bass | Hepatocyte area (µm2) | 119.1 ± 17.1 a | 205.3 ± 40.1 b | 372.1 ± 175.1 c | p < 0.001 |
| Nucleus area (µm2) | 15.5 ± 0.36 a | 14.6 ± 1.4 a | 12.32 ± 1.3 b | p < 0.001 | |
| N/H ratio | 0.13 ± 0.02 a | 0.07 ± 0.02 b | 0.04 ± 0.01 c | p < 0.001 | |
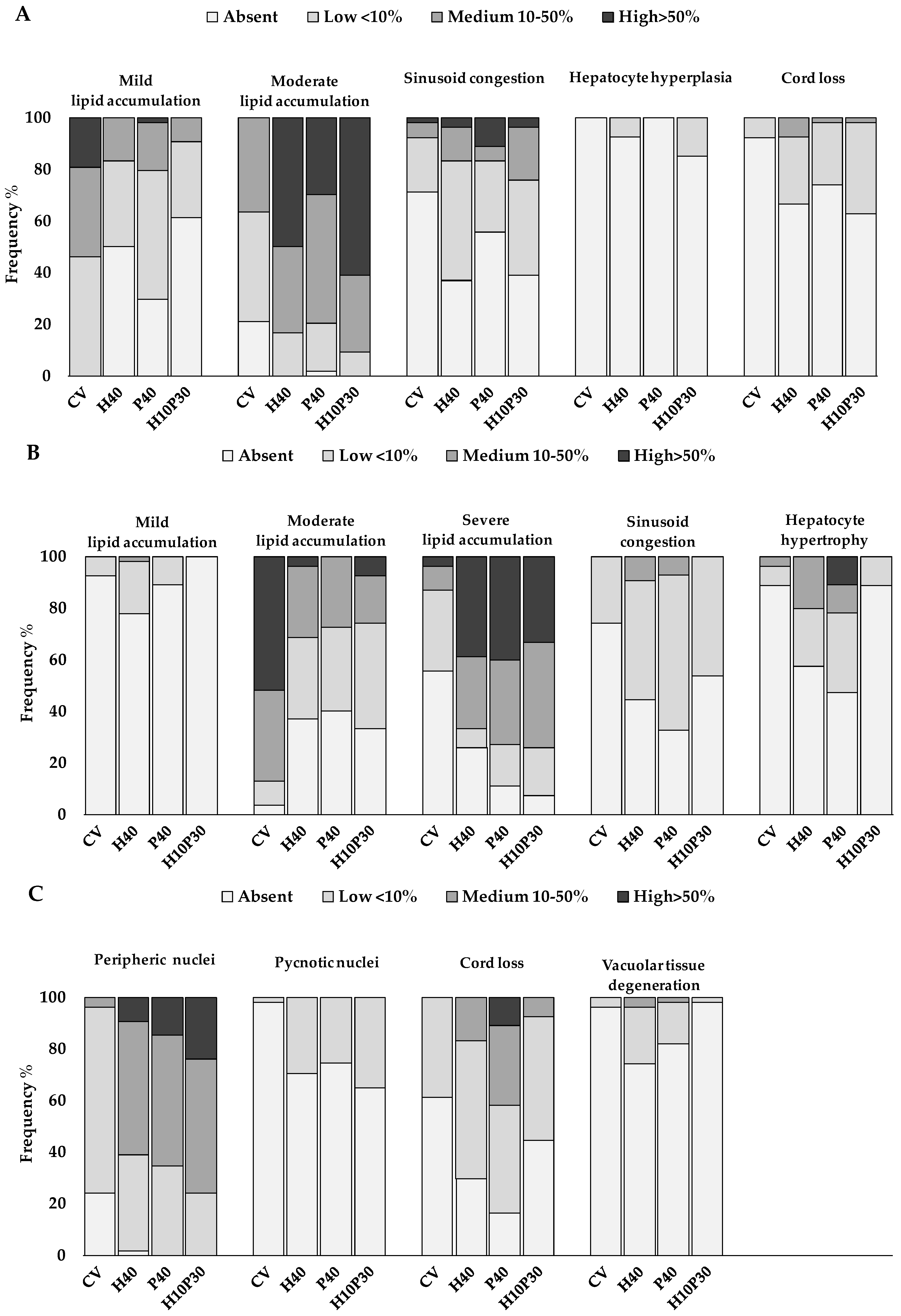
3.2.2. Histopathology
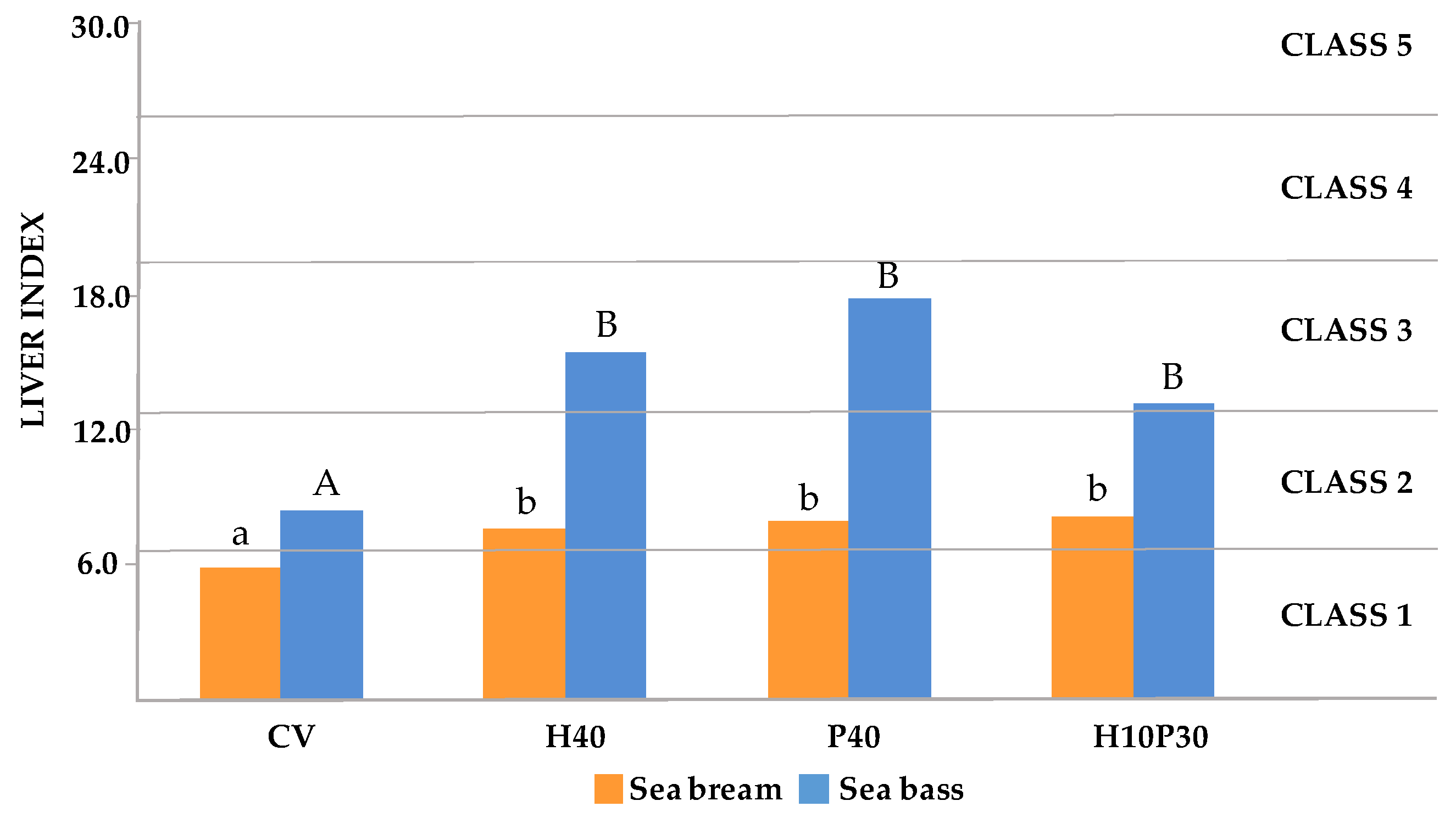
3.3. Liver Index
3.3.1. Formula and Scoring Scheme
3.3.2. Liver Health Calculation
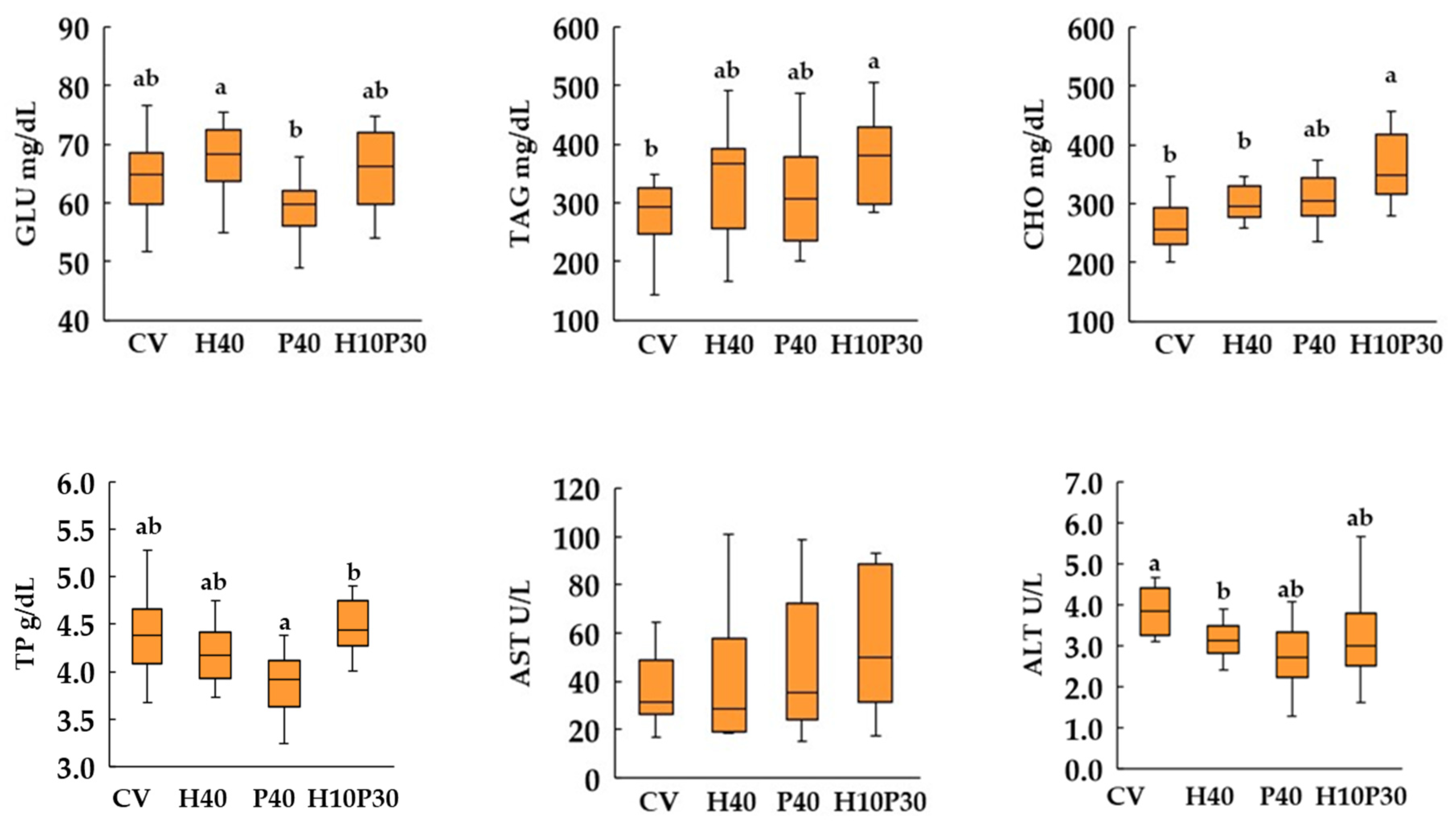
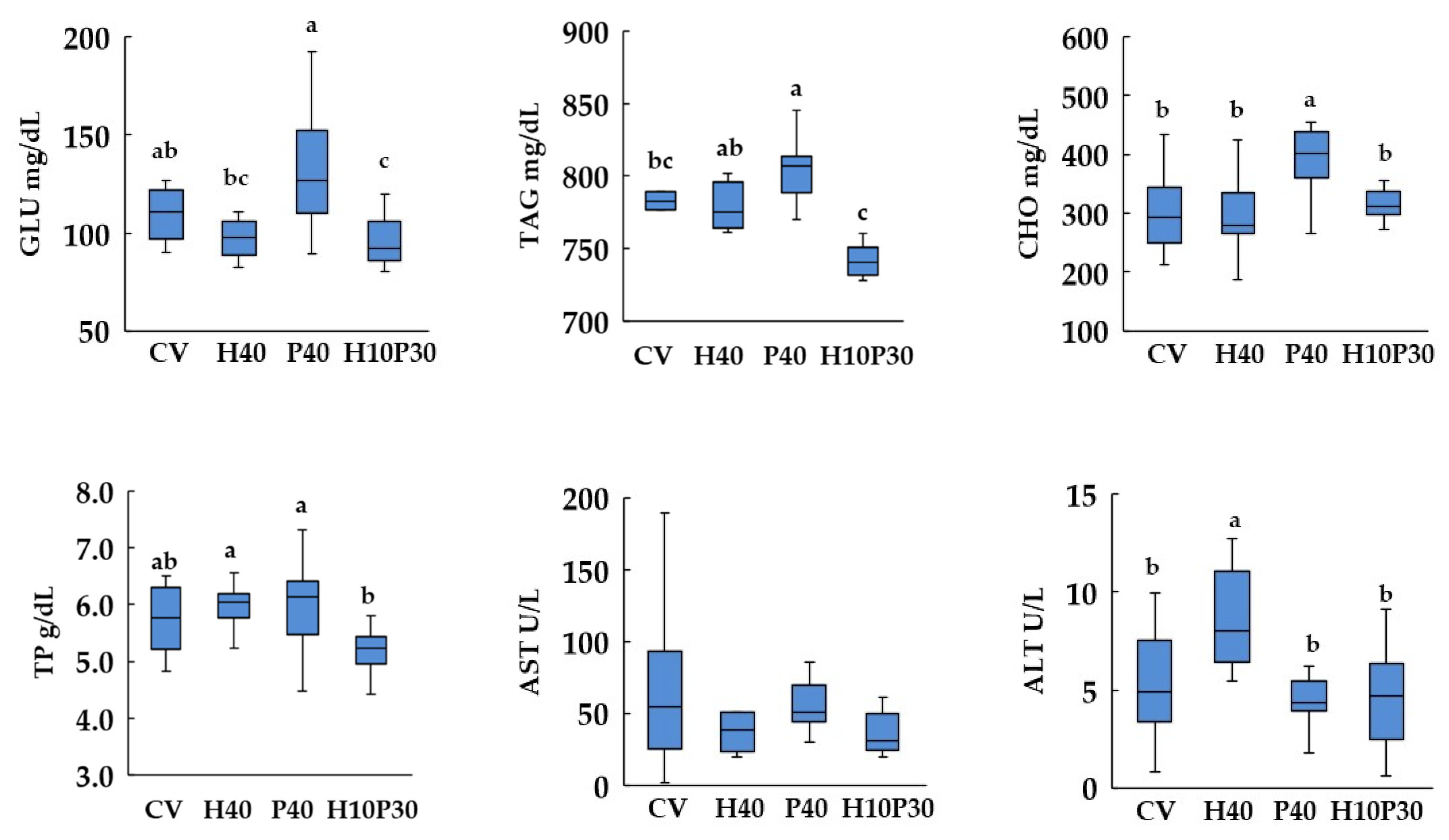
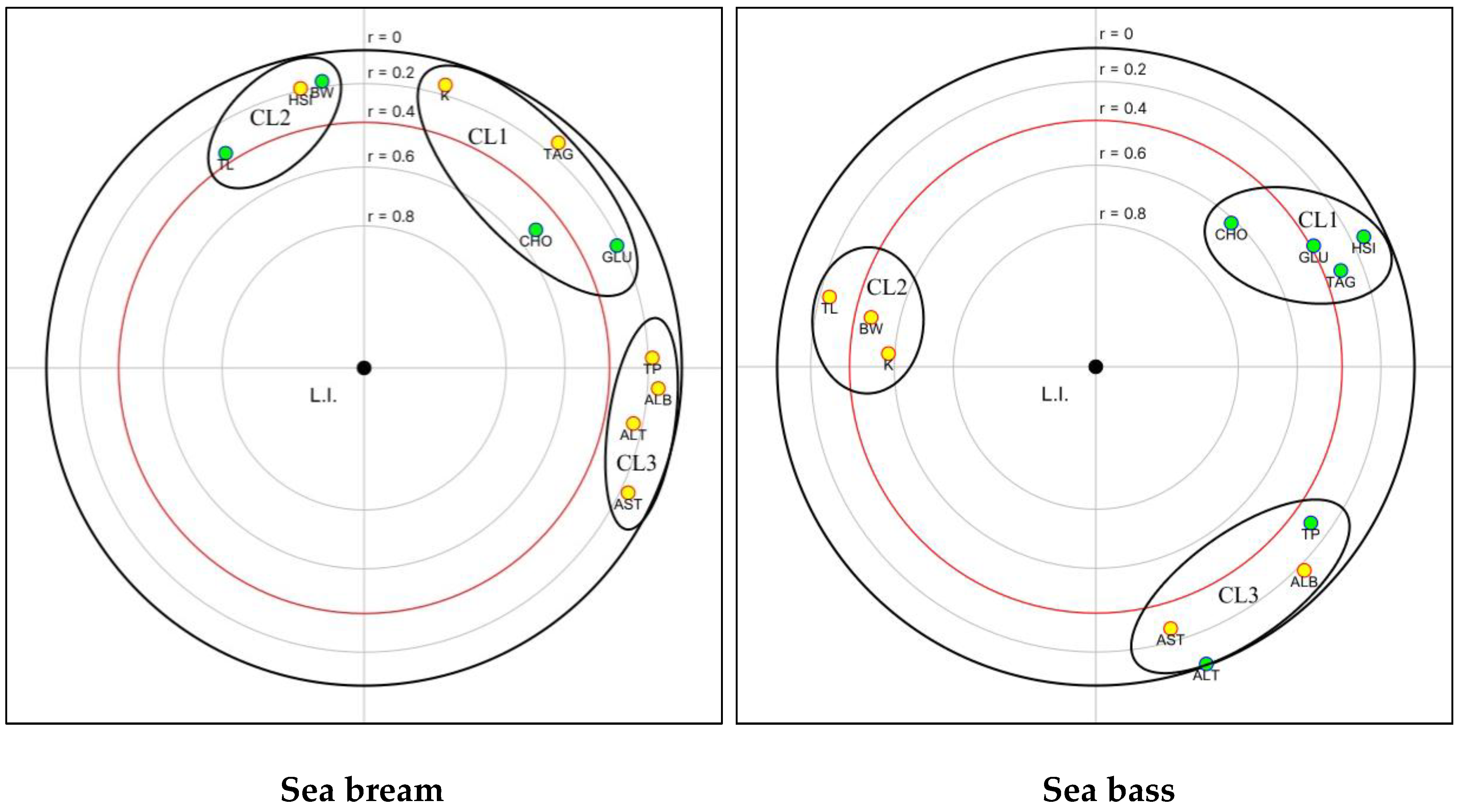
3.4. Blood Biochemistry
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. The State of World Fisheries and Aquaculture 2020. Sustainability in Action; FAO: Rome, Italy, 2000; ISBN 978–92–5-132692–3. [Google Scholar]
- FAO. The State of World Fisheries and Aquaculture 2022. Towards Blue Transformation; FAO: Rome, Italy, 2022; ISBN 978–92–5-136364-5. [Google Scholar]
- UN DESA. The Sustainable Development Goals Report 2023: Special Edition—July 2023; UN DESA: New York, NY, USA, 2023; ISBN 978-92-1-101460-0. [Google Scholar]
- Tacon, A.G.J.; Metian, M.; McNevin, A.A. Future Feeds: Suggested Guidelines for Sustainable Development. Rev. Fish. Sci. Aquac. 2022, 30, 135–142. [Google Scholar] [CrossRef]
- Hua, K.; Cobcroft, J.M.; Cole, A.; Condon, K.; Jerry, D.R.; Mangott, A.; Praeger, C.; Vucko, M.J.; Zeng, C.; Zenger, K.; et al. The Future of Aquatic Protein: Implications for Protein Sources in Aquaculture Diets. One Earth 2019, 1, 316–329. [Google Scholar] [CrossRef]
- Gatlin, D.M.; Barrows, F.T.; Brown, P.; Dabrowski, K.; Gaylord, T.G.; Hardy, R.W.; Herman, E.; Hu, G.; Krogdahl, Å.; Nelson, R.; et al. Expanding the Utilization of Sustainable Plant Products in Aquafeeds: A Review. Aquac. Res. 2007, 38, 551–579. [Google Scholar] [CrossRef]
- Glencross, B.D.; Baily, J.; Berntssen, M.H.G.; Hardy, R.; MacKenzie, S.; Tocher, D.R. Risk Assessment of the Use of Alternative Animal and Plant Raw Material Resources in Aquaculture Feeds. Rev. Aquac. 2020, 12, 703–758. [Google Scholar] [CrossRef]
- Colombo, S.M. Physiological Considerations in Shifting Carnivorous Fishes to Plant-Based Diets. In Fish Physiology; Benfey, T.J., Farrell, A.P., Brauner, C.J., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; Volume 38, pp. 53–82. ISBN 9780128207949. [Google Scholar] [CrossRef]
- Aragão, C.; Gonçalves, A.T.; Costas, B.; Azeredo, R.; Xavier, M.J.; Engrola, S. Alternative Proteins for Fish Diets: Implications beyond Growth. Animals 2022, 12, 1211. [Google Scholar] [CrossRef] [PubMed]
- Cottrell, R.S.; Blanchard, J.L.; Halpern, B.S.; Metian, M.; Froehlich, H.E. Global Adoption of Novel Aquaculture Feeds Could Substantially Reduce Forage Fish Demand by 2030. Nat. Food 2020, 1, 301–308. [Google Scholar] [CrossRef]
- Colombo, S.M.; Roy, K.; Mraz, J.; Wan, A.H.L.; Davies, S.J.; Tibbetts, S.M.; Øverland, M.; Francis, D.S.; Rocker, M.M.; Gasco, L.; et al. Towards Achieving Circularity and Sustainability in Feeds for Farmed Blue Foods. Rev. Aquac. 2023, 15, 1115–1141. [Google Scholar] [CrossRef]
- Cao, L.; Halpern, B.S.; Troell, M.; Short, R.; Zeng, C.; Jiang, Z.; Liu, Y.; Zou, C.; Liu, C.; Liu, S.; et al. Vulnerability of Blue Foods to Human-Induced Environmental Change. Nat. Sustain. 2023, 6, 1186–1198. [Google Scholar] [CrossRef]
- Van Riel, A.; Nederlof, M.A.J.; Chary, K.; Wiegertjes, G.F.; De Boer, I.J.M. Feed-food Competition in Global Aquaculture: Current Trends and Prospects. Rev. Aquac. 2023, 15, 1142–1158. [Google Scholar] [CrossRef]
- Meeker, D.L. Essential Rendering: All about the Animal by-Products Industry; National Renderers Association, Fats and Proteins Research Foundation, Animal Protein Producers Industry: Alexandria, VA, USA, 2006; ISBN 978-0-9654660-3-5. [Google Scholar]
