
Tissue Lipid Profiles of Rainbow Trout, Oncorhynchus mykiss, Cultivated under Environmental Variables on a Diet Supplemented with Dihydroquercetin and Arabinogalactan

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Diet Preparation
2.2. Experimental Design and Fish Sampling
2.3. Biometrical Measurements and Survival
2.4. Analyses of Tissue Lipids
2.5. Analysis of Fatty Acids
2.6. Statistical Analyses
3. Results
3.1. Growth Performance and Mortality
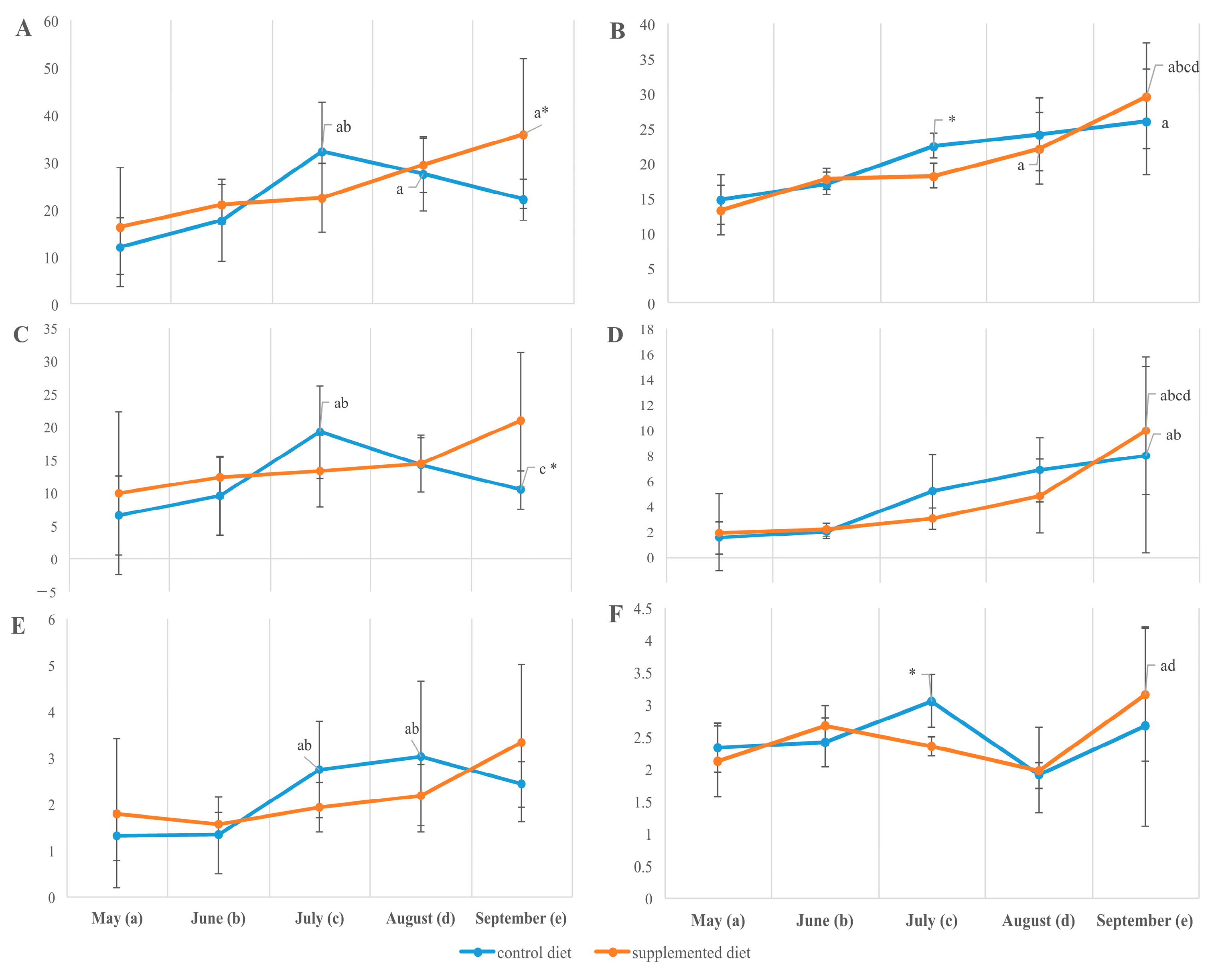
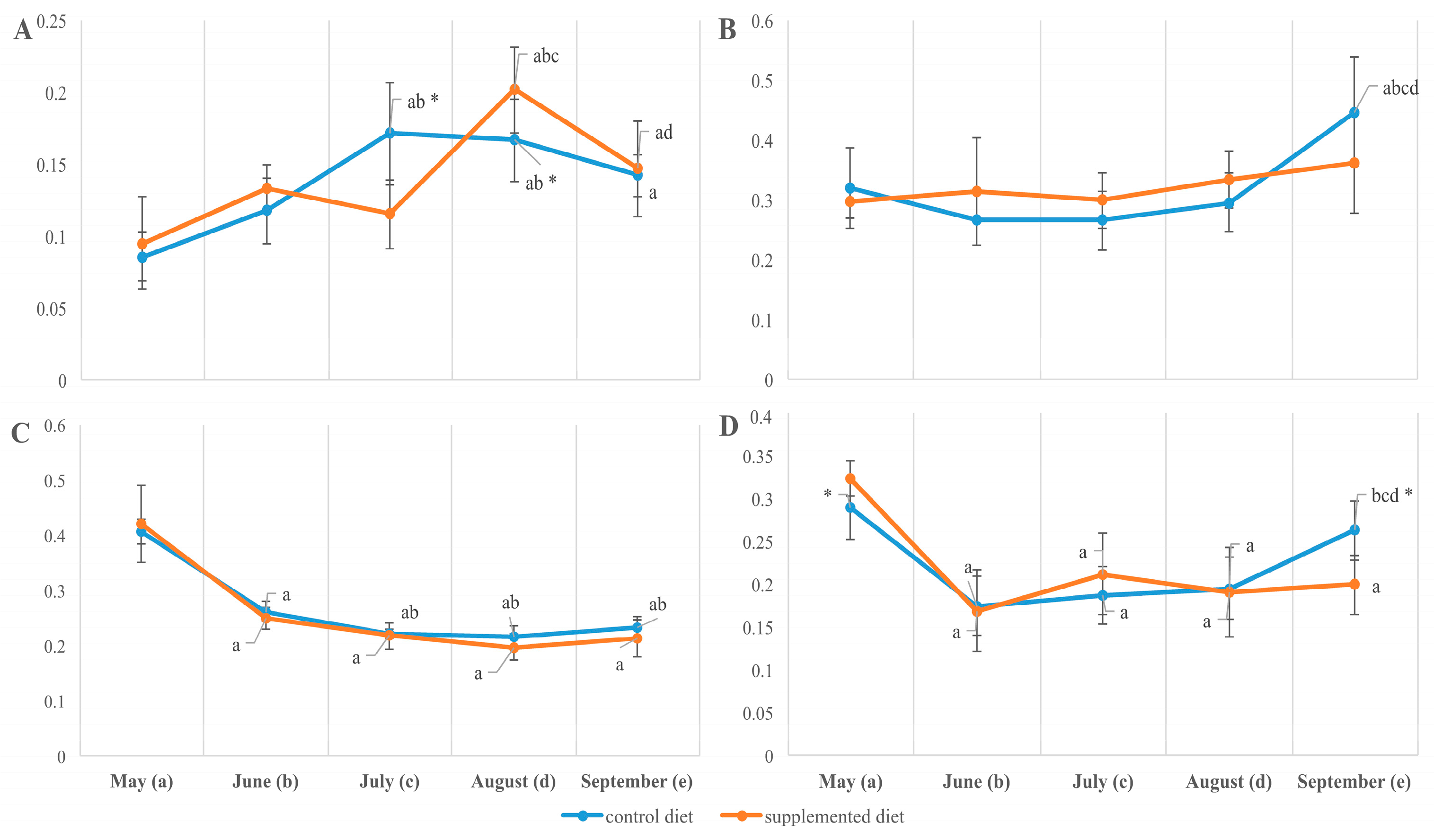
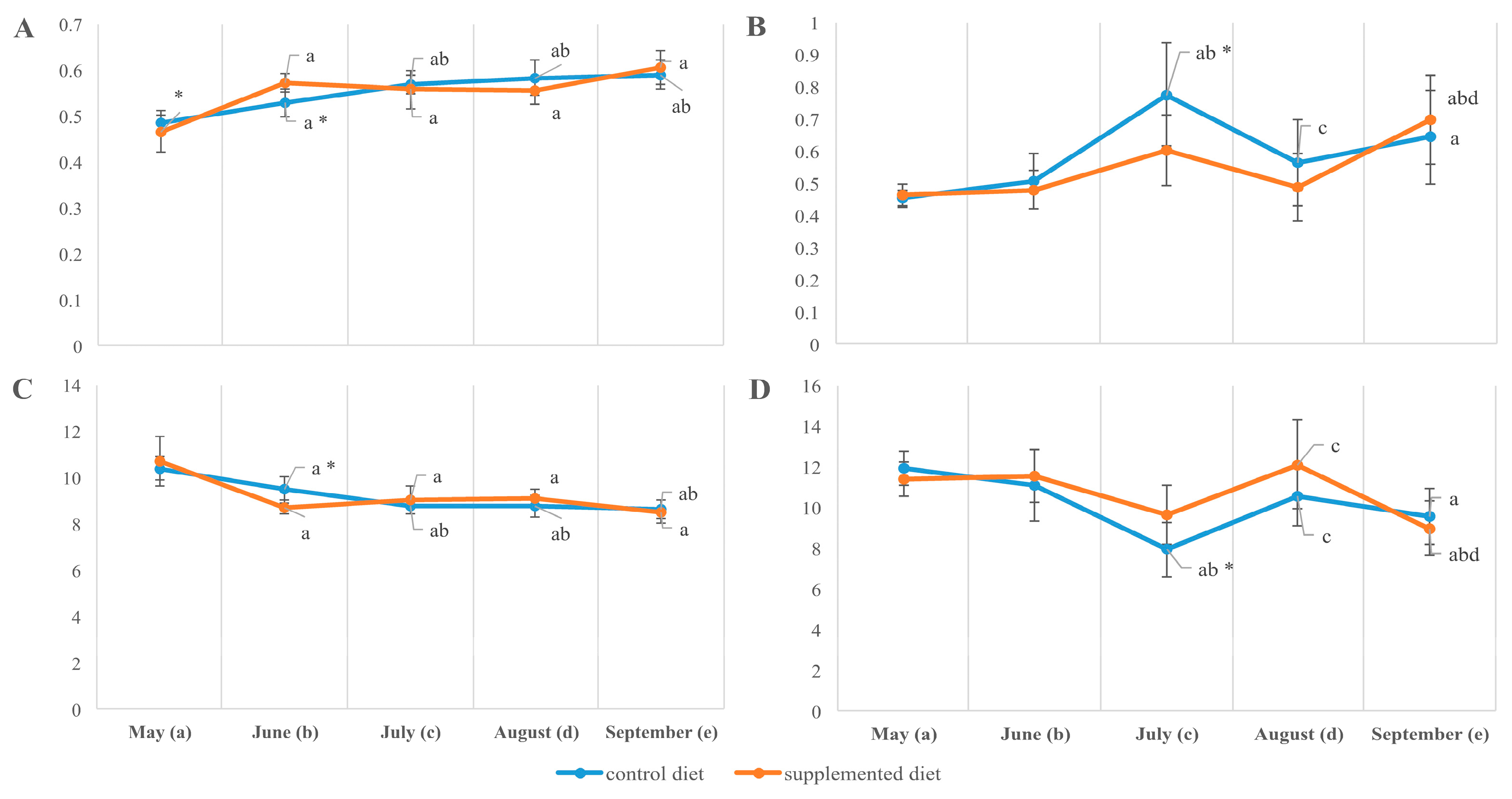
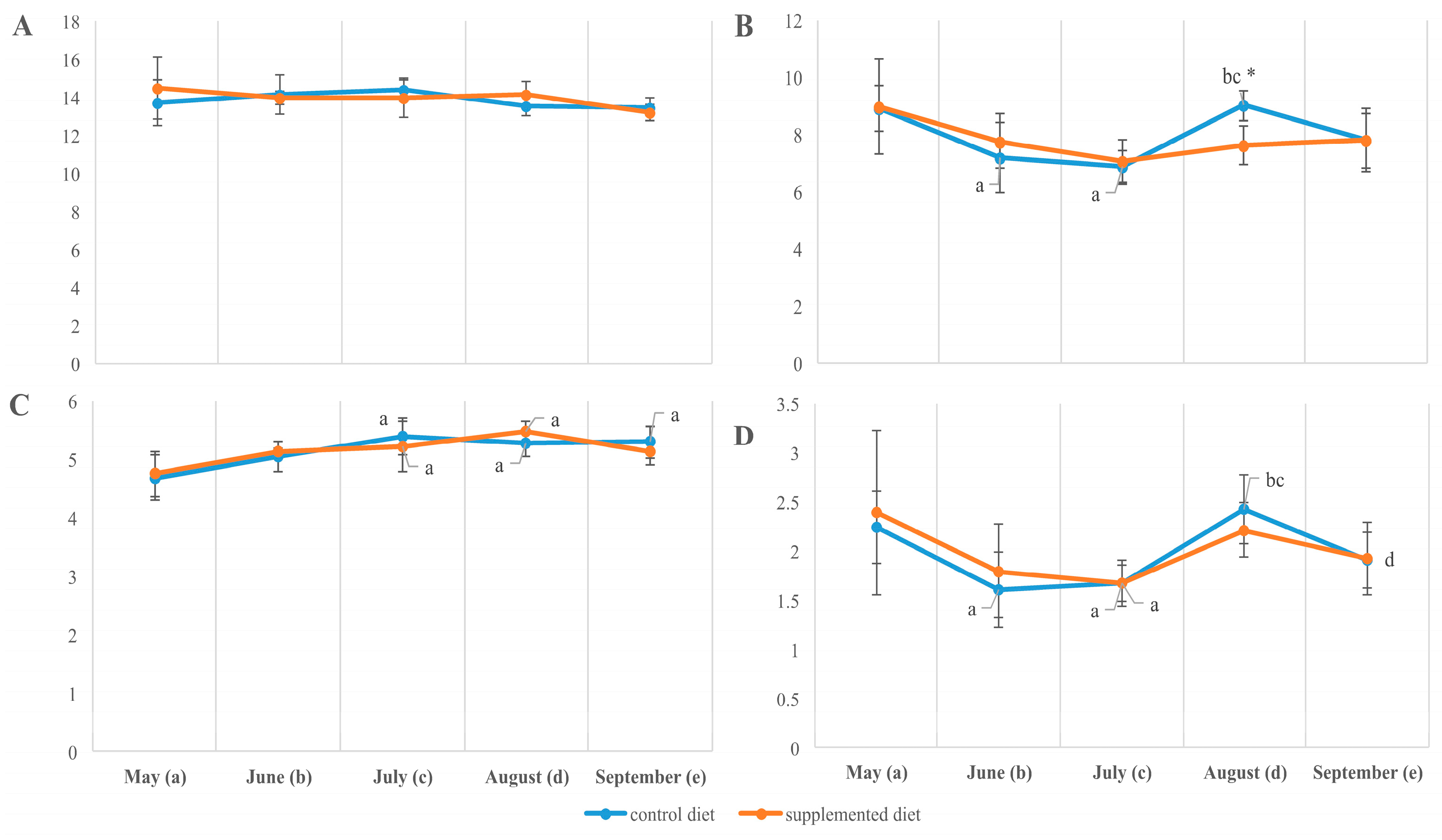
3.2. Tissue Lipid Composition
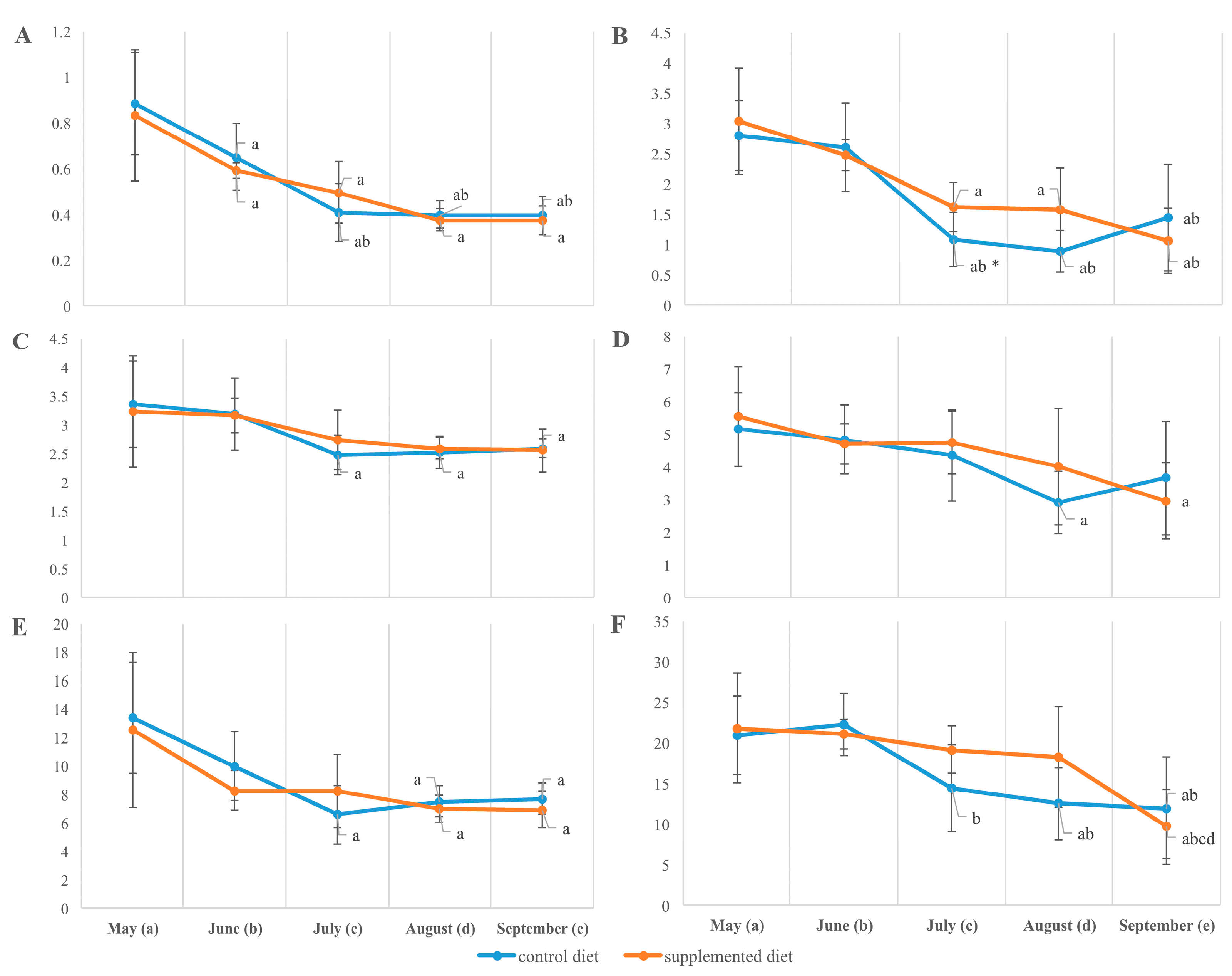
3.3. Tissue Fatty Acid Composition
4. Discussion
4.1. Summary of Growth Performance
4.2. Growth-Related Dynamics in Lipid Accumulation
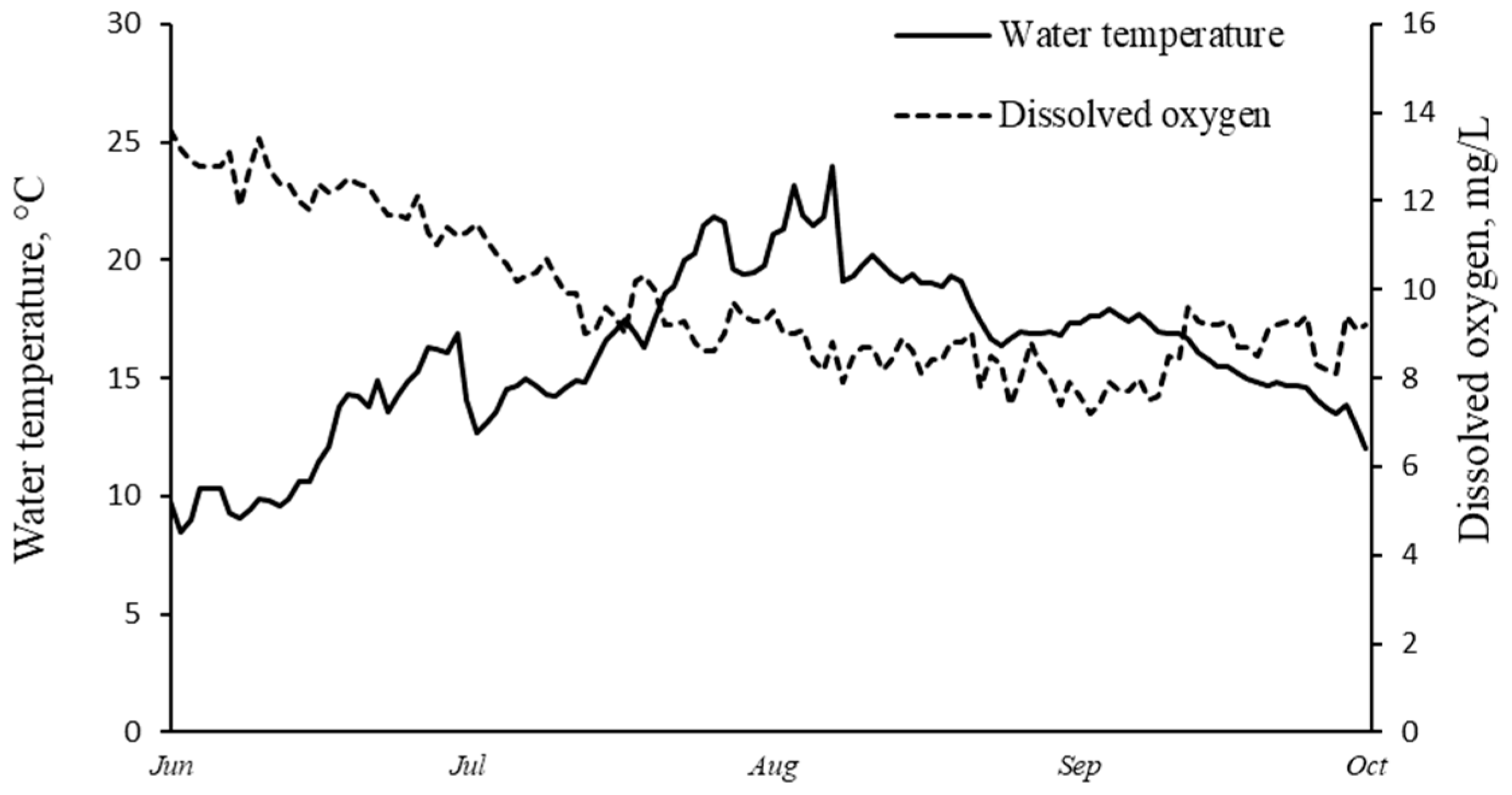
4.3. Temperature Response
4.4. Fillet Lipid and Fatty Acid Composition
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Salem, M.; Silverstein, J.; Rexroad, C.E., III; Yao, J. Effect of starvation on global gene expression and proteolysis in rainbow trout (Oncorhynchus mykiss). BMC Genom. 2007, 8, 328–343. [Google Scholar] [CrossRef] [PubMed]
- Maor, M.L.; Weber, G.M.; Salem, M.; Yao, J.; Aussanasuwannakul, A.; Kenney, P.B. Effect of sexual maturation and triploidy on chemical composition and fatty acid content of energy stores in female rainbow trout, Oncorhynchus mykiss. Aquaculture 2012, 364–365, 312–321. [Google Scholar] [CrossRef]
- Hazel, J.R.; Williams, E.E. The role of alterations in membrane lipid-composition in enabling physiological adaptation of organisms to their physical environment. Prog. Lipid Res. 1990, 29, 167–227. [Google Scholar] [CrossRef] [PubMed]
- Hochachka, P.W.; Somero, G.N. Biochemical Adaptation: Mechanism and Process in Physiological Evolution; Oxford University Press: Oxford, UK, 2002. [Google Scholar]
- Fokina, N.N.; Lysenko, L.A.; Ruokolainen, T.R.; Sukhovskaya, I.V.; Kantserova, N.P.; Nemova, N.N. Dependence of the lipid and unsaturated fatty-acid compositions in rainbow trout skeletal muscle on the rearing conditions and physiological status offish. Appl. Biochem. Microbiol. 2020, 56, 356–362. [Google Scholar] [CrossRef]
- Awad, E.; Awaad, A.S.; Esteban, M.A. Effects of dihydroquercetin obtained from deodar (Cedrus deodara) on immune status of gilthead sea bream (Sparus aurata L.). Fish Shellfish Immunol. 2015, 43, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Awad, E.; Awaad, A. Role of medicinal plants on growth performance and immune status infish. Fish Shellfish Immunol. 2017, 67, 40–54. [Google Scholar] [CrossRef] [PubMed]
