


Comparison of Immune-Related Gene Expression in Two Chicken Breeds Following Infectious Bronchitis Virus Vaccination

 , , and
, , and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
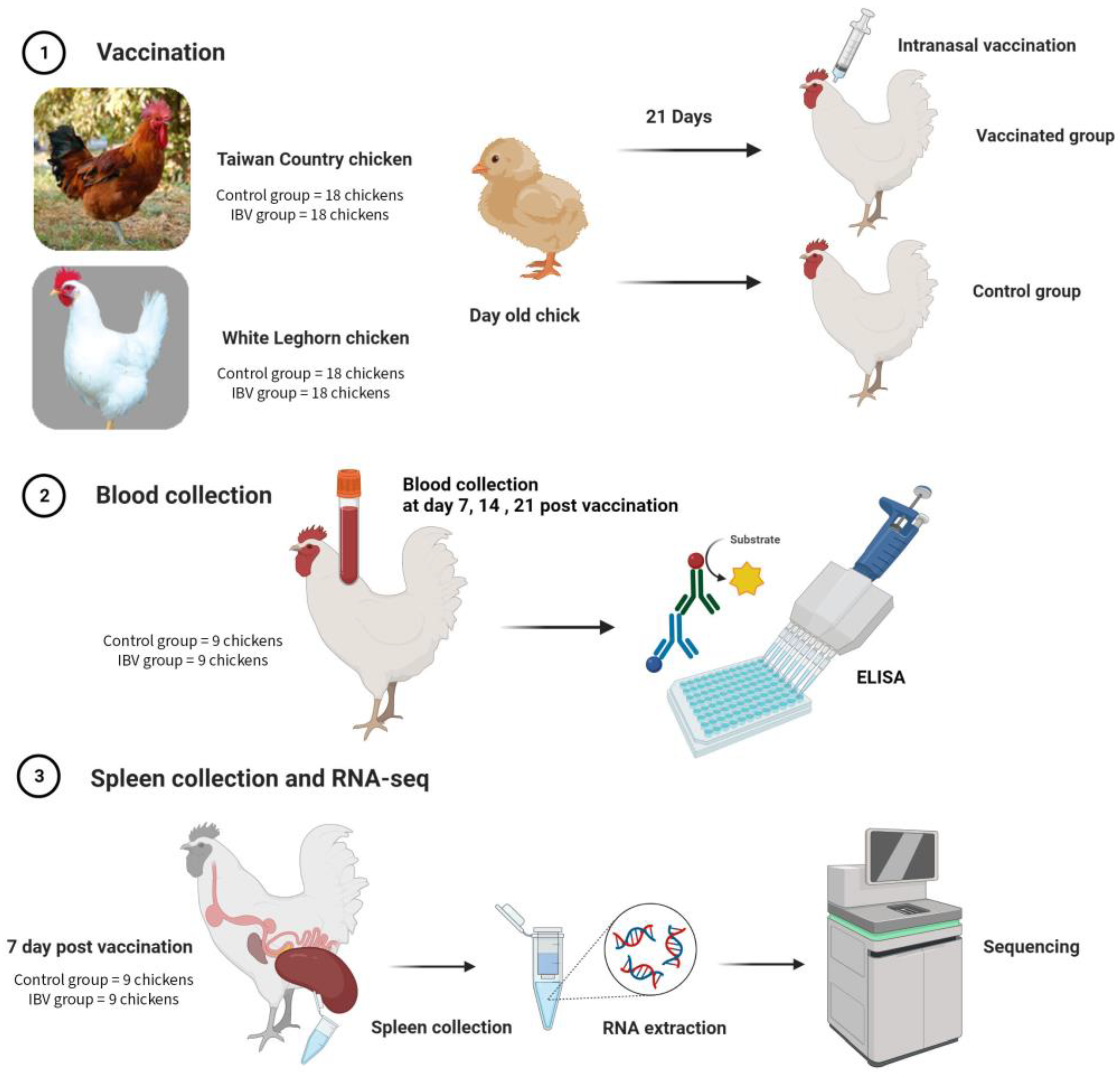
2.1. Animals and Tissue Sampling
2.2. Enzyme-Linked Immunosorbent Assay
2.3. RNA Extraction and cDNA Synthesis
2.4. Library Construction and RNA Sequencing
2.5. RNA-Seq Data Analysis and Go and KEGG Enrichment Analysis
2.6. Quantitative Real-Time PCR Verification
2.7. Statistical Analysis
3. Results
3.1. Anti-IBV ELISA Titers
3.2. Splenic Transcriptome Sequencing
3.3. Differentially Expressed Gene Profiling
3.4. GO and KEGG Databases of DEGs
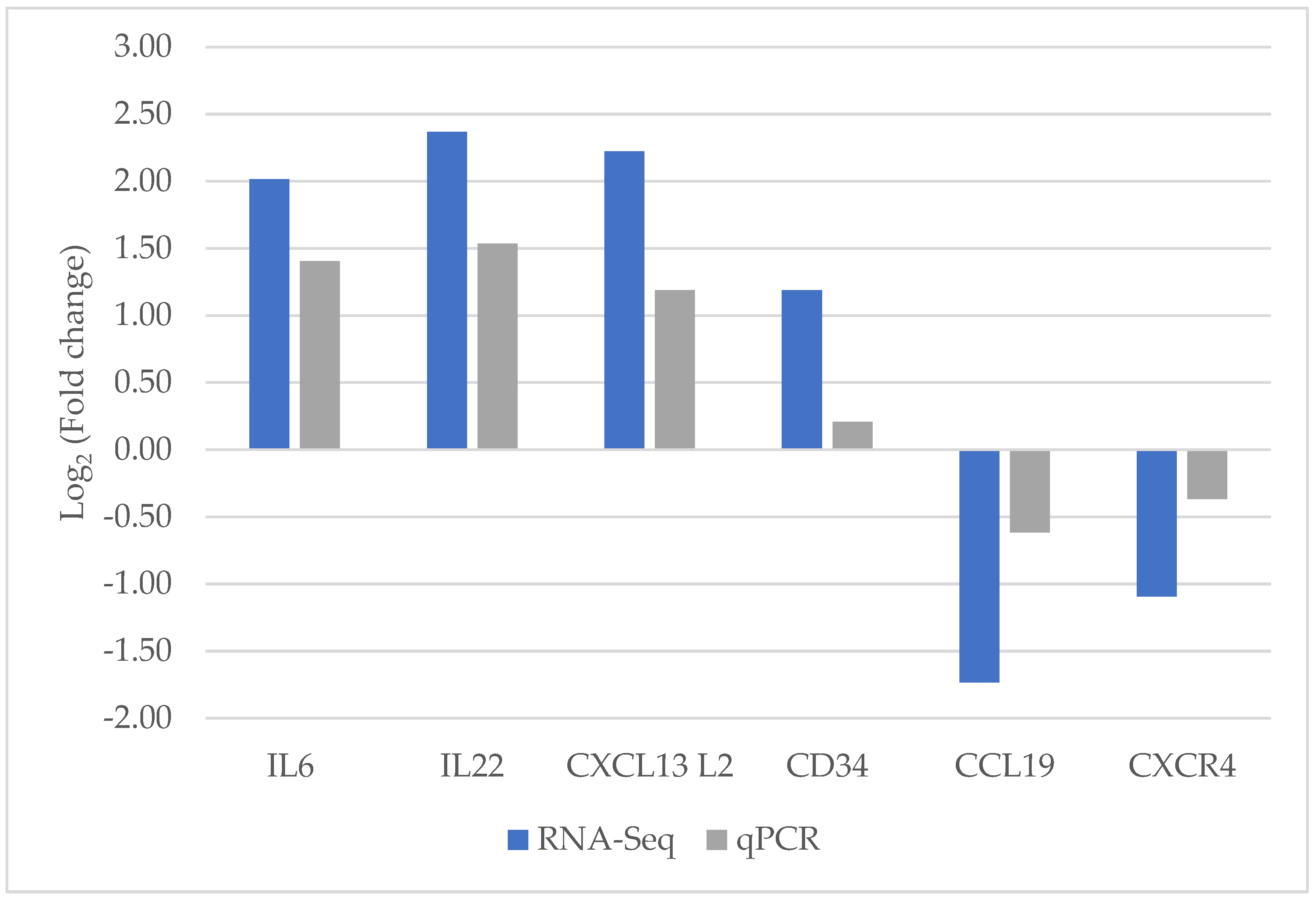
3.5. Validation Based on qPCR
4. Discussion
4.1. Immune Responsiveness in Taiwan Country Chickens
4.2. Immune Responsiveness in White Leghorn Chickens
4.3. Differences of Immune Responsiveness between Breeds
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- De Wit, J.J.; Cook, J.K.A. Spotlight on avian coronaviruses. Avian Pathol. 2020, 49, 313–316. [Google Scholar] [CrossRef] [PubMed]
- Ignjatović, J.; Sapats, S. Avian infectious bronchitis virus. Rev.—Off. Int. Epizoot. 2000, 19, 493–508. [Google Scholar] [CrossRef] [PubMed]
- Chhabra, R.; Forrester, A.; Lemiere, S.; Awad, F.; Chantrey, J.; Ganapathy, K. Mucosal, cellular, and humoral immune responses induced by different live infectious bronchitis virus vaccination regimes and protection conferred against infectious bronchitis virus Q1 strain. Clin. Vaccine Immunol. 2015, 22, 1050–1059. [Google Scholar] [CrossRef] [PubMed]
- Jackwood, M.W. Review of infectious bronchitis virus around the world. Avian Dis. 2012, 56, 634–641. [Google Scholar] [CrossRef] [PubMed]
- Roberts, J.R.; Chousalkar, K.K. Chapter 52—Infectious Bronchitis. In Egg Innovations and Strategies for Improvements; Hester, P.Y., Ed.; Academic Press: San Diego, CA, USA, 2017; pp. 561–570. [Google Scholar]
- Samiullah, S.; Roberts, J.; Chousalkar, K. Infectious bronchitis virus and brown shell colour: Australian strains of infectious bronchitis virus affect brown eggshell colour in commercial laying hens differently. Avian Pathol. 2016, 45, 552–558. [Google Scholar] [CrossRef]
- Hoerr, F.J. The pathology of infectious bronchitis. Avian Dis. 2021, 65, 600–611. [Google Scholar] [CrossRef]
- Iotsov, S.; Iotova, I.; Atanasov, K.; Nikolova, M.; Nikolov, N.D. Resistance of different breeds of laying poultry to Marek’s disease virus]. Vet. Med. Nauki 1983, 20, 3–8. [Google Scholar]