- Lim, C.; Webster, C.D.; Lee, C.-S. Alternative Protein Sources in Aquaculture Diets; Haworth Press: New York, NY, USA, 2008; ISBN 978-1-56022-148-7. [Google Scholar]
- Nogales-Mérida, S.; Gobbi, P.; Józefiak, D.; Mazurkiewicz, J.; Dudek, K.; Rawski, M.; Kierończyk, B.; Józefiak, A. Insect Meals in Fish Nutrition. Rev. Aquac. 2019, 11, 1080–1103. [Google Scholar] [CrossRef]
- Hawkey, K.J.; Lopez-Viso, C.; Brameld, J.M.; Parr, T.; Salter, A.M. Insects: A Potential Source of Protein and Other Nutrients for Feed and Food. Annu. Rev. Anim. Biosci. 2021, 9, 333–354. [Google Scholar] [CrossRef] [PubMed]
- Alfiko, Y.; Xie, D.; Astuti, R.T.; Wong, J.; Wang, L. Insects as a Feed Ingredient for Fish Culture: Status and Trends. Aquac. Fish. 2022, 7, 166–178. [Google Scholar] [CrossRef]
- Gougbedji, A.; Detilleux, J.; Lalèyè, P.; Francis, F.; Caparros Megido, R. Can Insect Meal Replace Fishmeal? A Meta-Analysis of the Effects of Black Soldier Fly on Fish Growth Performances and Nutritional Values. Animals 2022, 12, 1700. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.S.; Small, B.C.; Hardy, R. Insect Lipid in Fish Nutrition: Recent Knowledge and Future Application in Aquaculture. Rev. Aquac. 2023, 15, 1664–1685. [Google Scholar] [CrossRef]
- Arena, R.; Manuguerra, S.; Curcuraci, E.; Cusimano, M.; Lo Monaco, D.; Di Bella, C.; Santulli, A.; Messina, C.M. Fisheries and Aquaculture By-Products Modulate Growth, Body Composition, and Omega-3 Polyunsaturated Fatty Acid Content in Black Soldier Fly (Hermetia illucens) Larvae. Front. Anim. Sci. 2023, 4, 1204767. [Google Scholar] [CrossRef]
- Maroušek, J.; Strunecký, O.; Maroušková, A. Insect Rearing on Biowaste Represents a Competitive Advantage for Fish Farming. Rev. Aquac. 2023, 15, 965–975. [Google Scholar] [CrossRef]
- Galkanda-Arachchige, H.S.C.; Wilson, A.E.; Davis, D.A. Success of Fishmeal Replacement through Poultry By-product Meal in Aquaculture Feed Formulations: A Meta-analysis. Rev. Aquac. 2020, 12, 1624–1636. [Google Scholar] [CrossRef]
- Hua, K. A Meta-Analysis of the Effects of Replacing Fish Meals with Insect Meals on Growth Performance of Fish. Aquaculture 2021, 530, 735732. [Google Scholar] [CrossRef]
- Tran, H.Q.; Von Siebenthal, E.W.; Luce, J.-B.; Nguyen, T.T.; Tomčala, A.; Stejskal, V.; Janssens, T. Complementarity of Insect Meal and Poultry By-Product Meal as Replacement for Fishmeal Can Sustain the Production Performance of European Perch (Perca fluviatilis), Reduce Economic Fish-in Fish-out Ratio and Food-Feed Competition, and Influence the Environmental Indices. Aquaculture 2024, 579, 740166. [Google Scholar] [CrossRef]
- Gebremichael, A.; Kucska, B.; Ardó, L.; Biró, J.; Berki, M.; Lengyel-Kónya, É.; Tömösközi-Farkas, R.; Egessa, R.; Müller, T.; Gyalog, G.; et al. Physiological Response of Grower African Catfish to Dietary Black Soldier Fly and Mealworm Meal. Animals 2023, 13, 968. [Google Scholar] [CrossRef]
- Karapanagiotidis, I.T.; Psofakis, P.; Mente, E.; Malandrakis, E.; Golomazou, E. Effect of Fishmeal Replacement by Poultry By-Product Meal on Growth Performance, Proximate Composition, Digestive Enzyme Activity, Haematological Parameters and Gene Expression of Gilthead Seabream (Sparus aurata ). Aquacult. Nutr. 2019, 25, 3–14. [Google Scholar] [CrossRef]
- Sabbagh, M.; Schiavone, R.; Brizzi, G.; Sicuro, B.; Zilli, L.; Vilella, S. Poultry By-Product Meal as an Alternative to Fish Meal in the Juvenile Gilthead Seabream (Sparus aurata) Diet. Aquaculture 2019, 511, 734220. [Google Scholar] [CrossRef]