- GarcíaBeltrán, J.M.; Esteban, M.Á. Nature-identical compounds as feed additives in aquaculture. Fish Shellfish Immunol. 2022, 123, 409–416. [Google Scholar] [CrossRef]
- Vijayaram, S.; Sun, Y.Z.; Zuorro, A.; Ghafarifarsani, H.; Van Doan, H.; Hoseinifar, S.H. Bioactive immunostimulants as health-promoting feed additives in aquaculture: A review. Fish Shellfish Immunol. 2022, 130, 294–308. [Google Scholar] [CrossRef]
- Maranesi, M.; Marchetti, M.; Bochicchio, D.; Cabrini, L. Vitamin B6 supplementation increases the docosahexaenoic acid concentration of muscle lipids of rainbowtrout (Oncorhynchus mykiss). Aquac. Res. 2005, 36, 431–438. [Google Scholar] [CrossRef]
- Acar, Ü.; Türker, A. Response of rainbow trout (Oncorhynchus mykiss) to unrefined peanut oil diets: Effect on growth performance, fish health and fillet fatty acid composition. Aquac. Nutr. 2018, 24, 292–299. [Google Scholar] [CrossRef]
- Vizcaíno, A.J.; Rodiles, A.; López, G.; Sáez, M.I.; Herrera, M.; Hachero, I.; Martínez, T.F.; Cerón-García, M.C.; Javier Alarcón, F. Growth performance, body composition, and digestive functionality of Senegalese sole (Solea senegalensis Kaup, 1858) juveniles fed diets including microalgae freeze-dried biomass. Fish Physiol. Biochem. 2018, 44, 661–677. [Google Scholar] [CrossRef] [PubMed]
- Ross, J.A.; Kasum, C.M. Dietary flavonoids: Bioavailability, metabolic effects, and safety. Annu. Rev. Nutr. 2002, 22, 19–34. [Google Scholar] [CrossRef]
- Awad, E.; Austin, D.; Lyndon, A.R. Effect of black cumin seed oil (Nigella sativa) and nettle extract (Quercetin) on enhancement of immunity in rainbow trout, Oncorhynchus mykiss (Walbaum). Aquaculture 2013, 388, 93–197. [Google Scholar] [CrossRef]
- Polyak, S.J.; Morishima, C.; Lohmann, V.; Pal, S.; Lee, D.Y.W.; Liu, Y.; Graf, T.N.; Oberlies, N.H. Identification of hepatoprotective flavonolignans from silymarin. Proc. Natl. Acad. Sci. USA 2010, 107, 5995–5999. [Google Scholar] [CrossRef] [PubMed]
- Wagoner, J.; Negash, A.; Kane, O.J.; Martinez, L.E.; Nahmias, Y.; Bourne, N.; Owen, D.M.; Grove, J.; Brimacombe, C.; McKeating, J.A. Multiple effects of silymarin on the hepatitis C virus life cycle. J. Hepatol. 2010, 51, 1912–1921. [Google Scholar] [CrossRef] [PubMed]
- Abid, N.; Hamad, E.; Ibrahim, M.; Abid, H.A. Antibacterial and antibiofilm activities of taxifolin against vancomycin-resistant S. aureus (VRSA). BaghdadJ. Biochem. Appl. Biol.Sci. 2022, 3, 262–272. [Google Scholar] [CrossRef]
