
Genome-Wide Analysis of the KLF Gene Family in Chicken: Characterization and Expression Profile
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Genome-Wide Identification of KLF Family Members in Chickens
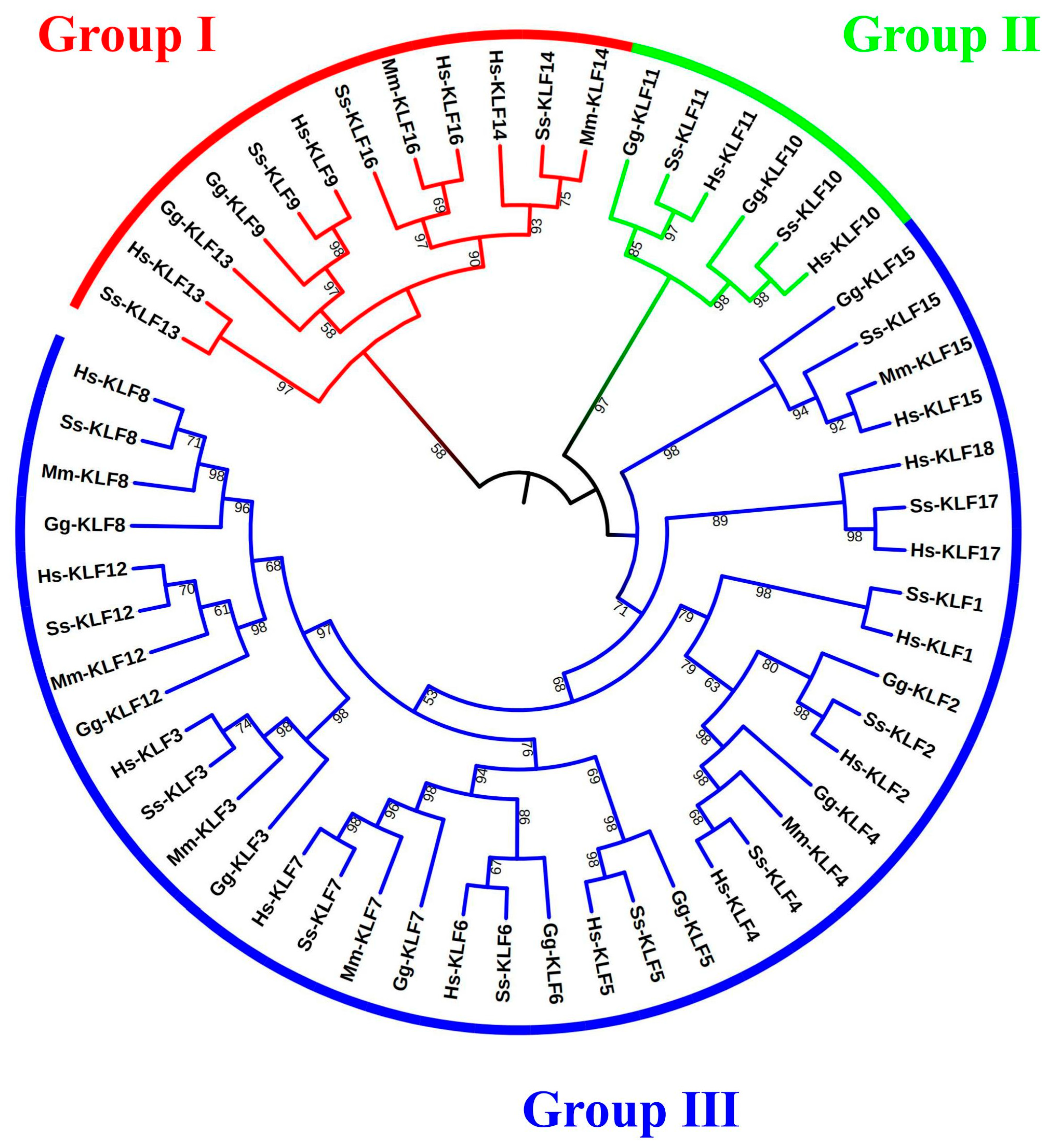
2.3. Protein Alignment, Phylogenetic Analysis, and Chromosome Location Analysis of KLF Genes
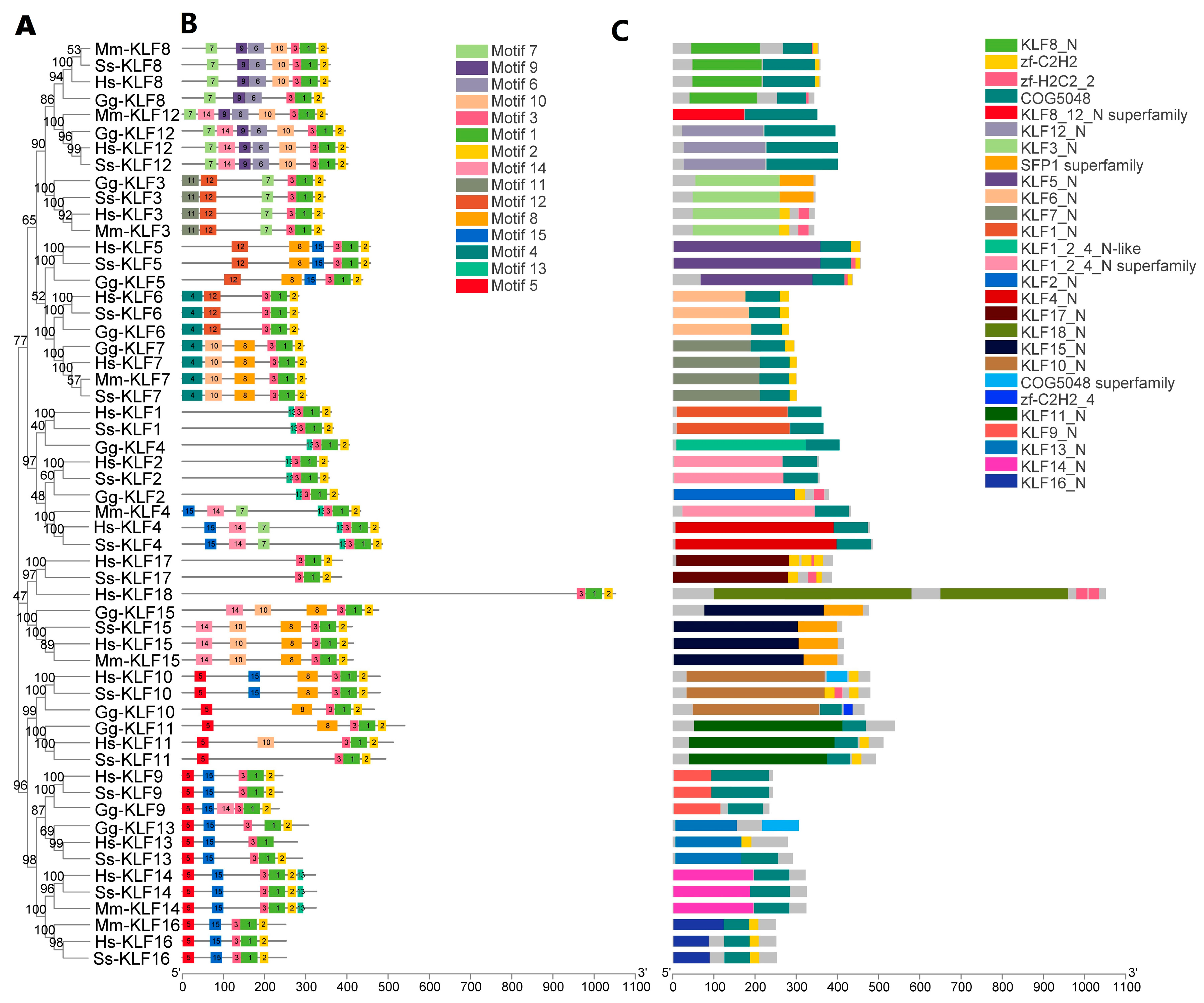
2.4. Motif Identification and Conserved Domain Analysis of KLF Genes
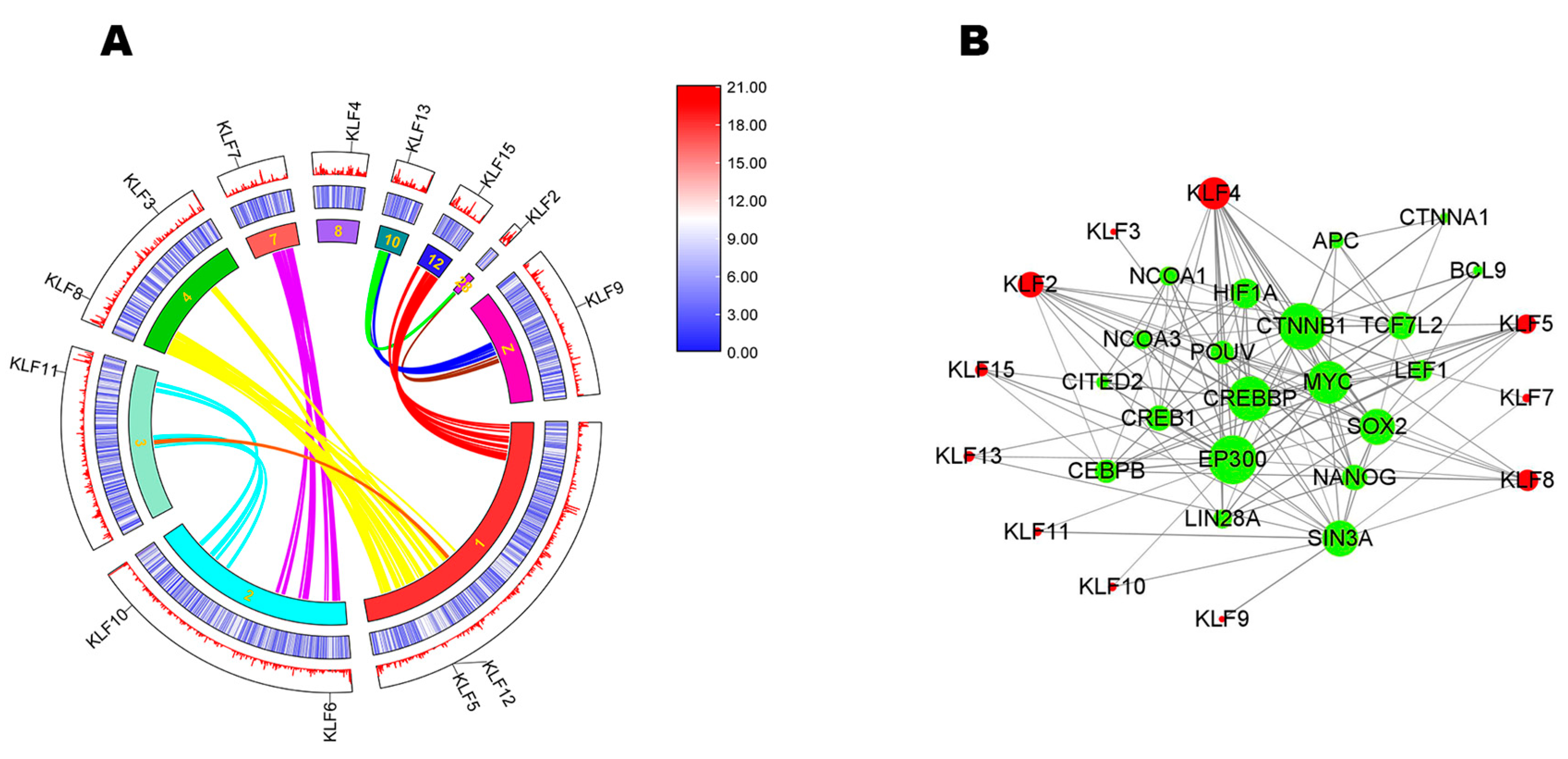
2.5. Synteny Analysis and Interaction Network Analysis of KLF Genes in Chickens
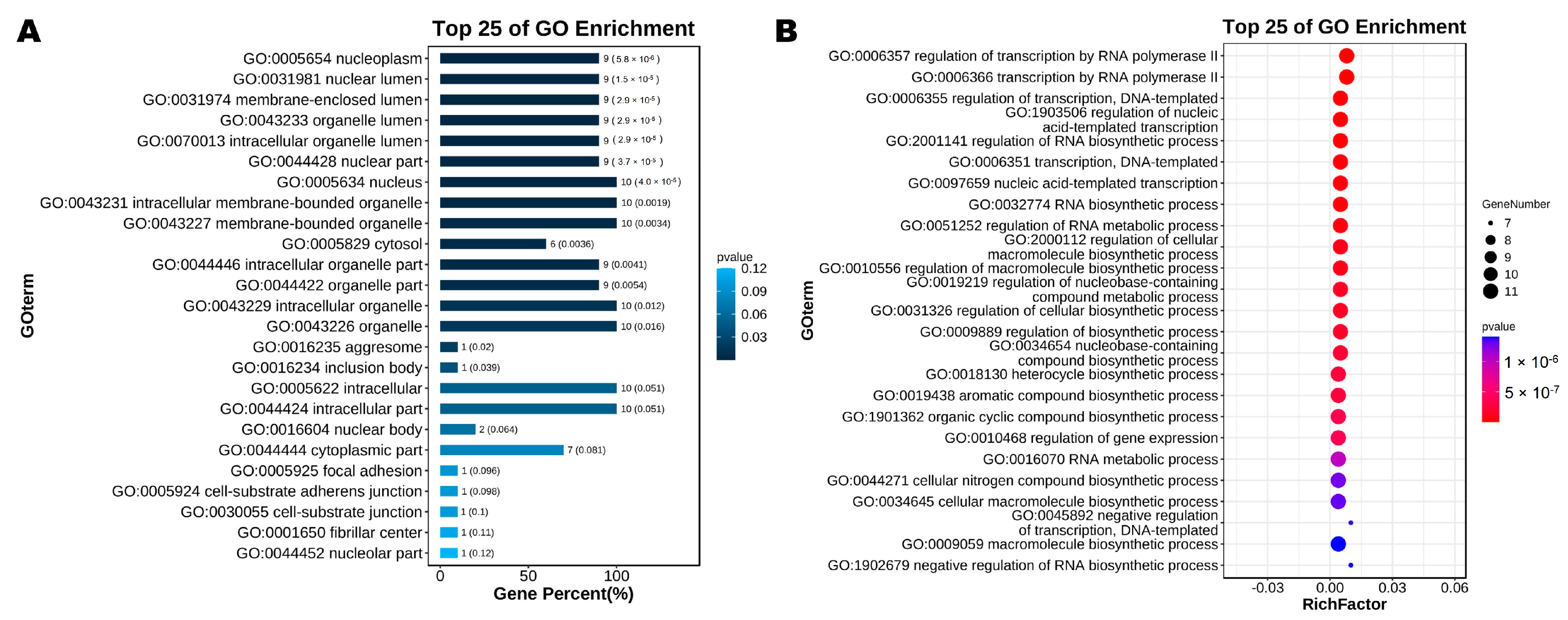
2.6. GO Enrichment Analysis and KEGG Pathway Analysis of KLF Genes in Chicken
2.7. RNA-seq Results Analysis of the G. gallus KLF Genes
2.8. Animals, Tissue Collections, RNA Extraction, and Quantitative Real-Time PCR (qPCR)
2.9. Statistics Analysis
3. Results
3.1. Identification and Phylogenetic Analysis of KLF Genes
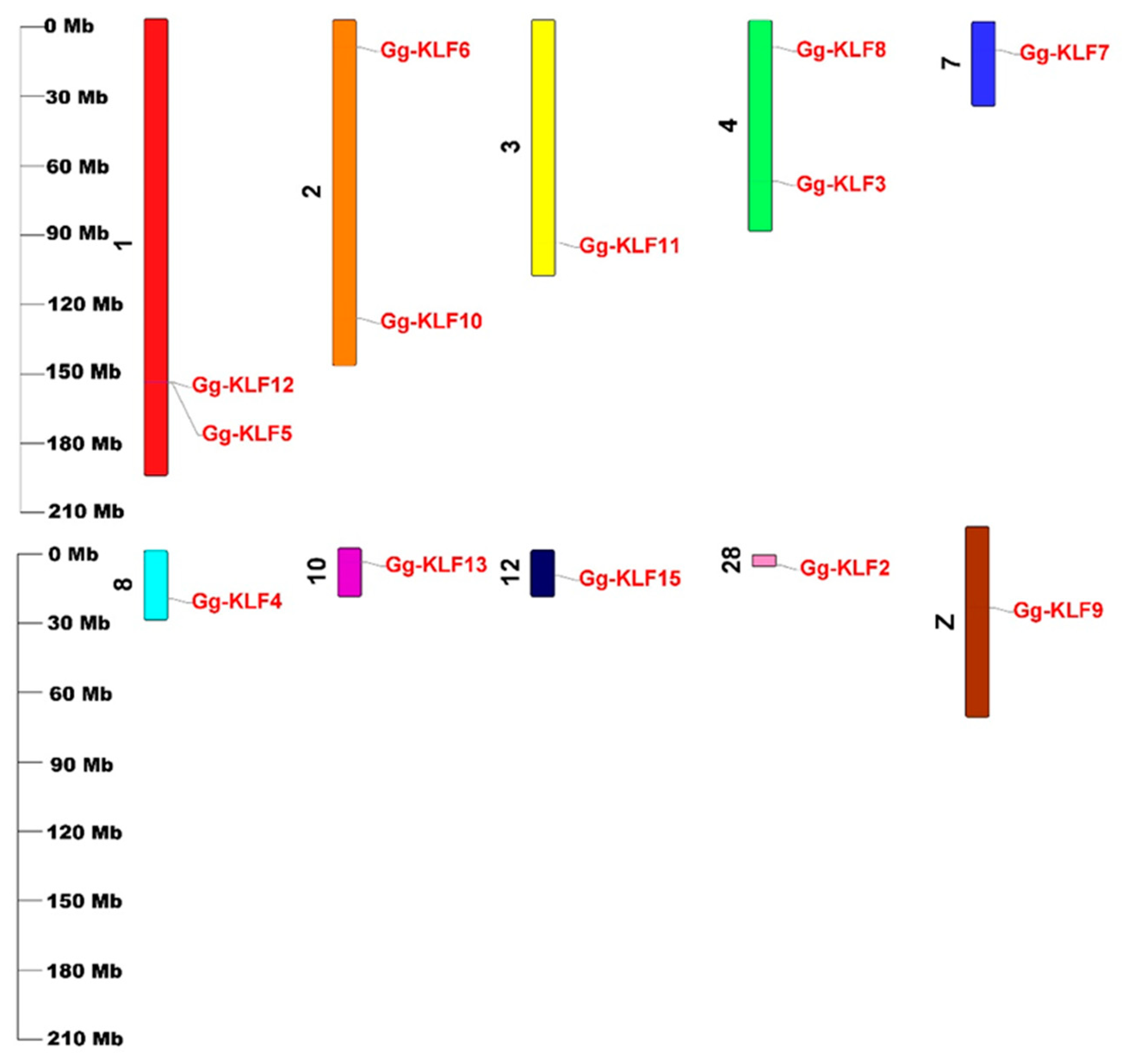
3.2. Chromosomal Locations of G. gallus KLF Genes
3.3. Motif Identification and Conserved Domain Analysis
3.4. Synteny Analysis and Interaction Network Analysis
3.5. GO Enrichment Analysis and KEGG Pathway Analysis of KLF Genes in Chicken
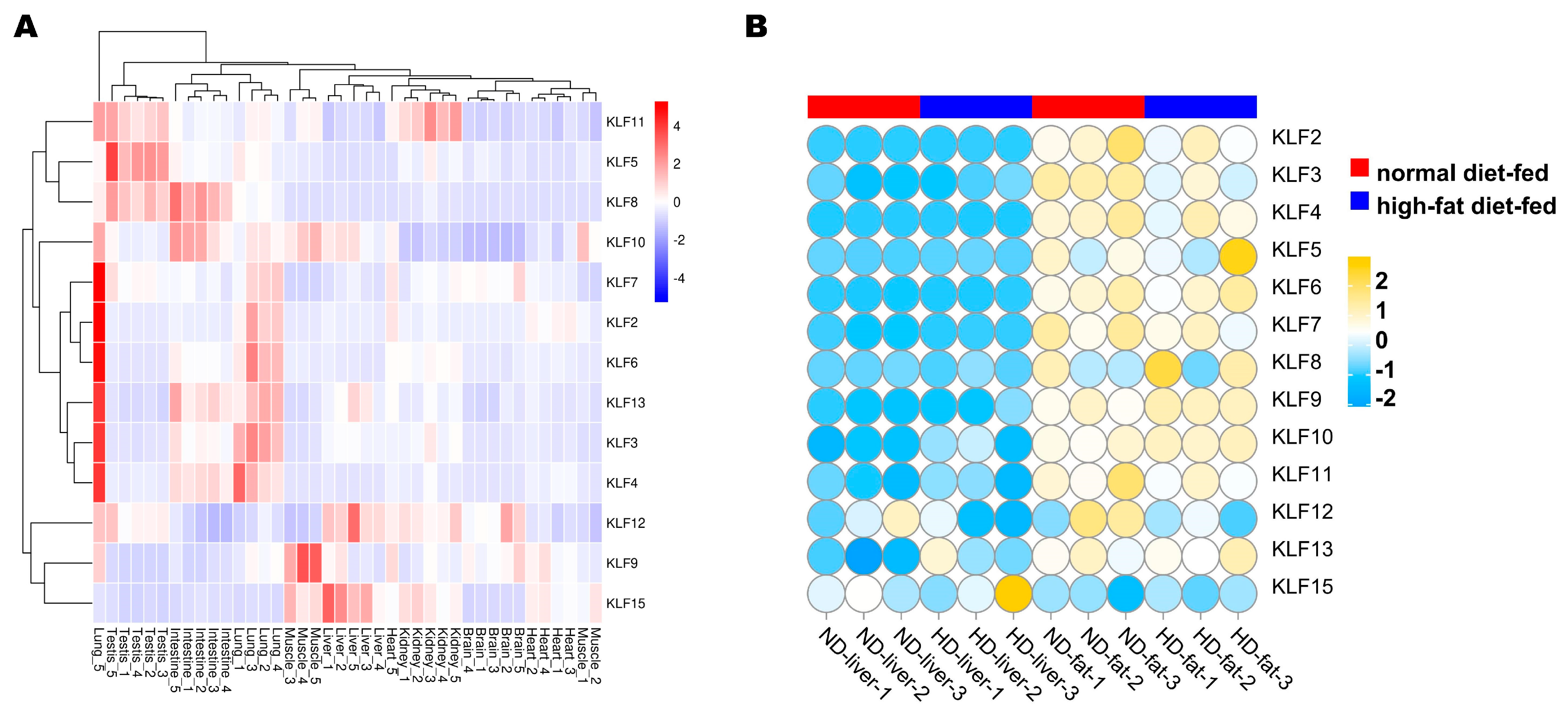
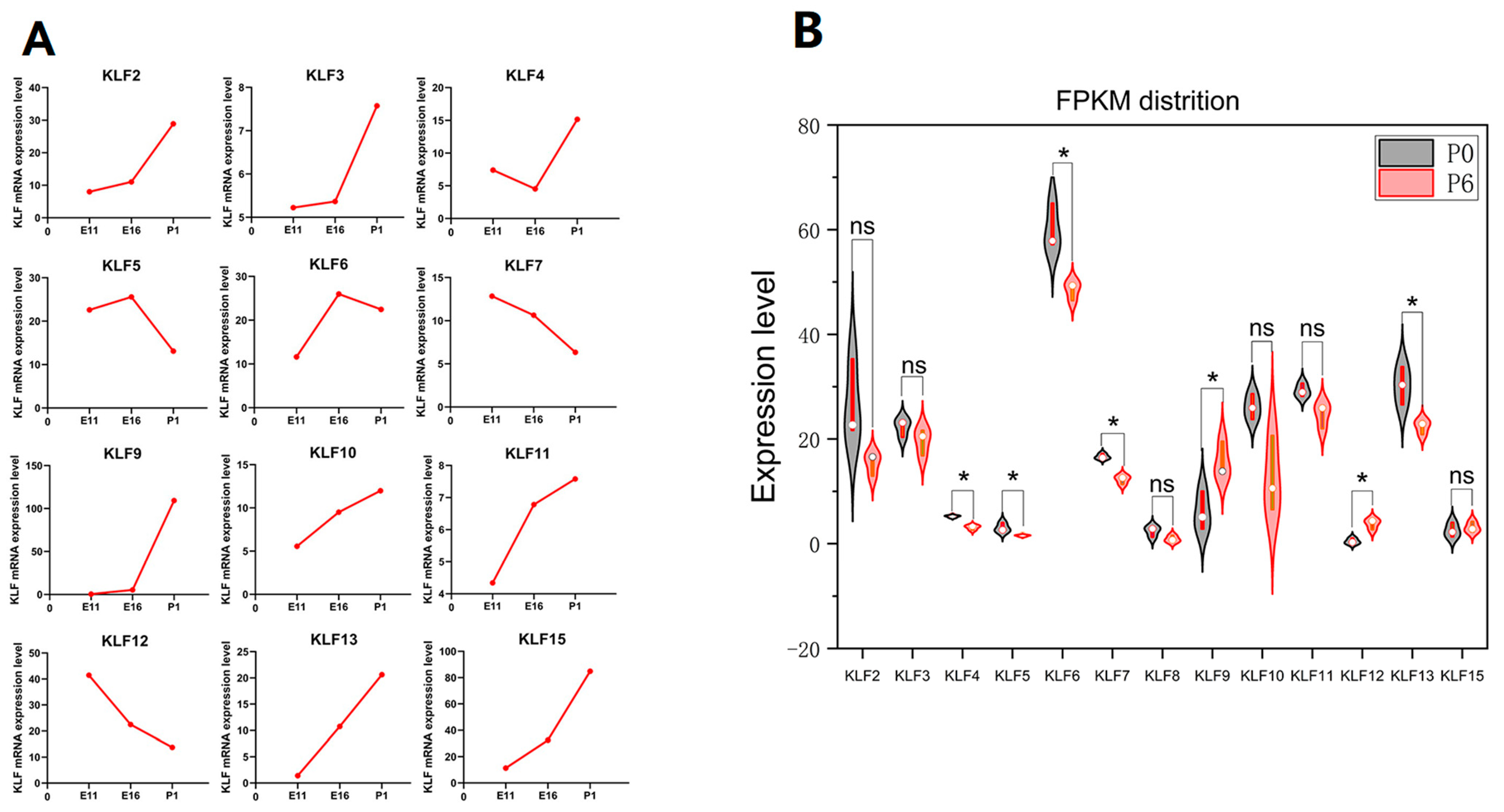
3.6. Expression Profile Analysis of G. gallus KLF Genes
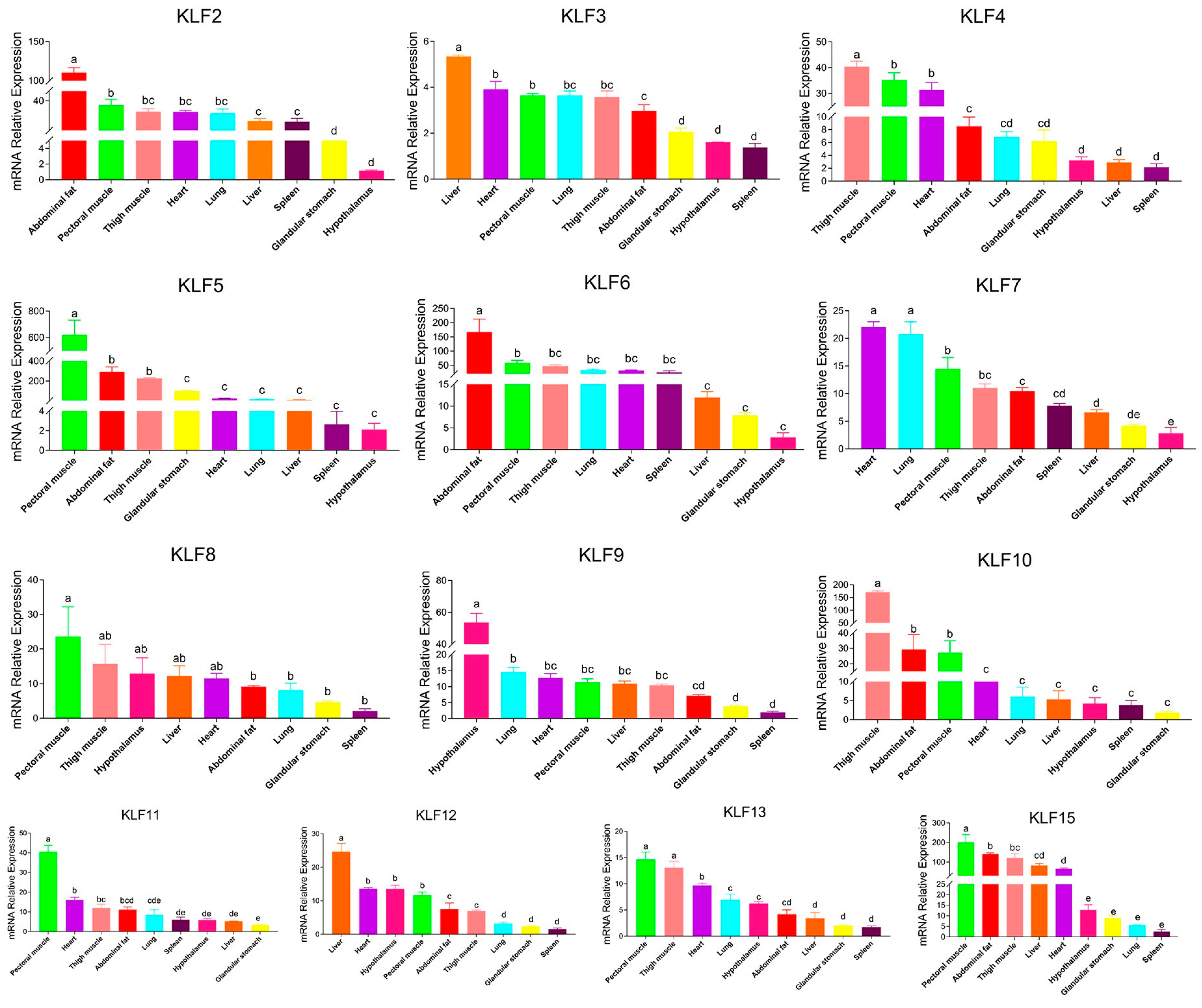
3.7. QPCR Results of the G. gallus KLF Genes
3.8. RNA-seq Result Analysis of the G. gallus KLF Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Katemala, S.; Molee, A.; Thumanu, K.; Yongsawatdigul, J. Meat quality and Raman spectroscopic characterization of Korat hybrid chicken obtained from various rearing periods. Poult. Sci. 2021, 100, 1248–1261. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Chang, Y.; Wu, G.; Zhang, W.; Man, C. Potential role of miR-155-5p in fat deposition and skeletal muscle development of chicken. Biosci. Rep. 2020, 40, BSR20193796. [Google Scholar] [CrossRef] [PubMed]
- Tian, W.; Wang, Z.; Wang, D.; Zhi, Y.; Dong, J.; Jiang, R.; Han, R.; Li, Z.; Kang, X.; Li, H.; et al. Chromatin Interaction Responds to Breast Muscle Development and Intramuscular Fat Deposition Between Chinese Indigenous Chicken and Fast-Growing Broiler. Front. Cell. Dev. Biol. 2021, 9, 782268. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.; Wei, Y.; You, G.; Liu, W.; Amevor, F.K.; Zhang, Y.; He, H.; Ma, M.; Zhang, Y.; Li, D.; et al. Circular PPP1R13B RNA Promotes Chicken Skeletal Muscle Satellite Cell Proliferation and Differentiation via Targeting miR-9-5p. Animals 2021, 11, 2396. [Google Scholar] [CrossRef]
- Cheng, J.; Wang, L.; Wang, S.; Chen, R.; Zhang, T.; Ma, H.; Lu, H.; Yuan, G. Transcriptomic analysis of thigh muscle of Lueyang black-bone chicken in free-range and caged feeding. Anim. Biotechnol. 2021, 1–11. [Google Scholar] [CrossRef]