- Psofakis, P.; Meziti, A.; Berillis, P.; Mente, E.; Kormas, K.A.; Karapanagiotidis, I.T. Effects of Dietary Fishmeal Replacement by Poultry By-Product Meal and Hydrolyzed Feather Meal on Liver and Intestinal Histomorphology and on Intestinal Microbiota of Gilthead Seabream (Sparus aurata). Appl. Sci. 2021, 11, 8806. [Google Scholar] [CrossRef]
- Magalhães, R.; Sánchez-López, A.; Leal, R.S.; Martínez-Llorens, S.; Oliva-Teles, A.; Peres, H. Black Soldier Fly (Hermetia illucens) Pre-Pupae Meal as a Fish Meal Replacement in Diets for European Seabass (Dicentrarchus labrax). Aquaculture 2017, 476, 79–85. [Google Scholar] [CrossRef]
- Abdel-Tawwab, M.; Khalil, R.H.; Metwally, A.A.; Shakweer, M.S.; Khallaf, M.A.; Abdel-Latif, H.M.R. Effects of Black Soldier Fly (Hermetia Illucens L.) Larvae Meal on Growth Performance, Organs-Somatic Indices, Body Composition, and Hemato-Biochemical Variables of European Sea Bass, Dicentrarchus labrax. Aquaculture 2020, 522, 735136. [Google Scholar] [CrossRef]
- Karapanagiotidis, I.T.; Neofytou, M.C.; Asimaki, A.; Daskalopoulou, E.; Psofakis, P.; Mente, E.; Rumbos, C.I.; Athanassiou, C.G. Fishmeal Replacement by Full-Fat and Defatted Hermetia illucens Prepupae Meal in the Diet of Gilthead Seabream (Sparus aurata). Sustainability 2023, 15, 786. [Google Scholar] [CrossRef]
- Fabrikov, D.; Vargas-García, M.D.C.; Barroso, F.G.; Sánchez-Muros, M.J.; Cacua Ortíz, S.M.; Morales, A.E.; Cardenete, G.; Tomás-Almenar, C.; Melenchón, F. Effect on Intermediary Metabolism and Digestive Parameters of the High Substitution of Fishmeal with Insect Meal in Sparus aurata Feed. Insects 2021, 12, 965. [Google Scholar] [CrossRef]
- Mastoraki, M.; Katsika, L.; Enes, P.; Guerreiro, I.; Kotzamanis, Y.P.; Gasco, L.; Chatzifotis, S.; Antonopoulou, E. Insect Meals in Feeds for Juvenile Gilthead Seabream (Sparus aurata): Effects on Growth, Blood Chemistry, Hepatic Metabolic Enzymes, Body Composition and Nutrient Utilization. Aquaculture 2022, 561, 738674. [Google Scholar] [CrossRef]
- Mastoraki, M.; Mollá Ferrándiz, P.; Vardali, S.C.; Kontodimas, D.C.; Kotzamanis, Y.P.; Gasco, L.; Chatzifotis, S.; Antonopoulou, E. A Comparative Study on the Effect of Fish Meal Substitution with Three Different Insect Meals on Growth, Body Composition and Metabolism of European Sea Bass (Dicentrarchus labrax L.). Aquaculture 2020, 528, 735511. [Google Scholar] [CrossRef]
- Mastoraki, M.; Panteli, N.; Kotzamanis, Y.P.; Gasco, L.; Antonopoulou, E.; Chatzifotis, S. Nutrient Digestibility of Diets Containing Five Different Insect Meals in Gilthead Sea Bream (Sparus aurata) and European Sea Bass (Dicentrarchus labrax). Anim. Feed. Sci. Technol. 2022, 292, 115425. [Google Scholar] [CrossRef]
- Bousdras, T.; Feidantsis, K.; Panteli, N.; Chatzifotis, S.; Piccolo, G.; Gasco, L.; Gai, F.; Antonopoulou, E. Dietary Tenebrio Molitor Larvae Meal Inclusion Exerts Tissue-Specific Effects on Cellular, Metabolic, and Antioxidant Status in European Sea Bass (Dicentrarchus labrax) and Gilthead Seabream (Sparus aurata). Aquac. Nutr. 2022, 2022. [Google Scholar] [CrossRef] [PubMed]
- Gai, F.; Cusimano, G.M.; Maricchiolo, G.; Caccamo, L.; Caimi, C.; Macchi, E.; Meola, M.; Perdichizzi, A.; Tartarisco, G.; Gasco, L. Defatted Black Soldier Fly Meal in Diet for Grow-Out Gilthead Seabream (Sparus aurata L. 1758): Effects on Growth Performance, Gill Cortisol Level, Digestive Enzyme Activities, and Intestinal Histological Structure. Aquac. Res. 2023, 2023. [Google Scholar] [CrossRef]
- Di Rosa, A.R.; Caccamo, L.; Pansera, L.; Oteri, M.; Chiofalo, B.; Maricchiolo, G. Influence of Hermetia Illucens Larvae Meal Dietary Inclusion on Growth Performance, Gut Histological Traits and Stress Parameters in Sparus aurata. Animals 2023, 13, 339. [Google Scholar] [CrossRef] [PubMed]