- Choi, E.M.; Kim, A.J.; Kim, Y.O.; Hwang, J.K. Immunomodulating activity of arabinogalactan and fucoidan in vitro. J. Med. Food 2005, 8, 446–453. [Google Scholar] [CrossRef] [PubMed]
- Ramberg, J.E.; Nelson, E.D.; Sinnott, R.A. Immunomodulatory dietary polysaccharides: A systematic review of the literature. Nutr. J. 2010, 9, 54. [Google Scholar] [CrossRef]
- Udani, J.K. Immunomodulatory effects of ResistAid: A randomized, double-blind, placebo-controlled, multidose study. J. Am. Coll. Nutr. 2013, 32, 331–338. [Google Scholar] [CrossRef]
- Udani, J.K.; Singh, B.B.; Barrett, M.L.; Singh, V.J. Proprietary arabinogalactan extract increases antibody response to the pneumonia vaccine: A randomized, double-blind, placebo-controlled, pilot study in healthy volunteers. Nutr. J. 2010, 9, 32. [Google Scholar] [CrossRef]
- Das, A.; Baidya, R.; Chakraborty, T.; Samanta, A.K.; Roy, S. Pharmacological basis and new insights of taxifolin: A comprehensive review. Biomed. Pharmacother. 2021, 142, 112004. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Shi, X.; Tian, Y.; Zhai, S.; Liu, Y.; Xiong, Z.; Chu, S. An insight into novel therapeutic potentials of taxifolin. Front. Pharmacol. 2023, 14, 1173855. [Google Scholar] [CrossRef] [PubMed]
- Jain, S.; Vaidya, A. Comprehensive review on pharmacological effects and mechanism of actions of taxifolin: A bioactive flavonoid. Pharmacol. Res. Mod. Chin. Med. 2023, 7, 10024. [Google Scholar] [CrossRef]
- Manigandan, K.; Jayaraj, R.L.; Jagatheesh, K.; Elangovan, N. Taxifolin mitigates oxidative DNA damage in vitro and protects zebra fish (Danio rerio) embryos against cadmium toxicity. Environ. Toxicol. Pharmacol. 2015, 39, 1252–1261. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.H.; Wang, W.Y.; Chang, C.C.; Liou, K.T.; Sung, Y.J.; Liao, J.F.; Chen, C.F.; Chang, S.; Hou, Y.C.; Chou, Y.C.; et al. Taxifolin ameliorates cerebral ischemia-reperfusion injury in rats through its anti-oxidative effect and modulationof NF-kappa B activation. J. Biomed. Sci. 2006, 13, 127–141. [Google Scholar] [CrossRef] [PubMed]
- Ahiskali, I.; Pinar, C.L.; Kiki, M.; Mammadov, R.; OzbekBilgin, A.; Hacimuftuoglu, A.; Cankaya, M.; KeskinCimen, F.; Altuner, D. Effect of taxifolin on development of retinopathy in alloxan-induced diabetic rats. Cutan. Ocul. Toxicol. 2019, 38, 227–232. [Google Scholar] [CrossRef] [PubMed]
- Wan, F.; Han, H.; Zhong, R.; Wang, M.; Tang, S.; Zhang, S.; Hou, F.; Yi, B.; Zhang, H. Dihydroquercetin supplement alleviates colonic inflammation potentially through improved gut microbiota community in mice. Food Funct. 2021, 12, 11420–11434. [Google Scholar] [CrossRef] [PubMed]