- Zhu, M.; Wang, M.; Shao, Y.; Nan, Y.; Blair, H.T.; Morris, S.T.; Zhao, Z.; Zhang, H. Characterization of muscle development and gene expression in early embryos of chicken, quail, and their hybrids. Gene 2021, 768, 145319. [Google Scholar] [CrossRef]
- Shen, X.; Cui, C.; Tang, S.; Han, S.; Zhang, Y.; Xia, L.; Tan, B.; Ma, M.; Kang, H.; Yu, J.; et al. MyoG-enhanced circGPD2 regulates chicken skeletal muscle development by targeting miR-203a. Int. J. Biol. Macromol. 2022, 222, 2212–2224. [Google Scholar] [CrossRef]
- Parakati, R.; DiMario, J.X. Repression of myoblast proliferation and fibroblast growth factor receptor 1 promoter activity by KLF10 protein. J. Biol. Chem. 2013, 288, 13876–13884. [Google Scholar] [CrossRef]
- Ferreira, K.K.S.; de Morais Gomes, E.R.; de Lima Filho, J.L.; Castelletti, C.H.M.; Martins, D.B.G. Bioinformatics analysis of non-synonymous variants in the KLF genes related to cardiac diseases. Gene 2018, 650, 68–76. [Google Scholar] [CrossRef]
- Cantú, I.; van de Werken, H.J.G.; Gillemans, N.; Stadhouders, R.; Heshusius, S.; Maas, A.; Esteghamat, F.; Ozgur, Z.; van IJcken, W.F.J.; Grosveld, F.; et al. The mouse KLF1 Nan variant impairs nuclear condensation and erythroid maturation. PLoS ONE 2019, 14, e0208659. [Google Scholar] [CrossRef]
- Shi, G.; Yang, F. Krüppel-like factor 1 (KLF1) promoted the proliferation, migration and invasion of human lens epithelial cells by enhancing the expression of Zinc Finger and BTB Domain Containing 7A (ZBTB7A) and activating Wnt/β-catenin pathway. Bioengineered 2021, 12, 4374–4384. [Google Scholar] [CrossRef] [PubMed]
- Ayuso, M.; Fernández, A.; Núñez, Y.; Benítez, R.; Isabel, B.; Fernández, A.I.; Rey, A.I.; González-Bulnes, A.; Medrano, J.F.; Cánovas, Á.; et al. Developmental Stage, Muscle and Genetic Type Modify Muscle Transcriptome in Pigs: Effects on Gene Expression and Regulatory Factors Involved in Growth and Metabolism. PLoS ONE 2016, 11, e0167858. [Google Scholar] [CrossRef] [PubMed]
- Song, C.; Fang, X.; Yang, Z.; Wang, Q.; Meng, F.; Chen, Y.; Chen, J.; Zhao, B.; Wang, Y.; Fang, X.; et al. miR-152 Regulates Bovine Myoblast Proliferation by Targeting KLF6. Animals 2021, 11, 3001. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, M.; Geng, F.S.; Humphreys, D.T.; Kristianto, E.; Sheng, D.Z.; Hui, S.P.; Zhang, Y.; Sugimoto, K.; Nakayama, M.; Zheng, D.; et al. Krüppel-like factor 1 is a core cardiomyogenic trigger in zebrafish. Science 2021, 372, 201–205. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.W.; Zheng, L.; Li, L.J.; Yao, Y.F.; Hua, H.; Yang, S.Z.; Wen, Y.F.; Song, C.C.; Cao, X.K.; Liu, K.P.; et al. Novel copy number variation of the KLF3 gene is associated with growth traits in beef cattle. Gene 2019, 680, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Hou, Z.; Wang, Z.; Tao, Y.; Bai, J.; Yu, B.; Shen, J.; Sun, H.; Xiao, L.; Xu, Y.; Zhou, J.; et al. KLF2 regulates osteoblast differentiation by targeting of Runx2. Lab. Investig. 2019, 99, 271–280. [Google Scholar] [CrossRef] [PubMed]