- Anedda, R.; Melis, R.; Palomba, A.; Vitangeli, I.; Biosa, G.; Braca, A.; Antonini, M.; Moroni, F.; Rimoldi, S.; Terova, G.; et al. Balanced Replacement of Fish Meal with Hermetia illucens Meal Allows Efficient Hepatic Nutrient Metabolism and Increases Fillet Lipid Quality in Gilthead Sea Bream (Sparus aurata). Aquaculture 2023, 576, 739862. [Google Scholar] [CrossRef]
- Basto, A.; Valente, L.M.P.; Sousa, V.; Conde-Sieira, M.; Soengas, J.L. Total Fishmeal Replacement by Defatted Tenebrio Molitor Larvae Meal Induces Alterations in Intermediary Metabolism of European Sea Bass (Dicentrarchus labrax). J. Anim. Sci. 2023, 101, skad040. [Google Scholar] [CrossRef]
- Randazzo, B.; Zarantoniello, M.; Cardinaletti, G.; Cerri, R.; Giorgini, E.; Belloni, A.; Contò, M.; Tibaldi, E.; Olivotto, I. Hermetia Illucens and Poultry By-Product Meals as Alternatives to Plant Protein Sources in Gilthead Seabream (Sparus aurata) Diet: A Multidisciplinary Study on Fish Gut Status. Animals 2021, 11, 677. [Google Scholar] [CrossRef]
- Pulido-Rodriguez, L.F.; Cardinaletti, G.; Secci, G.; Randazzo, B.; Bruni, L.; Cerri, R.; Olivotto, I.; Tibaldi, E.; Parisi, G. Appetite Regulation, Growth Performances and Fish Quality Are Modulated by Alternative Dietary Protein Ingredients in Gilthead Sea Bream (Sparus aurata) Culture. Animals 2021, 11, 1919. [Google Scholar] [CrossRef]
- Pleić, I.L.; Bušelić, I.; Messina, M.; Hrabar, J.; Žuvić, L.; Talijančić, I.; Žužul, I.; Pavelin, T.; Anđelić, I.; Pleadin, J.; et al. A Plant-Based Diet Supplemented with Hermetia illucens Alone or in Combination with Poultry by-Product Meal: One Step Closer to Sustainable Aquafeeds for European Seabass. J. Anim. Sci. Biotechnol. 2022, 13, 77. [Google Scholar] [CrossRef]
- Bakke, A.M.; Glover, C.; Krogdahl, Å. Feeding, Digestion and Absorption of Nutrients. In Fish Physiology; Grosell, M., Farrell, A.P., Brauner, C.J., Eds.; Elsevier: Amsterdam, The Netherlands, 2010; Volume 30, pp. 57–110. ISBN 9780123749826. [Google Scholar] [CrossRef]
- Bruslé, J.; Gonzàlez, I.; Anadon, G. The Structure and Function of Fish Liver. In Fish Morphology; Datta Munshi, J.S., Dutta, H.M., Eds.; Routledge: London, UK, 2017; pp. 77–93. ISBN 978-0-203-75599-0. [Google Scholar]
- Roberts, R.J. Nutritional Pathology. In Fish Nutrition; Academic Press: London, UK, 2022; pp. 823–855. ISBN 978-0-12-819587-1. [Google Scholar]
- Sirri, R.; Sarli, G.; Bianco, C.; Bonaldo, A.; Gatta, P.P.; Fontanillas, R.; De Vico, G.; Carella, F.; Brachelente, C.; Parma, L.; et al. Retrospective Study of Pathology-Based Investigative Techniques for the Assessment of Diet-Induced Changes in Liver and Intestine of Flatfish. Ital. J. Anim. Sci. 2018, 17, 518–529. [Google Scholar] [CrossRef]
- Rašković, B.; Berillis, P. Special Issue on the Histopathology of Aquatic Animals. Appl. Sci. 2022, 12, 971. [Google Scholar] [CrossRef]
- Caballero, M.J.; López-Calero, G.; Socorro, J.; Roo, F.J.; Izquierdo, M.S.; Férnandez, A.J. Combined Effect of Lipid Level and Fish Meal Quality on Liver Histology of Gilthead Seabream (Sparus aurata). Aquaculture 1999, 179, 277–290. [Google Scholar] [CrossRef]
- Russell, P.M.; Davies, S.J.; Gouveia, A.; Tekinay, A.A. Influence of Dietary Starch Source on Liver Morphology in Juvenile Cultured European Sea Bass (Dicentrarchus labrax L.): Dietary Starch in Sea Bass Diets. Aquac. Res. 2001, 32, 306–314. [Google Scholar] [CrossRef]
- Benedito-Palos, L.; Navarro, J.C.; Sitjà-Bobadilla, A.; Gordon Bell, J.; Kaushik, S.; Pérez-Sánchez, J. High Levels of Vegetable Oils in Plant Protein-Rich Diets Fed to Gilthead Sea Bream (Sparus aurata L.): Growth Performance, Muscle Fatty Acid Profiles and Histological Alterations of Target Tissues. Br. J. Nutr. 2008, 100, 992–1003. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo-Silva, A.; Rocha, E.; Dias, J.; Silva, P.; Rema, P.; Gomes, E.; Valente, L.M.P. Partial Replacement of Fish Oil by Soybean Oil on Lipid Distribution and Liver Histology in European Sea Bass (Dicentrarchus labrax) and Rainbow Trout (Oncorhynchus mykiss) Juveniles. Aquac. Nutr. 2005, 11, 147–155. [Google Scholar] [CrossRef]