- Theriault, A.; Wang, Q.; VanIderstine, S.C.; Chen, B.; Franke, A.A.; Adeli, K. Modulation of hepatic lipoprotein synthesis and secretion by taxifolin, a plant flavonoid. J. Lipid Res. 2000, 41, 1969–1979. [Google Scholar] [CrossRef]
- Casaschi, A.; Rubio, B.K.; Maiyoh, G.K.; Theriault, A.G. Inhibitory activity of diacylglycerol acyltransferase (DGAT) and microsomal triglyceride transfer protein (MTP) by the flavonoid, taxifolin, in HepG2cells: Potential role in the regulationof apolipoprotein B secretion. Atherosclerosis 2004, 176, 247–253. [Google Scholar] [CrossRef]
- Dion, C.; Chappuis, E.; Ripoll, C. Does larch arabinogalactan enhance immune function? A review of mechanistic and clinical trials. Nutr. Metab. 2016, 13, 28. [Google Scholar] [CrossRef]
- Lysenko, L.; Kantserova, N.; Parshukov, A.; Sukhovskaya, I. Dietary supply of the antioxidant and prebiotic mix promotes muscle growth and improves disease resistance in cultivated fish. Acta Physiol. 2019, 227, 45. [Google Scholar] [CrossRef]
- Kantserova, N.; Lysenko, L.; Churova, M.; Tushina, E.; Sukhovskaya, I.; Nemova, N. Dietary supplement with dihydroquercetin and arabinogalactan affects growth performance, intracellular protease activities and muscle-specific gene expression in bacterially infected Oncorhynchus mykiss. Int. Aquatic Res. 2020, 12, 63–73. [Google Scholar] [CrossRef]
- Sukhovskaya, I.V.; Lysenko, L.A.; Fokina, N.N.; Kantserova, N.P.; Borvinskaya, E.V. Survival, Growth Performance, and Hepatic Antioxidant and Lipid Profiles in Infected Rainbow Trout (Oncorhynchus mykiss) Fed a Diet Supplemented with Dihydroquercetin and Arabinogalactan. Animals 2023, 13, 1345. [Google Scholar] [CrossRef] [PubMed]
- Folch, J.; Lees, M.; Stanley, G.H.S. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef] [PubMed]
- Engelbrecht, F.M.; Mari, F.; Anderson, J.T. Cholesterol determination in serum: A rapid direction method. S. Afr. Med. J. 1974, 48, 250–256. [Google Scholar] [PubMed]
- Arduini, A.; Peschechera, A.; Dottori, S.; Sciarroni, A.F.; Serafini, F.; Calvani, M. High performance liquid chromatography of long-chain acylcarnitine and phospholipids in fatty acid turnover studies. J. Lipid Res. 1996, 37, 684–689. [Google Scholar] [CrossRef]
- Pirgozliev, V.; Westbrook, C.; Woods, S.; Karakeçili, M.R.; Karadaş, F.; Rose, S.P.; Mansbridge, S.C. Feeding dihydroquercetin to broiler chickens. Brit. Poultry Sci. 2019, 60, 241–245. [Google Scholar] [CrossRef] [PubMed]