- de Lázaro, I.; Yilmazer, A.; Nam, Y.; Qubisi, S.; Razak, F.M.A.; Degens, H.; Cossu, G.; Kostarelos, K. Non-viral, Tumor-free Induction of Transient Cell Reprogramming in Mouse Skeletal Muscle to Enhance Tissue Regeneration. Mol. Ther. 2019, 27, 59–75. [Google Scholar] [CrossRef]
- Hayashi, S.; Manabe, I.; Suzuki, Y.; Relaix, F.; Oishi, Y. Klf5 regulates muscle differentiation by directly targeting muscle-specific genes in cooperation with MyoD in mice. Elife 2016, 5, e17462–e17484. [Google Scholar] [CrossRef]
- Dionyssiou, M.G.; Salma, J.; Bevzyuk, M.; Wales, S.; Zakharyan, L.; McDermott, J.C. Krüppel-like factor 6 (KLF6) promotes cell proliferation in skeletal myoblasts in response to TGFβ/Smad3 signaling. Skelet. Muscle 2013, 3, 7. [Google Scholar] [CrossRef]
- Wang, X.; Shen, Q.W.; Wang, J.; Zhang, Z.; Feng, F.; Chen, T.; Zhang, Y.; Wei, H.; Li, Z.; Wang, X.; et al. KLF7 Regulates Satellite Cell Quiescence in Response to Extracellular Signaling. Stem. Cells 2016, 34, 1310–1320. [Google Scholar] [CrossRef]
- Fan, L.; Sweet, D.R.; Fan, E.K.; Prosdocimo, D.A.; Madera, A.; Jiang, Z.; Padmanabhan, R.; Haldar, S.M.; Vinayachandran, V.; Jain, M.K. Transcription factors KLF15 and PPARδ cooperatively orchestrate genome-wide regulation of lipid metabolism in skeletal muscle. J. Biol. Chem. 2022, 298, 101926. [Google Scholar] [CrossRef] [PubMed]
- Cui, A.; Fan, H.; Zhang, Y.; Zhang, Y.; Niu, D.; Liu, S.; Liu, Q.; Ma, W.; Shen, Z.; Shen, L.; et al. Dexamethasone-induced Krüppel-like factor 9 expression promotes hepatic gluconeogenesis and hyperglycemia. J. Clin. Investig. 2019, 129, 2266–2278. [Google Scholar] [CrossRef] [PubMed]
- Lam, V.C.; Folkersen, L.; Aguilar, O.A.; Lanier, L.L. KLF12 Regulates Mouse NK Cell Proliferation. J. Immunol. 2019, 203, 981–989. [Google Scholar] [CrossRef]
- Cherukunnath, A.; Davargaon, R.S.; Ashraf, R.; Kamdar, U.; Srivastava, A.K.; Tripathi, P.P.; Chatterjee, N.; Kumar, S. KLF8 is activated by TGF-β1 via Smad2 and contributes to ovarian cancer progression. J. Cell. Biochem. 2022, 123, 921–934. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; Chang, Z.; Zhao, Y.; Guo, Y.; Lu, H.; Liang, W.; Rom, O.; Wang, H.; Sun, J.; Zhu, T.; et al. KLF11 protects against abdominal aortic aneurysm through inhibition of endothelial cell dysfunction. JCI Insight 2021, 6, e141673. [Google Scholar] [CrossRef]
- Yao, W.; Jiao, Y.; Zhou, Y.; Luo, X. KLF13 suppresses the proliferation and growth of colorectal cancer cells through transcriptionally inhibiting HMGCS1-mediated cholesterol biosynthesis. Cell Biosci. 2020, 10, 76. [Google Scholar] [CrossRef] [PubMed]
- Xin, Y.; Su, P.; Liu, Y.; Gu, S.; An, X.; Zhang, X.; Yan, J.; Guo, Y.; Zhou, J.; Yang, G. Knock out hepatic Krüppel-like factor 16 (KLF16) improve myocardial damage and promoted myocardial protection of myocardial ischemia-reperfusion via anti-oxidative and anti-inflammation effects by TFAM/PPARβ signal passage. Bioengineered 2021, 12, 10219–10231. [Google Scholar] [CrossRef]