- Dunnington, E.A.; Larsen, C.T.; Gross, W.B.; Siegel, P.B. Antibody responses to combinations of antigens in white Leghorn chickens of different background genomes and major histocompatibility complex genotypes. Poult. Sci. 1992, 71, 1801–1806. [Google Scholar] [CrossRef]
- Kannaki, T.R.; Priyanka, E.; Haunshi, S. Research Note: Disease tolerance/resistance and host immune response to experimental infection with Pasteurella multocida A:1 isolate in Indian native Nicobari chicken breed. Poult. Sci. 2021, 100, 101268. [Google Scholar] [CrossRef]
- Khatab, S.A.; Hemeda, S.A.; El-Nahas, A.F.; Abd El Naby, W.S.H.; Hassan, S.; Alorabi, J.A.; Dawood, M.A.O. Intra- and inter-breed variation in immune response to acute and sub-chronic Salmonella infection and commercial immune-stimulant in two-layer breeds. Vet. Med. Sci. 2021, 7, 2353–2361. [Google Scholar] [CrossRef]
- Hedrick, P.W. Pathogen resistance and genetic variation at MHC loci. Evolution 2002, 56, 1902–1908. [Google Scholar] [CrossRef] [PubMed]
- Banat, G.R.; Tkalcic, S.; Dzielawa, J.A.; Jackwood, M.W.; Saggese, M.D.; Yates, L.; Kopulos, R.; Briles, W.E.; Collisson, E.W. Association of the chicken MHC B haplotypes with resistance to avian coronavirus. Dev. Comp. Immunol. 2013, 39, 430–437. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.; Sadeyen, J.R.; Cavanagh, D.; Kaiser, P.; Burt, D.W. The early immune response to infection of chickens with Infectious Bronchitis Virus (IBV) in susceptible and resistant birds. BMC Vet. Res. 2015, 11, 256. [Google Scholar] [CrossRef] [PubMed]
- Deist, M.S.; Gallardo, R.A.; Bunn, D.A.; Dekkers, J.C.M.; Zhou, H.; Lamont, S.J. Resistant and susceptible chicken lines show distinctive responses to Newcastle disease virus infection in the lung transcriptome. BMC Genomics 2017, 18, 989. [Google Scholar] [CrossRef]
- Samina, I.; Brenner, J.; Peleg, B.A. Differences in protection between heavy and light breeds of chickens following vaccination with Newcastle disease vaccines—A survey of data, 1971 to 1990. Avian Pathol. 1992, 21, 693–697. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.; Dunn, J.R.; Heidari, M.; Lee, L.F.; Song, J.; Ernst, C.W.; Ding, Z.; Bacon, L.D.; Zhang, H. Genetics and vaccine efficacy: Host genetic variation affecting Marek’s disease vaccine efficacy in White Leghorn chickens1. Poult. Sci. 2010, 89, 2083–2091. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.S.; Tixier-Boichard, M.; Chazara, O.; Lee, Y.P.; Chen, C.F.; Chang, P.C.; Chen, J.W.; Bed’hom, B. Different immune responses to three different vaccines following H6N1 low pathogenic avian influenza virus challenge in Taiwanese local chicken breeds. BMC Proc. 2011, 5 (Suppl. 4), S33. [Google Scholar] [CrossRef] [PubMed]