- Kantserova, N.; Borvinskaya, E.; Lysenko, L.; Sukhovskaya, I.; Churova, M.; Tushina, E. Data set on body weight and length of rainbow trout, Oncorhynchus mykiss, fed with dihydroquercetin, arabinogalactan or a mixture of both in an aquaria experiment. Data Brief 2020, 32, 106184. [Google Scholar] [CrossRef]
- Zhai, S.W.; Liu, S.L. Effects of dietary quercetin on growth performance, serum lipids level and body composition of tilapia (Oreochromis niloticus). Ital. J. Anim. Sci. 2013, 12, e85. [Google Scholar] [CrossRef]
- Jia, E.; Yan, Y.; Zhou, M.; Li, X.; Jiang, G.; Liu, W.; Zhang, D. Combined effects of dietary quercetin and resveratrol on growth performance, antioxidant capability and innate immunity of blunt snout bream (Megalobrama amblycephala). Anim. Feed Sci. Technol. 2019, 256, 114268. [Google Scholar] [CrossRef]
- Ahmadifar, E.; Yousefi, M.; Karimi, M.; Raieni, R.F.; Dadar, M.; Yilmaz, S.; Dawood, M.A.O.; Abdel-Latif, H.M.R. Benefits of dietary polyphenols and polyphenol-rich additives to aquatic animal health: An overview. Rev. Fish Sci. Aquacult. 2021, 29, 478–511. [Google Scholar] [CrossRef]
- Krstić, M.P.; Mitrović, R.R.; Marković, R.V.; Ivanović, J.S.; Ćirković, M.A.; Djordjević, V.Z.; Baltić, M.Z. Fatty acid composition in the fillets of rainbow trout grown in different conditions. Eur. J. Lipid Sci. Technol. 2017, 119, 1600082. [Google Scholar] [CrossRef]
- Norambuena, F.; Morais, S.; Emery, J.A.; Turchini, G.M. Arachidonic acid and eicosapentaenoic acid metabolism in juvenile Atlantic salmon as affected by water temperature. PLoS ONE 2015, 10, e0143622. [Google Scholar] [CrossRef]
- Seiliez, I.; Panserat, S.; Kaushik, S.; Bergot, P. Cloning, tissue distribution and nutritional regulation of a Δ6-desaturase-like enzyme in rainbow trout. Comp. Biochem. Phys. B 2001, 130, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Tocher, D.R. Fatty acid requirements in ontogeny of marine and freshwater fish. Aquac. Res. 2010, 41, 717–732. [Google Scholar] [CrossRef]
- Martinez-Rubio, L.; Morais, S.; Evensen, Ø.; Wadsworth, S.; Vecino, J.G.; Ruohonen, K.; Tocher, D.R. Effect of functional feeds on fatty acid and eicosanoid metabolism in liver and head kidney of Atlantic salmon (Salmo salar L.) with experimentally induced heart and skeletal muscle inflammation. Fish Shellfish. Immunol. 2013, 34, 1533–1545. [Google Scholar] [CrossRef] [PubMed]
- Das, U.N. Arachidonic acid in health and disease with focus on hypertension and diabetes mellitus: A review. J. Adv. Res. 2018, 11, 43–55. [Google Scholar] [CrossRef] [PubMed]