- Wang, S.H.; Hao, J.; Zhang, C.; Duan, F.F.; Chiu, Y.T.; Shi, M.; Huang, X.; Yang, J.; Cao, H.; Wang, Y. KLF17 promotes human naive pluripotency through repressing MAPK3 and ZIC2. Sci. China Life Sci. 2022, 65, 1985–1997. [Google Scholar] [CrossRef]
- Kammoun, M.; Pouletaut, P.; Morandat, S.; Subramaniam, M.; Hawse, J.R.; Bensamoun, S.F. Krüppel-like factor 10 regulates the contractile properties of skeletal muscle fibers in mice. Muscle Nerve 2021, 64, 765–769. [Google Scholar] [CrossRef]
- Emilia, Q.; Groover, K.; Clark, J.; Le, T.; Burrowes, B.; Liu, M. Complete Genome Sequence of Stenotrophomonas maltophilia Siphophage Suso. Microbiol. Resour. Announc. 2022, 11, e0011722. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, L.; Zou, Q.; Lin, C. prPred-DRLF: Plant R protein predictor using deep representation learning features. Proteomics 2022, 22, e2100161. [Google Scholar] [CrossRef] [PubMed]
- Randhawa, G.S.; Hill, K.A.; Kari, L. ML-DSP: Machine Learning with Digital Signal Processing for ultrafast, accurate, and scalable genome classification at all taxonomic levels. BMC Genom. 2019, 20, 267. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.Y.; Zhang, Q.F.; Jiang, D.D.; Liu, Y.F.; Chen, B.; Yang, S.P.; Shao, Z.T.; Jiang, H.; Wang, J.; Fang, Y.H.; et al. Complete mitogenomes and phylogenetic relationships of Haemaphysalis nepalensis and Haemaphysalis yeni. Front. Vet. Sci. 2022, 9, 1007631. [Google Scholar] [CrossRef]
- Wang, J.; Chitsaz, F.; Derbyshire, M.K.; Gonzales, N.R.; Gwadz, M.; Lu, S.; Marchler, G.H.; Song, J.S.; Thanki, N.; Yamashita, R.A.; et al. The conserved domain database in 2023. Nucleic Acids Res. 2023, 51, D384–D388. [Google Scholar] [CrossRef]
- Perkel, J.M. Smart software untangles gene regulation in cells. Nature 2022, 609, 428–431. [Google Scholar] [CrossRef] [PubMed]
- Martin, F.J.; Amode, M.R.; Aneja, A.; Austine-Orimoloye, O.; Azov, A.G.; Barnes, I.; Becker, A.; Bennett, R.; Berry, A.; Bhai, J.; et al. Ensembl 2023. Nucleic Acids Res. 2023, 51, D933–D941. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Ngo, V.; Wang, M.; Wang, W. Finding de novo methylated DNA motifs. Bioinformatics 2019, 35, 3287–3293. [Google Scholar] [CrossRef]
- Doncheva, N.T.; Morris, J.H.; Holze, H.; Kirsch, R.; Nastou, K.C.; Cuesta-Astroz, Y.; Rattei, T.; Szklarczyk, D.; von Mering, C.; Jensen, L.J. Cytoscape stringApp 2.0: Analysis and Visualization of Heterogeneous Biological Networks. J. Proteome Res. 2023, 22, 637–646. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Nastou, K.C.; Lyon, D.; Kirsch, R.; Pyysalo, S.; Doncheva, N.T.; Legeay, M.; Fang, T.; Bork, P.; et al. The STRING database in 2021: Customizable protein-protein networks, and functional characterization of user-uploaded gene/measurement sets. Nucleic Acids Res. 2021, 49, D605–D612. [Google Scholar] [CrossRef]