- Caballero, M.J.; Izquierdo, M.S.; Kjørsvik, E.; Fernández, A.J.; Rosenlund, G. Histological Alterations in the Liver of Sea Bream, Sparus aurata L., Caused by Short- or Long-term Feeding with Vegetable Oils. Recovery of Normal Morphology after Feeding Fish Oil AS the Sole Lipid Source. J. Fish. Dis. 2004, 27, 531–541. [Google Scholar] [CrossRef]
- Kokou, F.; Sarropoulou, E.; Cotou, E.; Rigos, G.; Henry, M.; Alexis, M.; Kentouri, M. Effects of Fish Meal Replacement by a Soybean Protein on Growth, Histology, Selected Immune and Oxidative Status Markers of Gilthead Sea Bream, Sparus aurata. J. World Aquac. Soc. 2015, 46, 115–128. [Google Scholar] [CrossRef]
- Baeza-Ariño, R.; Martínez-Llorens, S.; Nogales-Mérida, S.; Jover-Cerda, M.; Tomás-Vidal, A. Study of Liver and Gut Alterations in Sea Bream, Sparus aurata L., Fed a Mixture of Vegetable Protein Concentrates. Aquac. Res. 2016, 47, 460–471. [Google Scholar] [CrossRef]
- Saleh, N.E. Assessment of Sesame Meal as a Soybean Meal Replacement in European Sea Bass (Dicentrarchus labrax) Diets Based on Aspects of Growth, Amino Acid Profiles, Haematology, Intestinal and Hepatic Integrity and Macroelement Contents. Fish. Physiol. Biochem. 2020, 46, 861–879. [Google Scholar] [CrossRef]
- Bernet, D.; Schmidt, H.; Meier, W.; Burkhardt-Holm, P.; Wahli, T. Histopathology in Fish: Proposal for a Protocol to Assess Aquatic Pollution. J. Fish. Dis. 1999, 22, 25–34. [Google Scholar] [CrossRef]
- Bernet, D.; Schmidt-Posthaus, H.; Wahli, T.; Burkhardt-Holm, P. Evaluation of Two Monitoring Approaches to Assess Effects of Waste Water Disposal on Histological Alterations in Fish. Hydrobiologia 2004, 524, 53–66. [Google Scholar] [CrossRef]
- Zimmerli, S.; Bernet, D.; Burkhardt-Holm, P.; Schmidt-Posthaus, H.; Vonlanthen, P.; Wahli, T.; Segner, H. Assessment of Fish Health Status in Four Swiss Rivers Showing a Decline of Brown Trout Catches. Aquat. Sci. 2007, 69, 11–25. [Google Scholar] [CrossRef]
- Van Dyk, J.C.; Cochrane, M.J.; Wagenaar, G.M. Liver Histopathology of the Sharptooth Catfish Clarias gariepinus as a Biomarker of Aquatic Pollution. Chemosphere 2012, 87, 301–311. [Google Scholar] [CrossRef]
- Beegam, A.; Lopes, M.; Fernandes, T.; Jose, J.; Barreto, A.; Oliveira, M.; Soares, A.M.V.M.; Trindade, T.; Thomas, S.; Pereira, M.L. Multiorgan Histopathological Changes in the Juvenile Seabream Sparus aurata as a Biomarker for Zinc Oxide Particles Toxicity. Env. Sci. Pollut. Res. 2020, 27, 30907–30917. [Google Scholar] [CrossRef]
- Santos, R.M.B.; Monteiro, S.M.V.; Cortes, R.M.V.; Pacheco, F.A.L.; Fernandes, L.F.S. Seasonal Differences in Water Pollution and Liver Histopathology of Iberian Barbel (Luciobarbus bocagei) and Douro Nase (Pseudochondrostoma duriense) in an Agricultural Watershed. Water 2022, 14, 444. [Google Scholar] [CrossRef]
- Rašković, B.; Jarić, I.; Koko, V.; Spasić, M.; Dulić, Z.; Marković, Z.; Poleksić, V. Histopathological Indicators: A Useful Fish Health Monitoring Tool in Common Carp (Cyprinus carpio Linnaeus, 1758) Culture. Open Life Sci. 2013, 8, 975–985. [Google Scholar] [CrossRef]
- Saraiva, A.; Costa, J.; Serrão, J.; Cruz, C.; Eiras, J.C. A Histology-Based Fish Health Assessment of Farmed Seabass (Dicentrarchus Labrax L.). Aquaculture 2015, 448, 375–381. [Google Scholar] [CrossRef]
- Pacorig, V.; Galeotti, M.; Beraldo, P. Multiparametric Semi-Quantitative Scoring System for the Histological Evaluation of Marine Fish Larval and Juvenile Quality. Aquac. Rep. 2022, 26, 101285. [Google Scholar] [CrossRef]