- Caballero, M.J.; Obach, A.; Rosenlund, G.; Montero, D.; Gisvold, M.; Izquierdo, M.S. Impact of different dietary lipid sources on growth, lipid digestibility, tissue fatty acid composition and histology of rainbow trout, Oncorhynchus mykiss. Aquaculture 2002, 214, 253–271. [Google Scholar] [CrossRef]
- Roncarati, A.; Sirri, F.; Felici, A.; Stocchi, L.; Melotti, P.; Meluzzi, A. Effects of dietary supplementation with krill meal on pigmentation and quality of flesh of rainbow trout (Oncorhynchus mykiss). Ital. J. Anim. Sci. 2011, 10, e27. [Google Scholar] [CrossRef]
- Brenna, J.; Salem, N.; Sinclair, A.; Cunnane, S. α-Linolenic acid supplementation and conversion to n-3 long-chain polyunsaturated fatty acids in humans. Prostaglandins Leukot. Essent. Fat. Acids 2009, 80, 85–91. [Google Scholar] [CrossRef]
- Hixson, S.M.; Parrish, C.C.; Anderson, D.M. Full substitution of fish oil with camelina (Camelina sativa) oil, with partial substitution of fish meal with camelina meal, in diets for farmed Atlantic salmon (Salmo salar) and its effect on tissue lipids and sensory quality. Food Chem. 2014, 157, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Cleveland, B.M.; Gao, G.; Radler, L.M.; Picklo, M.J. Hepatic fatty acid and transcriptome profiles during the transition from vegetable- to fish oil-based diets in rainbow trout (Oncorhynchus mykiss). Lipids 2021, 56, 189–200. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pellet Size | 6 mm | 8 mm |
|---|---|---|
| Total lipids, % dry weight | 23.28 ± 0.37 | 24.03 ± 0.53 |
| Main lipid classes, % total lipid dry weight | ||
| Phospholipids | 0.19 ± 0.17 | 0.16 ± 0.24 |
| Triacylglycerols | 18.68 ± 0.45 | 18.55 ± 0.94 |
| Cholesterols | 4.60 ± 0.40 | 5.32 ± 0.63 |
| Fatty acid compositions, % sum fatty acids | ||
| 14:0 | 2.48 ± 0.04 | 2.35 ± 0.04 |
| 16:0 | 9.65 ± 0.15 | 8.73 ± 0.24 |
| 16:1n-7 | 2.22 ± 0.04 | 2.15 ± 0.02 |
| 18:0 | 2.61 ± 0.13 | 2.44 ± 0.23 |
| 18:1n-9cis | 41.15 ± 0.18 | 39.72 ± 0.18 |
| 18:2n-6cis | 16.15 ± 0.09 | 16.15 ± 0.10 |
| 18:3n-3 | 8.49 ± 0.06 | 8.52 ± 0.05 |
| 20:1n-9 | 2.16 ± 0.12 | 3.38 ± 0.06 |
| 20:5n-3 | 4.99 ± 0.08 | 4.25 ± 0.03 |
| 22:1n-9 | 2.40 ± 0.44 | 3.78 ± 0.41 |
| 22:6n-3 | 4.24 ± 0.08 | 5.63 ± 0.06 |
| Variables | Standard Diet | Supplemented Diet |
|---|---|---|
| Mortality (%) | 3.65 ± 0.33 | 2.88 ± 0.32 * |
| Initial weight (g) | 733.0 ± 58.3 | 770.1 ± 56.5 |
| Final weight (g) | 1947.3 ± 314.7 | 2289.3 ± 120.3 |
| Relative growth rate (%) | 286.4 ± 42.8 | 313.2 ± 67.7 |
| Specific growth rate (% day−1) | 1.47 ± 0.27 | 1.60 ± 0.46 |
| Feed conversion ratio | 1.27 ± 0.08 | 1.31 ± 0.06 |