- Qiu, S.; Yu, G.; Lu, X.; Domeniconi, C.; Guo, M. Isoform function prediction by Gene Ontology embedding. Bioinformatics 2022, 38, 4581–4588. [Google Scholar] [CrossRef] [PubMed]
- Bu, D.; Luo, H.; Huo, P.; Wang, Z.; Zhang, S.; He, Z.; Wu, Y.; Zhao, L.; Liu, J.; Guo, J.; et al. KOBAS-i: Intelligent prioritization and exploratory visualization of biological functions for gene enrichment analysis. Nucleic Acids Res. 2021, 49, W317–W325. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Ren, X.; Li, L.; Lu, S.; Chen, T.; Tan, L.; Liu, M.; Luo, Q.; Liang, S.; Nie, Q.; et al. Integrative Analyses of mRNA Expression Profile Reveal the Involvement of IGF2BP1 in Chicken Adipogenesis. Int. J. Mol. Sci. 2019, 20, 2923. [Google Scholar] [CrossRef]
- Jiang, A.; Lehnert, K.; You, L.; Snell, R.G. ICARUS, an interactive web server for single cell RNA-seq analysis. Nucleic Acids Res. 2022, 50, W427–W433. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Ouyang, H.; Zheng, M.; Cai, B.; Han, P.; Abdalla, B.A.; Nie, Q.; Zhang, X. Integrated Analysis of Long Non-coding RNAs (LncRNAs) and mRNA Expression Profiles Reveals the Potential Role of LncRNAs in Skeletal Muscle Development of the Chicken. Front. Physiol. 2017, 7, 687. [Google Scholar] [CrossRef]
- Zhang, G.; Zhang, J.; Wu, P.; Ling, X.; Wang, Q.; Zhou, K.; Li, P.; Zhang, L.; Ye, H.; Zhang, Q.; et al. Transcriptome Sequencing Analysis of circRNA in Skeletal Muscle between Fast- and Slow-Growing Chickens at Embryonic Stages. Animals 2022, 12, 3166. [Google Scholar] [CrossRef] [PubMed]
- Khumpeerawat, P.; Duangjinda, M.; Phasuk, Y. Factors affecting gene expression associated with the skin color of black-bone chicken in Thailand. Poult. Sci. 2021, 100, 101440. [Google Scholar] [CrossRef]
- Kanehisa, M.; Furumichi, M.; Sato, Y.; Kawashima, M.; Ishiguro-Watanabe, M. KEGG for taxonomy-based analysis of pathways and genomes. Nucleic Acids Res. 2023, 51, D587–D592. [Google Scholar] [CrossRef]
- Li, Y.; Zhao, X.; Xu, M.; Chen, M. Krüppel-like factors in glycolipid metabolic diseases. Mol. Biol. Rep. 2022, 49, 8145–8152. [Google Scholar] [CrossRef]
- Yang, G.; Lu, H.; Wang, L.; Zhao, J.; Zeng, W.; Zhang, T. Genome-Wide Identification and Transcriptional Expression of the METTL21C Gene Family in Chicken. Genes 2019, 10, 628. [Google Scholar] [CrossRef]
- Lei, P.; Wei, X.; Gao, R.; Huo, F.; Nie, X.; Tong, W.; Song, W. Genome-wide identification of PYL gene family in wheat: Evolution, expression and 3D structure analysis. Genomics 2021, 113, 854–866. [Google Scholar] [CrossRef]
- Liu, D.; Li, J.; Hao, W.; Lin, X.; Xia, J.; Zhu, J.; Yang, S.; Yang, X. Chimeric RNA TNNI2-ACTA1-V1 Regulates Cell Proliferation by Regulating the Expression of NCOA3. Front. Vet. Sci. 2022, 9, 895190. [Google Scholar] [CrossRef] [PubMed]