- Rey, A.L.; Asín, J.; Ruiz Zarzuela, I.; Luján, L.; Iregui, C.A.; De Blas, I. A Proposal of Standardization for Histopathological Lesions to Characterize Fish Diseases. Rev. Aquac. 2020, 12, 2304–2315. [Google Scholar] [CrossRef]
- Raskovic, B.; Stankovic, M.; Markovic, Z.; Poleksic, V. Histological Methods in the Assessment of Different Feed Effects on Liver and Intestine of Fish. J. Agric. Sci. BGD 2011, 56, 87–100. [Google Scholar] [CrossRef]
- Mandrioli, L.; Sirri, R.; Gatta, P.P.; Morandi, F.; Sarli, G.; Parma, L.; Fontanillas, R.; Bonaldo, A. Histomorphologic Hepatic Features and Growth Performances of Juvenile Senegalese Sole (Solea senegalensis) Fed Isogenertic Practical Diets with Variable Protein/Lipid Levels: Histomorphologic Hepatic Features and Growth Performances in Senegalese Sole. J. Appl. Ichthyol. 2012, 28, 628–632. [Google Scholar] [CrossRef]
- Wassef, E.A.; Wahbi, O.M.; Saqr, E.M.; Saleh, N.E. Response of European Seabass (Dicentrarchus labrax) to Canola Oil Diets: Effect on Growth Performance, Fish Health and Liver and Intestine Histomorphology. Aquacult. Int. 2016, 24, 1073–1088. [Google Scholar] [CrossRef]
- Carvalho, M.; Montero, D.; Torrecillas, S.; Castro, P.; Zamorano, M.J.; Izquierdo, M. Hepatic Biochemical, Morphological and Molecular Effects of Feeding Microalgae and Poultry Oils to Gilthead Sea Bream (Sparus aurata). Aquaculture 2021, 532, 736073. [Google Scholar] [CrossRef]
- Tefal, E.; Jauralde, I.; Tomás-Vidal, A.; Martínez-Llorens, S.; Peñaranda, D.S.; Jover-Cerdá, M. New Organic Raw Materials for Gilthead Seabream (Sparus aurata) Feeding and the Effects on Growth, Nutritive Parameters, Digestibility, and Histology. Fishes 2023, 8, 330. [Google Scholar] [CrossRef]
- Valaroutsou, E.; Voudanta, E.; Mente, E.; Berillis, P. A Microscope and Image Analysis Study of the Liver and Exocrine Pancreas of Sea Bream Sparus aurata Fed Different Diets. Int. J. Zool. Res. 2013, 3, 55–58. [Google Scholar]
- Bilen, A.M.; Bilen, S. Effect of Diet on the Fatty Acids Composition of Cultured Sea Bass (Dicentrarchus labrax) Liver Tissues and Histology Compared with Wild Sea Bass Caught in Eagean Sea. Mar. Sci. Tech. Bull. 2013, 2, 15–21. [Google Scholar]
- FAO FishStatJ. FishStatJ—Software for Fishery and Aquaculture Statistical Time Series. FAO, Statistics and Information Service, FAO Fisheries Department, Fishery Information, Data and Statistics Unit, Rome, Italy. 2023. Available online: https://www.fao.org/fishery/statistics/software/fishstatj/en (accessed on 15 September 2023).
- Latimer, G.W. (Ed.) Official Methods of Analysis of AOAC International, 20th ed.; AOAC International: Gaithersburg, MD, USA, 2016; ISBN 978-0-935584-87-5. [Google Scholar]
- Burja, A.M.; Armenta, R.E.; Radianingtyas, H.; Barrow, C.J. Evaluation of Fatty Acid Extraction Methods for Thraustochytrium Sp. ONC-T18. J. Agric. Food Chem. 2007, 55, 4795–4801. [Google Scholar] [CrossRef] [PubMed]
- Di Marco, P.; Priori, A.; Finoia, M.G.; Massari, A.; Mandich, A.; Marino, G. Physiological Responses of European Sea Bass Dicentrarchus labrax to Different Stocking Densities and Acute Stress Challenge. Aquaculture 2008, 275, 319–328. [Google Scholar] [CrossRef]
- Cardinaletti, G.; Di Marco, P.; Daniso, E.; Messina, M.; Donadelli, V.; Finoia, M.G.; Petochi, T.; Fava, F.; Faccenda, F.; Contò, M.; et al. Growth and Welfare of Rainbow Trout (Oncorhynchus mykiss) in Response to Graded Levels of Insect and Poultry By-Product Meals in Fishmeal-Free Diets. Animals 2022, 12, 1698. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: https://www.R-project.org/ (accessed on 15 September 2023).