- Mountford, J.; Gheyas, A.; Vervelde, L.; Smith, J. Genetic variation in chicken interferon signalling pathway genes in research lines showing differential viral resistance. Anim. Genet. 2022, 53, 640–656. [Google Scholar] [CrossRef]
- Lowenthal, J.W.; Bean, A.G.; Kogut, M.H. What’s so special about chicken immunology? Dev. Comp. Immunol. 2013, 41, 307–309. [Google Scholar] [CrossRef]
- Jeurissen, S.H. The role of various compartments in the chicken spleen during an antigen-specific humoral response. Immunology 1993, 80, 29–33. [Google Scholar]
- Zhang, Q.; Waqas, Y.; Yang, P.; Sun, X.; Liu, Y.; Ahmed, N.; Chen, B.; Li, Q.; Hu, L.; Huang, Y.; et al. Cytological study on the regulation of lymphocyte homing in the chicken spleen during LPS stimulation. Oncotarget 2017, 8, 7405–7419. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Chen, B.; Yang, P.; Zhang, L.; Liu, Y.; Ullah, S.; Wu, L.; Waqas, Y.; Le, Y.; Chen, W.; et al. Identification and structural composition of the blood-spleen barrier in chickens. Vet. J. 2015, 204, 110–116. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.S. Disease Resistance to Marek’s Disease in Country Chickens and White Leghorns. Master’s Thesis, National Chung Hsing University, Taichung, Taiwan, 1987. [Google Scholar]
- Chao, C.H.; Lee, Y.P. Immune responses to Newcastle disease virus vaccine and sheep erythrocytes in country chickens and White Leghorns. J. Chin. Soc. Anim. Sci. 1991, 20, 189–201. [Google Scholar]
- Chen, C.F.; Lee, Y.P. Comparison of disease resistance to leucocytozoonosis among country chickens, broilers and White Leghorns. J. Chin. Soc. Anim. Sci. 1991, 30, 305–316. [Google Scholar]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. 2010. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 17 May 2018).
- Ewels, P.; Magnusson, M.; Lundin, S.; Käller, M. MultiQC: Summarize analysis results for multiple tools and samples in a single report. Bioinformatics 2016, 32, 3047–3048. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Kaufman, J. The simple chicken major histocompatibility complex: Life and death in the face of pathogens and vaccines. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2000, 355, 1077–1084. [Google Scholar] [CrossRef]