- Han, H.; Gu, S.; Chu, W.; Sun, W.; Wei, W.; Dang, X.; Tian, Y.; Liu, K.; Chen, J. miR-17-5p Regulates Differential Expression of NCOA3 in Pig Intramuscular and Subcutaneous Adipose Tissue. Lipids 2017, 52, 939–949. [Google Scholar] [CrossRef] [PubMed]
- Nematbakhsh, S.; Pei Pei, C.; Selamat, J.; Nordin, N.; Idris, L.H.; Abdull Razis, A.F. Molecular Regulation of Lipogenesis, Adipogenesis and Fat Deposition in Chicken. Genes 2021, 12, 414. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.W.; Rong, E.G.; Shi, M.X.; Wu, C.Y.; Sun, B.; Wang, Y.X.; Wang, N.; Li, H. Expression and functional analysis of Krüppel-like factor 2 in chicken adipose tissue. J. Anim. Sci. 2014, 92, 4797–4805. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Raza, S.H.A.; Schreurs, N.M.; Khan, R.; Wei, D.; Wang, L.; Zhang, S.; Zhang, L.; Wu, S.; Ullah, I.; et al. Genetic variants in the promoter region of the KLF3 gene associated with fat deposition in Qinchuan cattle. Gene 2018, 672, 50–55. [Google Scholar] [CrossRef]
- Liu, H.; Wei, W.; Lin, W.; Yu, W.; Luo, W.; Niu, Y.; Zhang, L.; Chen, J. miR-32-5p Regulates Lipid Accumulation in Intramuscular Fat of Erhualian Pigs by Suppressing KLF3. Lipids 2021, 56, 279–287. [Google Scholar] [CrossRef]
- Guo, H.; Khan, R.; Raza, S.H.A.; Ning, Y.; Wei, D.; Wu, S.; Hosseini, S.M.; Ullah, I.; Garcia, M.D.; Zan, L. KLF15 promotes transcription of KLF3 gene in bovine adipocytes. Gene 2018, 659, 77–83. [Google Scholar] [CrossRef]
- Ding, L.; Li, S.; Wang, F.; Xu, J.; Li, S.; Wang, B.; Kou, J.; Wang, Y.; Cao, W. Berberine improves dietary-induced cardiac remodeling by upregulating Kruppel-like factor 4-dependent mitochondrial function. Biol. Chem. 2021, 402, 795–803. [Google Scholar] [CrossRef]
- Pol, C.J.; Pollak, N.M.; Jurczak, M.J.; Zacharia, E.; Karagiannides, I.; Kyriazis, I.D.; Ntziachristos, P.; Scerbo, D.A.; Brown, B.R.; Aifantis, I.; et al. Cardiac myocyte KLF5 regulates body weight via alteration of cardiac FGF21. Biochim. Biophys Acta Mol. Basis. Dis. 2019, 1865, 2125–2137. [Google Scholar] [CrossRef]
- Iqbal, A.; Yu, H.; Jiang, P.; Zhao, Z. Deciphering the Key Regulatory Roles of KLF6 and Bta-miR-148a on Milk Fat Metabolism in Bovine Mammary Epithelial Cells. Genes 2022, 13, 1828. [Google Scholar] [CrossRef] [PubMed]
- Raza, S.H.A.; Khan, R.; Cheng, G.; Long, F.; Bing, S.; Easa, A.A.; Schreurs, N.M.; Pant, S.D.; Zhang, W.; Li, A.; et al. RNA-Seq reveals the potential molecular mechanisms of bovine KLF6 gene in the regulation of adipogenesis. Int. J. Biol. Macromol. 2022, 195, 198–206. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Wang, H.; Sun, Y.; Li, H.; Wang, N. Klf7 modulates the differentiation and proliferation of chicken preadipocyte. Acta Biochim. Biophys Sin. 2013, 45, 280–288. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Wang, Y.; Li, X.; Du, Y.; Li, Y.; Zhu, J.; Lin, Y. miR-10a-5p Inhibits the Differentiation of Goat Intramuscular Preadipocytes by Targeting KLF8 in Goats. Front. Mol. Biosci. 2021, 8, 700078. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.R.; Zhang, M.; Sun, J.W.; Li, F.; Ma, X.F.; Li, W.T.; Han, R.L.; Li, Z.J.; Jiang, R.R.; Li, G.X.; et al. Krüppel-like factor KLF9 inhibits chicken intramuscular preadipocyte differentiation. Br. Poult. Sci. 2019, 60, 790–797. [Google Scholar] [CrossRef]