- Franks, B.; Ewell, C.; Jacquet, J. Animal Welfare Risks of Global Aquaculture. Sci. Adv. 2021, 7, eabg0677. [Google Scholar] [CrossRef]
- Oliva-Teles, A.; Enes, P.; Peres, H. Replacing Fishmeal and Fish Oil in Industrial Aquafeeds for Carnivorous Fish. In Woodhead Publishing Series in Food Science, Technology and Nutrition, Feed and Feeding Practices in Aquaculture; Davis, D.A., Ed.; Woodhead Publishing: Sawston, UK, 2015; pp. 203–233. ISBN 9780081005064. [Google Scholar] [CrossRef]
- Panserat, S.; Marandel, L.; Seiliez, I.; Skiba-Cassy, S. New Insights on Intermediary Metabolism for a Better Understanding of Nutrition in Teleosts. Annu. Rev. Anim. Biosci. 2019, 7, 195–220. [Google Scholar] [CrossRef]
- Ciji, A.; Akhtar, M.S. Stress Management in Aquaculture: A Review of Dietary Interventions. Rev. Aquac. 2021, 13, 2190–2247. [Google Scholar] [CrossRef]
- Ballester-Lozano, G.F.; Benedito-Palos, L.; Estensoro, I.; Sitjà-Bobadilla, A.; Kaushik, S.; Pérez-Sánchez, J. Comprehensive Biometric, Biochemical and Histopathological Assessment of Nutrient Deficiencies in Gilthead Sea Bream Fed Semi-Purified Diets. Br. J. Nutr. 2015, 114, 713–726. [Google Scholar] [CrossRef]
- Bonvini, E.; Bonaldo, A.; Mandrioli, L.; Sirri, R.; Dondi, F.; Bianco, C.; Fontanillas, R.; Mongile, F.; Gatta, P.P.; Parma, L. Effects of Feeding Low Fishmeal Diets with Increasing Soybean Meal Levels on Growth, Gut Histology and Plasma Biochemistry of Sea Bass. Animal 2018, 12, 923–930. [Google Scholar] [CrossRef] [PubMed]
- Peres, H.; Santos, S.; Oliva-Teles, A. Selected Plasma Biochemistry Parameters in Gilthead Seabream (Sparus aurata) Juveniles. J. Appl. Ichthyol. 2013, 29, 630–636. [Google Scholar] [CrossRef]
- Peres, H.; Santos, S.; Oliva-Teles, A. Blood Chemistry Profile as Indicator of Nutritional Status in European Seabass (Dicentrarchus labrax). Fish. Physiol. Biochem. 2014, 40, 1339–1347. [Google Scholar] [CrossRef]
- Couto, A.; Kortner, T.M.; Penn, M.; Bakke, A.M.; Krogdahl, Å.; Oliva-Teles, A. Effects of Dietary Soy Saponins and Phytosterols on Gilthead Sea Bream (Sparus aurata) during the on-Growing Period. Anim. Feed. Sci. Technol. 2014, 198, 203–214. [Google Scholar] [CrossRef]
- Kaushik, S.J.; Covès, D.; Dutto, G.; Blanc, D. Almost Total Replacement of Fish Meal by Plant Protein Sources in the Diet of a Marine Teleost, the European Seabass, Dicentrarchus Labrax. Aquaculture 2004, 230, 391–404. [Google Scholar] [CrossRef]
- Dias, J.; Alvarez, M.J.; Arzel, J.; Corraze, G.; Diez, A.; Bautista, J.M.; Kaushik, S.J. Dietary Protein Source Affects Lipid Metabolism in the European Seabass (Dicentrarchus labrax). Comp. Biochem. Physiol. Part. A Mol. Integr. Physiol. 2005, 142, 19–31. [Google Scholar] [CrossRef]
- Ekmann, K.S.; Dalsgaard, J.; Holm, J.; Campbell, P.J.; Skov, P.V. Effects of Dietary Energy Density and Digestible Protein:Energy Ratio on de Novo Lipid Synthesis from Dietary Protein in Gilthead Sea Bream (Sparus aurata) Quantified with Stable Isotopes. Br. J. Nutr. 2013, 110, 1771–1781. [Google Scholar] [CrossRef]
- Messina, M.; Piccolo, G.; Tulli, F.; Messina, C.M.; Cardinaletti, G.; Tibaldi, E. Lipid Composition and Metabolism of European Sea Bass (Dicentrarchus labrax L.) Fed Diets Containing Wheat Gluten and Legume Meals as Substitutes for Fish Meal. Aquaculture 2013, 376–379, 6–14. [Google Scholar] [CrossRef]
- Castro, C.; Corraze, G.; Pérez-Jiménez, A.; Larroquet, L.; Cluzeaud, M.; Panserat, S.; Oliva-Teles, A. Dietary Carbohydrate and Lipid Source Affect Cholesterol Metabolism of European Sea Bass ( Dicentrarchus labrax ) Juveniles. Br. J. Nutr. 2015, 114, 1143–1156. [Google Scholar] [CrossRef] [PubMed]
- Castro, C.; Corraze, G.; Firmino-Diógenes, A.; Larroquet, L.; Panserat, S.; Oliva-Teles, A. Regulation of Glucose and Lipid Metabolism by Dietary Carbohydrate Levels and Lipid Sources in Gilthead Sea Bream Juveniles. Br. J. Nutr. 2016, 116, 19–34. [Google Scholar] [CrossRef] [PubMed]
- Torrecillas, S.; Robaina, L.; Caballero, M.J.; Montero, D.; Calandra, G.; Mompel, D.; Karalazos, V.; Kaushik, S.; Izquierdo, M.S. Combined Replacement of Fishmeal and Fish Oil in European Sea Bass (Dicentrarchus labrax): Production Performance, Tissue Composition and Liver Morphology. Aquaculture 2017, 474, 101–112. [Google Scholar] [CrossRef]
- Rašković, B.; Čičovački, S.; Ćirić, M.; Marković, Z.; Poleksić, V. Integrative Approach of Histopathology and Histomorphometry of Common Carp (Cyprinus Carpio L.) Organs as a Marker of General Fish Health State in Pond Culture. Aquac. Res. 2016, 47, 3455–3463. [Google Scholar] [CrossRef]
- Spisni, E.; Tugnoli, M.; Ponticelli, A.; Mordenti, T.; Tomasi, V. Hepatic Steatosis in Artificially Fed Marine Teleosts. J. Fish. Dis. 1998, 21, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Montero, D.; Izquierdo, M. Welfare and Health of Fish Fed Vegetable Oils as Alternative Lipid Sources to Fish Oil. In Fish Oil Replacement and Alternative Lipid Sources in Aquaculture Feeds; Turchini, G., Ng, W.-K., Tocher, D., Eds.; CRC Press: Boca Raton, FL, USA, 2010; pp. 439–485. ISBN 978-1-4398-0862-7. [Google Scholar]
- Geay, F.; Ferraresso, S.; Zambonino-Infante, J.L.; Bargelloni, L.; Quentel, C.; Vandeputte, M.; Kaushik, S.; Cahu, C.L.; Mazurais, D. Effects of the Total Replacement of Fish-Based Diet with Plant-Based Diet on the Hepatic Transcriptome of Two European Sea Bass (Dicentrarchus labrax) Half-Sibfamilies Showing Different Growth Rates with the Plant-Based Diet. BMC Genom. 2011, 12, 522. [Google Scholar] [CrossRef]
- Coz-Rakovac, R.; Strunjak-Perovic1, I.; Hacmanjek, M.; Popovic, N.T.; Lipej, Z.; Sostaric, B. Blood Chemistry and Histological Properties of Wild and Cultured Sea Bass (Dicentrarchus labrax) in the North Adriatic Sea. Vet. Res. Commun. 2005, 29, 677–687. [Google Scholar] [CrossRef]