- Silva, A.P.D.; Gallardo, R.A. The chicken MHC: Insights into genetic resistance, immunity, and inflammation following infectious bronchitis virus infections. Vaccines 2020, 8, 637. [Google Scholar] [CrossRef]
- Viertlboeck, B.C.; Crooijmans, R.P.; Groenen, M.A.; Göbel, T.W. Chicken Ig-like receptor B2, a member of a multigene family, is mainly expressed on B lymphocytes, recruits both Src homology 2 domain containing protein tyrosine phosphatase (SHP)-1 and SHP-2, and inhibits proliferation. J. Immunol. 2004, 173, 7385–7393. [Google Scholar] [CrossRef] [PubMed]
- Mårtensson, I.L.; Ceredig, R. Review article: Role of the surrogate light chain and the pre-B-cell receptor in mouse B-cell development. Immunology 2000, 101, 435–441. [Google Scholar] [CrossRef]
- Rosnet, O.; Blanco-Betancourt, C.; Grivel, K.; Richter, K.; Schiff, C. Binding of free immunoglobulin light chains to VpreB3 inhibits their maturation and secretion in chicken B cells. J. Biol. Chem. 2004, 279, 10228–10236. [Google Scholar] [CrossRef] [PubMed]
- Dardenne, M.; Smaniotto, S.; de Mello-Coelho, V.; Villa-Verde, D.M.; Savino, W. Growth hormone modulates migration of developing T cells. Ann. N. Y. Acad. Sci. 2009, 1153, 1–5. [Google Scholar] [CrossRef]
- Velazquez-Salinas, L.; Verdugo-Rodriguez, A.; Rodriguez, L.L.; Borca, M.V. The role of interleukin 6 during viral infections. J. Immunol. 2019, 10, 1057. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.A.; Li, M.O. TGF-β: Guardian of T cell function. J. Immunol. 2013, 191, 3973–3979. [Google Scholar] [CrossRef] [PubMed]
- Romagnani, S. IL4I1: Key immunoregulator at a crossroads of divergent T-cell functions. Eur. J. Immunol. 2016, 46, 2302–2305. [Google Scholar] [CrossRef]
- Li, M.O.; Wan, Y.Y.; Sanjabi, S.; Robertson, A.-K.L.; Flavell, R.A. Transforming growth factor-β regulation of immune responses. Annu. Rev. Immunol. 2006, 24, 99–146. [Google Scholar] [CrossRef]
- Franitza, S.; Kollet, O.; Brill, A.; Vaday, G.G.; Petit, I.; Lapidot, T.; Alon, R.; Lider, O. TGF-beta1 enhances SDF-1alpha-induced chemotaxis and homing of naive T cells by up-regulating CXCR4 expression and downstream cytoskeletal effector molecules. Eur. J. Immunol. 2002, 32, 193–202. [Google Scholar] [CrossRef]
- Gurung, A.; Kamble, N.; Kaufer, B.B.; Pathan, A.; Behboudi, S. Association of Marek’s disease induced immunosuppression with activation of a novel regulatory T cells in chickens. PLoS Pathog. 2017, 13, e1006745. [Google Scholar] [CrossRef]