- Raza, S.H.A.; Pant, S.D.; Wani, A.K.; Mohamed, H.H.; Khalifa, N.E.; Almohaimeed, H.M.; Alshanwani, A.R.; Assiri, R.; Aggad, W.S.; Noreldin, A.E.; et al. Krüppel-like factors family regulation of adipogenic markers genes in bovine cattle adipogenesis. Mol. Cell. Probes. 2022, 65, 101850. [Google Scholar] [CrossRef]
- Manoharan, P.; Song, T.; Radzyukevich, T.L.; Sadayappan, S.; Lingrel, J.B.; Heiny, J.A. KLF2 in Myeloid Lineage Cells Regulates the Innate Immune Response during Skeletal Muscle Injury and Regeneration. iScience 2019, 17, 334–346. [Google Scholar] [CrossRef]
- Chen, K.; Gao, P.; Li, Z.; Dai, A.; Yang, M.; Chen, S.; Su, J.; Deng, Z.; Li, L. Forkhead Box O Signaling Pathway in Skeletal Muscle Atrophy. Am. J. Pathol. 2022, 192, 1648–1657. [Google Scholar] [CrossRef]
- Zhang, D.; Ran, J.; Li, J.; Yu, C.; Cui, Z.; Amevor, F.K.; Wang, Y.; Jiang, X.; Qiu, M.; Du, H.; et al. miR-21-5p Regulates the Proliferation and Differentiation of Skeletal Muscle Satellite Cells by Targeting KLF3 in Chicken. Genes 2021, 12, 814. [Google Scholar] [CrossRef]
- Zhang, G.; Chen, F.; Wu, P.; Li, T.; He, M.; Yin, X.; Shi, H.; Duan, Y.; Zhang, T.; Wang, J.; et al. MicroRNA-7 Targets the KLF4 Gene to Regulate the Proliferation and Differentiation of Chicken Primary Myoblasts. Front. Genet 2020, 11, 842. [Google Scholar] [CrossRef]
- Zhang, D.H.; Yin, H.D.; Li, J.J.; Wang, Y.; Yang, C.W.; Jiang, X.S.; Du, H.R.; Liu, Y.P. KLF5 regulates chicken skeletal muscle atrophy via the canonical Wnt/β-catenin signaling pathway. Exp. Anim. 2020, 69, 430–440. [Google Scholar] [CrossRef] [PubMed]
- Song, C.; Yang, J.; Jiang, R.; Yang, Z.; Li, H.; Huang, Y.; Lan, X.; Lei, C.; Ma, Y.; Qi, X.; et al. miR-148a-3p regulates proliferation and apoptosis of bovine muscle cells by targeting KLF6. J. Cell. Physiol. 2019, 234, 15742–15750. [Google Scholar] [CrossRef] [PubMed]
- Raza, S.H.A.; Khan, R.; Schreurs, N.M.; Guo, H.; Gui, L.S.; Mei, C.; Zan, L. Expression of the bovine KLF6 gene polymorphisms and their association with carcass and body measures in Qinchuan cattle (Bos Taurus). Genomics 2020, 112, 423–431. [Google Scholar] [CrossRef]
- Naldurtiker, A.; Batchu, P.; Kouakou, B.; Terrill, T.H.; Shaik, A.; Kannan, G. RNA-Seq exploration of the influence of stress on meat quality in Spanish goats. Sci. Rep. 2022, 12, 20573. [Google Scholar] [CrossRef]