- Di Marco, P.; Petochi, T.; Marino, G.; Priori, A.; Finoia, M.G.; Tomassetti, P.; Porrello, S.; Giorgi, G.; Lupi, P.; Bonelli, A.; et al. Insights into Organic Farming of European Sea Bass Dicentrarchus labrax and Gilthead Sea Bream Sparus aurata through the Assessment of Environmental Impact, Growth Performance, Fish Welfare and Product Quality. Aquaculture 2017, 471, 92–105. [Google Scholar] [CrossRef]
- Pavelin, T.; Kević, N.; Restović, I.; Bočina, I. Histological and Biochemical Features of the Digestive System in the Cage-Reared Gilthead Sea Bream (Sparus aurata). Int. J. Biotechnol. Recent. Adv. 2018, 2, 51–56. [Google Scholar] [CrossRef]
- Peres, H.; Oliva-Teles, A. Effect of Dietary Lipid Level on Growth Performance and Feed Utilization by European Sea Bass Juveniles (Dicentrarchus labrax). Aquaculture 1999, 179, 325–334. [Google Scholar] [CrossRef]
- Dias, J.; Alvarez, M.J.; Diez, A.; Arzel, J.; Corraze, G.; Bautista, J.M.; Kaushik, S.J. Regulation of Hepatic Lipogenesis by Dietary Protein/Energy in Juvenile European Seabass (Dicentrarchus labrax). Aquaculture 1998, 161, 169–186. [Google Scholar] [CrossRef]
- Marchi, A.; Bonaldo, A.; Di Biase, A.; Cerri, R.; Scicchitano, D.; Nanetti, E.; Candela, M.; Picone, G.; Capozzi, F.; Dondi, F.; et al. Towards a Free Wild-Caught Fishmeal, Fish Oil and Soy Protein in European Sea Bass Diet Using by-Products from Fishery and Aquaculture. Aquaculture 2023, 573, 739571. [Google Scholar] [CrossRef]








| Ingredient Composition (g 100 g−1) | CV | H40 | P40 | H10P30 |
|---|---|---|---|---|
| Vegetable protein mix 1 | 69.0 | 36.6 | 35.4 | 35.4 |
| Hermetia illucens meal 2 | - | 32.4 | - | 8.1 |
| Poultry by-product meal 3 | - | - | 27.5 | 20.6 |
| Feeding stimulants 4 | 5.5 | 5.5 | 5.5 | 5.5 |
| Wheat meal * | 0.4 | 4.5 | 5.6 | 5.5 |
| Whole pea * | 3.0 | 6.0 | 9.0 | 8.8 |
| Fish oil 5 | 6.2 | 6.2 | 6.2 | 6.2 |
| Vegetable oil mix 6 | 11.4 | 5.4 | 8.2 | 7.4 |
| Vit. & Min. Premix 7 | 0.4 | 0.4 | 0.4 | 0.4 |
| Sodium phosphate | 1.6 | 1.0 | 0.3 | 0.2 |
| L-Lysine 8 | 0.5 | 0.2 | 0.1 | 0.1 |
| DL-Methionine 9 | 0.5 | 0.3 | 0.3 | 0.3 |
| Celite | 1.5 | 1.5 | 1.5 | 1.5 |
| Proximate composition (% as fed): | ||||
| Moisture | 6.7 | 6.0 | 7.1 | 8.7 |
| Protein (Nx6.25) | 45.0 | 45.2 | 45.1 | 45.1 |
| Total lipid | 20.4 | 20.4 | 20.3 | 20.4 |
| Ash | 5.8 | 6.6 | 7.8 | 7.7 |
| Chitin # | 0.02 | 1.54 | 0.02 | 0.40 |
| Energy (MJ/kg) | 21.5 | 21.0 | 21.7 | 21.5 |
| Reaction Pattern | Histological Alteration (j) | Importance Factor (w) |
|---|---|---|
| Circulatory disturbance | Sinusoid congestion | w = 1 |
| Blood vessel congestion | w = 1 | |
| Hemorrhages | w = 2 | |
| Regressive changes | Mild-to-moderate lipid accumulation | w = 1 |
| Severe lipid accumulation | w = 2 | |
| Peripheric nuclei | w = 1 | |
| Pycnotic nuclei | w = 2 | |
| Cord loss | w = 2 | |
| Vacuolar tissue degeneration | w = 2 | |
| Tissue necrosis | w = 3 | |
| Progressive changes | Hepatocyte hyperplasia | w = 2 |
| Hepatocytes hypertrophy | w = 2 | |
| Bile duct hypertrophy | w = 1 | |
| Inflammation | Granulocyte infiltration | w = 2 |
| MMc occurrence | w = 1 | |
| Tumor | Benign tumor | w = 2 |
| Malignant tumor | w = 3 |
| Test Diets | Kruskal–Wallis Test | |||||
|---|---|---|---|---|---|---|
| CV | H40 | P40 | H10P30 | |||
| Sea bream | Hepatocyte area (µm2) | 109.9 ± 26.8 a | 146.5 ± 33.1 b | 135.7 ± 38.8 b | 177.9 ± 50.5 c | p < 0.001 |
| Nucleus area (µm2) | 16.2 ± 2.4 a | 13.8 ± 1.2 b | 14.1 ± 1.9 b | 12.6 ± 1.2 c | p < 0.001 | |
| N/H ratio | 0.16 ± 0.05 a | 0.10 ± 0.03 b | 0.12 ± 0.05 b | 0.08 ± 0.03 c | p < 0.001 | |
| Sea bass | Hepatocyte area (µm2) | 199.2 ± 44.9 a | 277.4 ± 89.6 b | 428.1 ± 23.5 c | 247.6 ± 56.5 b | p < 0.001 |
| Nucleus area (µm2) | 15.5 ± 0.9 a | 13.32 ± 1.6 b | 11.87 ± 1.0 c | 13.26 ± 1.5 b | p < 0.001 | |
| N/H ratio | 0.08 ± 0.02 a | 0.06 ± 0.03 b | 0.03 ± 0.01 c | 0.06 ± 0.02 b | p < 0.001 | |
| Scoring Scheme | Description | |
|---|---|---|
| Class 1 | ≤6.5 | Normal liver structure |
| Class 2 | 6.6–13.0 | Normal liver structure with slight histological alterations |
| Class 3 | 13.1–19.5 | Normal liver structure with moderate histological alterations |
| Class 4 | 19.6–25.9 | Pronounced alterations of liver structure |
| Class 5 | >26 | Severe alterations of liver structure |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Donadelli, V.; Di Marco, P.; Mandich, A.; Finoia, M.G.; Cardinaletti, G.; Petochi, T.; Longobardi, A.; Tibaldi, E.; Marino, G. Effects of Dietary Plant Protein Replacement with Insect and Poultry By-Product Meals on the Liver Health and Serum Metabolites of Sea Bream (Sparus aurata) and Sea Bass (Dicentrarchus labrax). Animals 2024, 14, 241. https://doi.org/10.3390/ani14020241
Donadelli V, Di Marco P, Mandich A, Finoia MG, Cardinaletti G, Petochi T, Longobardi A, Tibaldi E, Marino G. Effects of Dietary Plant Protein Replacement with Insect and Poultry By-Product Meals on the Liver Health and Serum Metabolites of Sea Bream (Sparus aurata) and Sea Bass (Dicentrarchus labrax). Animals. 2024; 14(2):241. https://doi.org/10.3390/ani14020241
Chicago/Turabian StyleDonadelli, Valeria, Patrizia Di Marco, Alberta Mandich, Maria Grazia Finoia, Gloriana Cardinaletti, Tommaso Petochi, Alessandro Longobardi, Emilio Tibaldi, and Giovanna Marino. 2024. "Effects of Dietary Plant Protein Replacement with Insect and Poultry By-Product Meals on the Liver Health and Serum Metabolites of Sea Bream (Sparus aurata) and Sea Bass (Dicentrarchus labrax)" Animals 14, no. 2: 241. https://doi.org/10.3390/ani14020241