- Molinier-Frenkel, V.; Prévost-Blondel, A.; Castellano, F. The IL4I1 Enzyme: A new player in the immunosuppressive tumor microenvironment. Cells 2019, 8, 757. [Google Scholar] [CrossRef]
- Eyerich, K.; Dimartino, V.; Cavani, A. IL-17 and IL-22 in immunity: Driving protection and pathology. Eur. J. Immunol. 2017, 47, 607–614. [Google Scholar] [CrossRef]
- Buck, C.A. Immunoglobulin superfamily: Structure, function and relationship to other receptor molecules. Semin. Cell Dev. Biol. 1992, 3, 179–188. [Google Scholar] [CrossRef]
- Tessarz, A.S.; Weiler, S.; Zanzinger, K.; Angelisová, P.; Horejsí, V.; Cerwenka, A. Non-T cell activation linker (NTAL) negatively regulates TREM-1/DAP12-induced inflammatory cytokine production in myeloid cells. J. Immunol. 2007, 178, 1991–1999. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.U.; Kim, L.K.; Choi, J.M. Revisiting the concept of targeting NFAT to control T cell immunity and autoimmune diseases. Front. Immunol. 2018, 9, 2747. [Google Scholar] [CrossRef] [PubMed]
- Dong, W.; Zhang, H.; Huang, H.; Zhou, J.; Hu, L.; Lian, A.; Zhu, L.; Ma, N.; Yang, P.; Wei, K.; et al. Chicken IgY Fc linked to Bordetella avium ompA and Taishan Pinus massoniana pollen polysaccharide adjuvant enhances macrophage function and specific immune responses. Front. Microbiol. 2016, 7, 1708. [Google Scholar] [CrossRef]
- Bouwman, A.C.; van Daalen, K.R.; Crnko, S.; Ten Broeke, T.; Bovenschen, N. Intracellular and extracellular roles of granzyme K. Front. Immunol. 2021, 12, 677707. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Day | TCC | WLC | ||
|---|---|---|---|---|
| Treatment | Control | Treatment | Control | |
| Day 0 | 1.78 ± 0.70 | 1.37 ± 0.36 | 1.72 ± 0.33 | 1.44 ± 0.28 |
| Day 7 | 3.83 a ± 1.15 | 1.04 b ± 0.13 | 1.68 ± 0.37 | 1.32 ± 0.37 |
| Day 14 | 31.40 a,x ± 6.55 | 1.04 b ± 0.37 | 7.90 a,y ± 1.36 | 0.88 b ± 0.21 |
| Day 21 | 38.00 a,x ± 5.73 | 0.93 b ± 0.35 | 8.79 a,y ± 1.43 | 1.02 b ± 0.17 |
| Breed | Upregulated X | Downregulated Y | Immune Related Genes | log2FoldChange | p-Value |
|---|---|---|---|---|---|
| TCC | 257 | 186 | Immunoglobulin superfamily DCC subclass member 4 X | 1.89 | 3.50 × 10−5 |
| Transforming growth factor beta receptor 3 x | 1.43 | 4.82 × 10−4 | |||
| Macrophage mannose receptor 1-like 2 X | 1.40 | 1.60 × 10−3 | |||
| Mitogen-activated protein kinase 10 X | 1.80 | 5.88 × 10−3 | |||
| Major histocompatibility complex, class I X | 2.49 | 5.66 × 10−3 | |||
| V-set pre-B cell surrogate light chain 3 X | 2.49 | 7.56 × 10−4 | |||
| Interleukin 6 Y | −2.86 | 4.42 × 10−3 | |||