- Baroukh, N.; Canteleux, N.; Lefèvre, A.; Dupuy, C.; Martias, C.; Presset, A.; Subramaniam, M.; Hawse, J.R.; Emond, P.; Pouletaut, P.; et al. Serum and Soleus Metabolomics Signature of Klf10 Knockout Mice to Identify Potential Biomarkers. Metabolites 2022, 12, 556. [Google Scholar] [CrossRef]
- Tang, H.; Liu, D.; Zhang, H.; Fan, W.; Hu, J.; Xu, Y.; Guo, Z.; Huang, W.; Hou, S.; Zhou, Z. Genome-wide association studies demonstrate the genes associated with perimysial thickness in ducks. Anim. Genet. 2023, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; An, X.; Bao, L.; Li, Y.; Pan, Y.; He, J.; Liu, L.; Zhu, X.; Zhang, J.; Cheng, J.; et al. MiR-125a-3p-KLF15-BCAA Regulates the Skeletal Muscle Branched-Chain Amino Acid Metabolism in Nile Tilapia (Oreochromis niloticus) During Starvation. Front. Genet. 2020, 11, 852. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene ID | Protein ID | CDS ID | Chromosome | Gene Position | |
|---|---|---|---|---|---|---|
| Start | End | |||||
| Gg-KLF2 | ENSGALG00000003939 | ENSGALP00000006265 | ENSGALT00000006275 | Chr28 | 4484818 | 4487291 |
| Gg-KLF3 | ENSGALG00000041717 | ENSGALP00000053488 | ENSGALT00000090511 | Chr4 | 69533898 | 69558360 |
| Gg-KLF4 | ENSGALG00000047208 | ENSGALP00000065832 | ENSGALT00000097875 | Chr8 | 20767770 | 20771114 |
| Gg-KLF5 | ENSGALG00000016927 | ENSGALP00000086539 | ENSGALT00000124066 | Chr1 | 157437529 | 157456371 |
| Gg-KLF6 | ENSGALG00000033178 | ENSGALP00000049438 | ENSGALT00000073174 | Chr2 | 11821968 | 11830999 |
| Gg-KLF7 | ENSGALG00000008501 | ENSGALP00000013833 | ENSGALT00000013848 | Chr7 | 12266259 | 12323221 |
| Gg-KLF8 | ENSGALG00000037361 | ENSGALP00000068386 | ENSGALT00000099301 | Chr4 | 11462278 | 11469482 |
| Gg-KLF9 | ENSGALG00000027374 | ENSGALP00000043265 | ENSGALT00000043393 | chrZ | 35032072 | 35045242 |
| Gg-KLF10 | ENSGALG00000037913 | ENSGALP00000055499 | ENSGALT00000059886 | Chr2 | 129099993 | 129104922 |
| Gg-KLF11 | ENSGALG00000016440 | ENSGALP00000075856 | ENSGALT00000118896 | Chr3 | 96486867 | 96495981 |
| Gg-KLF12 | ENSGALG00000016926 | ENSGALP00000067131 | ENSGALT00000099648 | Chr1 | 156971380 | 157226168 |
| Gg-KLF13 | ENSGALG00000026590 | ENSGALP00000081341 | ENSGALT00000120340 | Chr10 | 6022200 | 6048656 |
| Gg-KLF15 | ENSGALG00000036133 | ENSGALP00000094959 | ENSGALT00000124262 | Chr12 | 11023377 | 11040828 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ling, X.; Wang, Q.; Zhang, J.; Zhang, G. Genome-Wide Analysis of the KLF Gene Family in Chicken: Characterization and Expression Profile. Animals 2023, 13, 1429. https://doi.org/10.3390/ani13091429
Ling X, Wang Q, Zhang J, Zhang G. Genome-Wide Analysis of the KLF Gene Family in Chicken: Characterization and Expression Profile. Animals. 2023; 13(9):1429. https://doi.org/10.3390/ani13091429
Chicago/Turabian StyleLing, Xuanze, Qifan Wang, Jin Zhang, and Genxi Zhang. 2023. "Genome-Wide Analysis of the KLF Gene Family in Chicken: Characterization and Expression Profile" Animals 13, no. 9: 1429. https://doi.org/10.3390/ani13091429