| Variables | Muscle | Liver | ||
|---|---|---|---|---|
| Factor 1 | Factor 2 | Factor 1 | Factor 2 | |
| Total lipids | 0.42 | 0.89 | 0.96 | 0.21 |
| Phospholipids, PLs | 0.90 | 0.42 | 0.94 | 0.01 |
| Triacylglycerols, TAGs | 0.20 | 0.93 | 0.87 | 0.29 |
| Cholesterols, Chol | 0.27 | 0.85 | 0.70 | 0.20 |
| Phosphatidylserine, PS | 0.89 | 0.31 | 0.35 | 0.80 |
| Phosphatidylethanolamine, PEA | 0.90 | 0.01 | 0.22 | 0.92 |
| Phosphatidylcholine, PC | 0.86 | 0.49 | 0.92 | −0.26 |
| PEA/PC ratio | −0.29 | −0.61 | −0.52 | 0.83 |
| Expl.Var | 3.50 | 3.26 | 4.31 | 2.40 |
| Prp.Totl | 0.44 | 0.41 | 0.54 | 0.30 |
| Fatty acid composition | ||||
| Factor 1 | Factor 2 | Factor 1 | Factor 2 | |
| 16:0, palmitic | 0.44 | −0.84 | 0.47 | −0.71 |
| 18:0, stearic | 0.38 | −0.66 | 0.44 | −0.72 |
| Total saturate, SFA | 0.21 | −0.92 | 0.52 | −0.81 |
| 16:1n-7, palmitoleic | −0.91 | −0.01 | −0.88 | −0.32 |
| 18:1n-9, oleic | −0.85 | 0.50 | −0.94 | 0.27 |
| 18:1n-7, vaccenic | −0.37 | 0.29 | −0.67 | 0.29 |
| Total monoenes, MUFA | −0.91 | 0.38 | −0.97 | 0.21 |
| 18:2n-6, linoleic, LA | −0.13 | 0.93 | −0.17 | 0.82 |
| 18:3n-6, γ-linolenic | 0.39 | 0.22 | 0.04 | 0.02 |
| 20:4n-6, arachidonic, ArA | 0.92 | −0.25 | 0.92 | 0.04 |
| Total n-6 PUFA | 0.20 | 0.88 | 0.57 | 0.71 |
| 18:3n-3, α-linolenic, ALA | −0.69 | 0.43 | −0.13 | 0.74 |
| 20:5n-3, eicosapentaenoic, EPA | 0.82 | −0.40 | 0.90 | −0.22 |
| 22:6n-3, docosahexaenoic, DHA | 0.91 | −0.38 | 0.98 | −0.12 |
| Total n-3 PUFA | 0.92 | −0.38 | 0.99 | −0.08 |
| Total PUFA | 0.98 | −0.16 | 0.99 | 0.06 |
| n-3/n-6 PUFA ratio | 0.81 | −0.55 | 0.87 | −0.38 |
| SFA/PUFA ratio | −0.88 | −0.44 | −0.70 | −0.66 |
| 16:1n-7/16:0 ratio | −0.84 | 0.38 | −0.94 | −0.04 |
| 18:1n-9/18:0 ratio | −0.68 | 0.66 | −0.77 | 0.49 |
| 18:1n-7/16:1n-7 ratio | 0.79 | 0.10 | 0.80 | 0.37 |
| Unsaturation index | 0.92 | 0.26 | 0.51 | 0.78 |
| Expl.Var | 11.78 | 6.03 | 12.30 | 5.35 |
| Prp.Totl | 0.54 | 0.27 | 0.56 | 0.24 |
| Variables | Muscle | Liver | ||||
|---|---|---|---|---|---|---|
| Sampling Date | Fish Weight | Dietary Supplement | Sampling Date | Fish Weight | Dietary Supplement | |
| Total lipids | 0.50 | 0.53 | −0.13 | 0.65 | 0.68 | 0.03 |
| Phospholipids, PLs | 0.60 | 0.57 | −0.04 | 0.62 | 0.63 | 0.06 |
| Triacylglycerols, TAGs | 0.34 | 0.40 | −0.15 | 0.60 | 0.64 | 0.02 |
| Cholesterols, Chol | 0.45 | 0.48 | −0.02 | 0.16 | 0.25 | 0.02 |
| Phosphatidylserine, PS | 0.56 | 0.52 | −0.01 | 0.41 | 0.36 | −0.01 |
| Phosphatidylethanolamine, PEA | 0.19 | 0.20 | 0.05 | 0.20 | 0.18 | 0.10 |
| Phosphatidylcholine, PC | 0.64 | 0.61 | −0.05 | 0.56 | 0.58 | 0.05 |