| Interleukin 4 induced 1 Y | −2.65 | 9.62 × 10−3 | |||
| Interleukin 22 Y | −2.54 | 3.50 × 10−3 | |||
| WLC | 49 | 72 | Interleukin 4 induced 1 X | 1.45 | 4.18 × 10−5 |
| Interleukin 22 receptor subunit alpha 2 X | 1.26 | 8.11 × 10−4 | |||
| Interleukin 6 X | 2.02 | 5.26 × 10−3 | |||
| MHC class I polypeptide-related sequence A Y | −1.56 | 9.59 × 10−6 | |||
| Eosinophil peroxidase Y | −1.36 | 4.65 × 10−4 | |||
| Class I histocompatibility antigen, F10 alpha chain-like 3 Y | −1.17 | 3.06 × 10−3 |
| Challenge Status | Breed | Immune Related Genes | log2FoldChange | p-Value | |
|---|---|---|---|---|---|
| TCC | WLC | ||||
| Unvaccinated | 577 | 1118 | C-X-C, motif chemokine ligand 14 X | 6.46 | 4.74 × 10−7 |
| Interleukin 22 X | 4.62 | 1.05 × 10−4 | |||
| Interleukin 4 induced 1 X | 3.49 | 1.25 × 10−4 | |||
| Interleukin 6 X | 3.20 | 1.39 × 10−4 | |||
| Leukocyte immunoglobulin-like receptor subfamily B member 5 X | 4.98 | 2.63 × 10−3 | |||
| Interleukin 1 receptor-like 2 Y | −3.09 | 2.52 × 10−13 | |||
| TNF receptor superfamily member 19 Y | −1.58 | 3.14 × 10−8 | |||
| T cell-interacting, activating receptor on myeloid cells protein 1-like Y | −3.20 | 3.95 × 10−7 | |||
| Mitogen-activated protein kinase 10 Y | −1.82 | 3.49 × 10−03 | |||
| Vaccinated | 470 | 416 | Class I histocompatibility antigen, F10 alpha chain-like X | 6.82 | 4.78 × 10−83 |
| C-type lectin domain family 2 member L-like X | 3.78 | 3.35 × 10−25 | |||
| Major histocompatibility complex class II beta chain BLB1 X | 1.42 | 1.14 × 10−21 | |||
| IgG Fc-binding protein-like X | 1.49 | 4.84 × 10−15 | |||
| Interleukin 8-like 1 X | 1.54 | 1.03 × 10−3 | |||
| Major histocompatibility complex class II beta chain BLB2 Y | −1.07 | 1.61 × 10−13 | |||
| T cell surface glycoprotein CD8 alpha chain-like Y | −2.20 | 8.57 × 10−7 | |||
| Granzyme K Y | −1.07 | 1.00 × 10−7 | |||
| MHC class I antigen YF5 Y | −1.03 | 4.29 × 10−6 | |||
| Interleukin 22 receptor subunit alpha 2 Y | −1.51 | 9.32 × 10−5 | |||
| Interleukin 6 Y | −1.67 | 2.56 × 10−2 | |||
| Breed | Description | p-Value | Count | Genes ID |
|---|---|---|---|---|
| TCC | Cell-cell adhesion via plasma-membrane adhesion molecules | 3.60 × 10−4 | 5 | CDH3/DSCAML1/FLRT3/NFASC/SDK2 |
| Cell differentiation | 1.00 × 10−3 | 19 | ACE/ZP3/OLFM1/RNASE6/ACVR2B/BRINP1/ APOA1/DSCAML1/FLRT3/TLL1/RARB/MGP/ TAGLN/SERPINB10B/CSRP2/NFASC/SDK2/MSTN | |
| Chemotaxis | 0.164 | 3 | FLRT3/NFASC/MSTN | |
| Regulation of leukocyte migration | 0.214 | 1 | MSTN | |
| Myeloid cell differentiation | 2.29 × 10−4 | 2 | ACE/SERPINB10B | |
| WLC | Cell proliferation | 7.00 × 10−3 | 7 | NPPC/CDH13/ANKRD1/HPGDS/IL6/NTRK2/INHBA |
| Acute inflammatory response | 7.00 × 10−3 | 2 | FN1/IL6 | |
| Cell differentiation | 1.10 × 10−2 | 11 | NPPC/ANKRD1/VSIG1/TENM1/IL6/RPE65/THRB/NTRK2/INHBA/STMN2/BHLHE22 | |
| Regulation of cell proliferation | 1.10 × 10−2 | 6 | CDH13/ANKRD1/HPGDS/IL6/NTRK2/INHBA | |
| Positive regulation of cell adhesion | 5.20 × 10−2 | 2 | CDH13/IL6 | |
| Cell-cell adhesion via plasma-membrane adhesion molecules | 5.20 × 10−2 | 2 | CDH13/TENM1 |
| Breed | Description | p-Value | Count | Symbol |
|---|---|---|---|---|
| TCC | Cell adhesion molecules (CAMs) | 2.40 × 10−2 | 6 | BLB1/CDH3/CDH1/NRXN1/NFASC/CNTNAP2 |
| Cytokine-cytokine receptor interaction | 3.00 × 10−2 | 8 | GH/IL11RA/ACVR2B/IL8L1/BMP6/EDAR/CCR10/MSTN | |
| Phagosome | 9.70 × 10−2 | 5 | BLB1/LOC425049/MMR1L1/ATP6V0D2/TUBA1C | |
| NOD-like receptor signaling pathway | 0.202 | 4 | IL8SL1/CAMP/GBP/GBP4L | |
| Salmonella infection | 0.720 | 1 | IL8L1 | |
| Influenza A | 0.728 | 2 | BLB1/IL8L1 | |
| Toll-like receptor signaling pathway | 0.793 | 1 | IL8L1 | |
| Herpes simplex virus 1 infection | 0.941 | 1 | BLB1 | |
| WLC | Cytokine-cytokine receptor interaction | 6.70 × 10−2 | 5 | TNFSF15/IL6/IL18RAP/IL1RL2/INHBA |
| Cell adhesion molecules (CAMs) | 0.148 | 3 | NCAM2/NRXN3/BLB2 | |
| NOD-like receptor signaling pathway | 0.402 | 2 | IL6/CASR | |
| Influenza A | 0.448 | 2 | IL6/BLB2 | |
| Herpes simplex virus 1 infection | 0.492 | 2 | IL6/BLB2 | |
| Salmonella infection | 0.529 | 1 | IL6 | |
| Toll-like receptor signaling pathway | 0.606 | 1 | IL6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chuwatthanakhajorn, S.; Chang, C.-S.; Ganapathy, K.; Tang, P.-C.; Chen, C.-F. Comparison of Immune-Related Gene Expression in Two Chicken Breeds Following Infectious Bronchitis Virus Vaccination. Animals 2023, 13, 1642. https://doi.org/10.3390/ani13101642
Chuwatthanakhajorn S, Chang C-S, Ganapathy K, Tang P-C, Chen C-F. Comparison of Immune-Related Gene Expression in Two Chicken Breeds Following Infectious Bronchitis Virus Vaccination. Animals. 2023; 13(10):1642. https://doi.org/10.3390/ani13101642
Chicago/Turabian StyleChuwatthanakhajorn, Schwann, Chi-Sheng Chang, Kannan Ganapathy, Pin-Chi Tang, and Chih-Feng Chen. 2023. "Comparison of Immune-Related Gene Expression in Two Chicken Breeds Following Infectious Bronchitis Virus Vaccination" Animals 13, no. 10: 1642. https://doi.org/10.3390/ani13101642