| PEA/PC ratio | −0.75 | −0.68 | 0.04 | −0.31 | −0.34 | 0.00 |
| 16:0, palmitic | −0.19 | −0.16 | 0.14 | −0.37 | −0.44 | 0.01 |
| 18:0, stearic | −0.23 | −0.18 | 0.11 | −0.15 | −0.16 | 0.02 |
| Total saturate, SFA | −0.03 | −0.01 | 0.10 | −0.30 | −0.36 | 0.02 |
| 16:1n-7, palmitoleic | 0.59 | 0.64 | −0.08 | 0.61 | 0.64 | 0.08 |
| 18:1n-9, oleic | 0.51 | 0.51 | −0.04 | 0.53 | 0.57 | 0.10 |
| 18:1n-7, vaccenic | 0.29 | 0.27 | −0.02 | 0.76 | 0.74 | −0.01 |
| Total monoenes, MUFA | 0.63 | 0.61 | −0.06 | 0.63 | 0.67 | 0.08 |
| 18:2n-6, linoleic, LA | −0.27 | −0.25 | −0.06 | −0.18 | −0.14 | 0.03 |
| 18:3n-6, γ-linolenic | −0.35 | −0.42 | 0.16 | −0.19 | −0.21 | 0.18 |
| 20:4n-6, arachidonic, ArA | −0.74 | −0.70 | 0.03 | −0.69 | −0.69 | −0.08 |
| Total n-6 PUFA | −0.57 | −0.54 | −0.04 | −0.61 | −0.58 | −0.07 |
| 18:3n-3, α-linolenic, ALA | 0.50 | 0.52 | −0.01 | −0.07 | −0.02 | −0.05 |
| 20:5n-3, eicosapentaenoic, EPA | −0.49 | −0.45 | −0.01 | −0.52 | −0.55 | −0.06 |
| 22:6n-3, docosahexaenoic, DHA | −0.56 | −0.57 | 0.06 | −0.61 | −0.64 | −0.10 |
| Total n-3 PUFA | −0.56 | −0.55 | 0.05 | −0.61 | −0.64 | −0.10 |
| Total PUFA | −0.71 | −0.70 | 0.04 | −0.66 | −0.68 | −0.11 |
| n-3/n-6 PUFA ratio | −0.39 | −0.39 | 0.06 | −0.44 | −0.48 | −0.09 |
| SFA/PUFA ratio | 0.73 | 0.73 | 0.00 | 0.45 | 0.45 | 0.13 |
| 16:1n-7/16:0 ratio | 0.49 | 0.53 | −0.12 | 0.66 | 0.70 | 0.04 |
| 18:1n-9/18:0 ratio | 0.40 | 0.38 | −0.08 | 0.42 | 0.45 | 0.08 |
| 18:1n-7/16:1n-7 ratio | −0.47 | −0.52 | 0.06 | −0.52 | −0.51 | −0.16 |
| Unsaturation index | −0.67 | −0.69 | −0.02 | −0.37 | −0.32 | −0.12 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fokina, N.N.; Sukhovskaya, I.V.; Kantserova, N.P.; Lysenko, L.A. Tissue Lipid Profiles of Rainbow Trout, Oncorhynchus mykiss, Cultivated under Environmental Variables on a Diet Supplemented with Dihydroquercetin and Arabinogalactan. Animals 2024, 14, 94. https://doi.org/10.3390/ani14010094
Fokina NN, Sukhovskaya IV, Kantserova NP, Lysenko LA. Tissue Lipid Profiles of Rainbow Trout, Oncorhynchus mykiss, Cultivated under Environmental Variables on a Diet Supplemented with Dihydroquercetin and Arabinogalactan. Animals. 2024; 14(1):94. https://doi.org/10.3390/ani14010094
Chicago/Turabian StyleFokina, Natalia N., Irina V. Sukhovskaya, Nadezhda P. Kantserova, and Liudmila A. Lysenko. 2024. "Tissue Lipid Profiles of Rainbow Trout, Oncorhynchus mykiss, Cultivated under Environmental Variables on a Diet Supplemented with Dihydroquercetin and Arabinogalactan" Animals 14, no. 1: 94. https://doi.org/10.3390/ani14010094