


Identification of Duplication Genotypes of the Feathering Rate Gene in Chicken by a Multiplex PCR Following Electrophoresis and/or Sanger Sequencing


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Chickens and DNA Collection
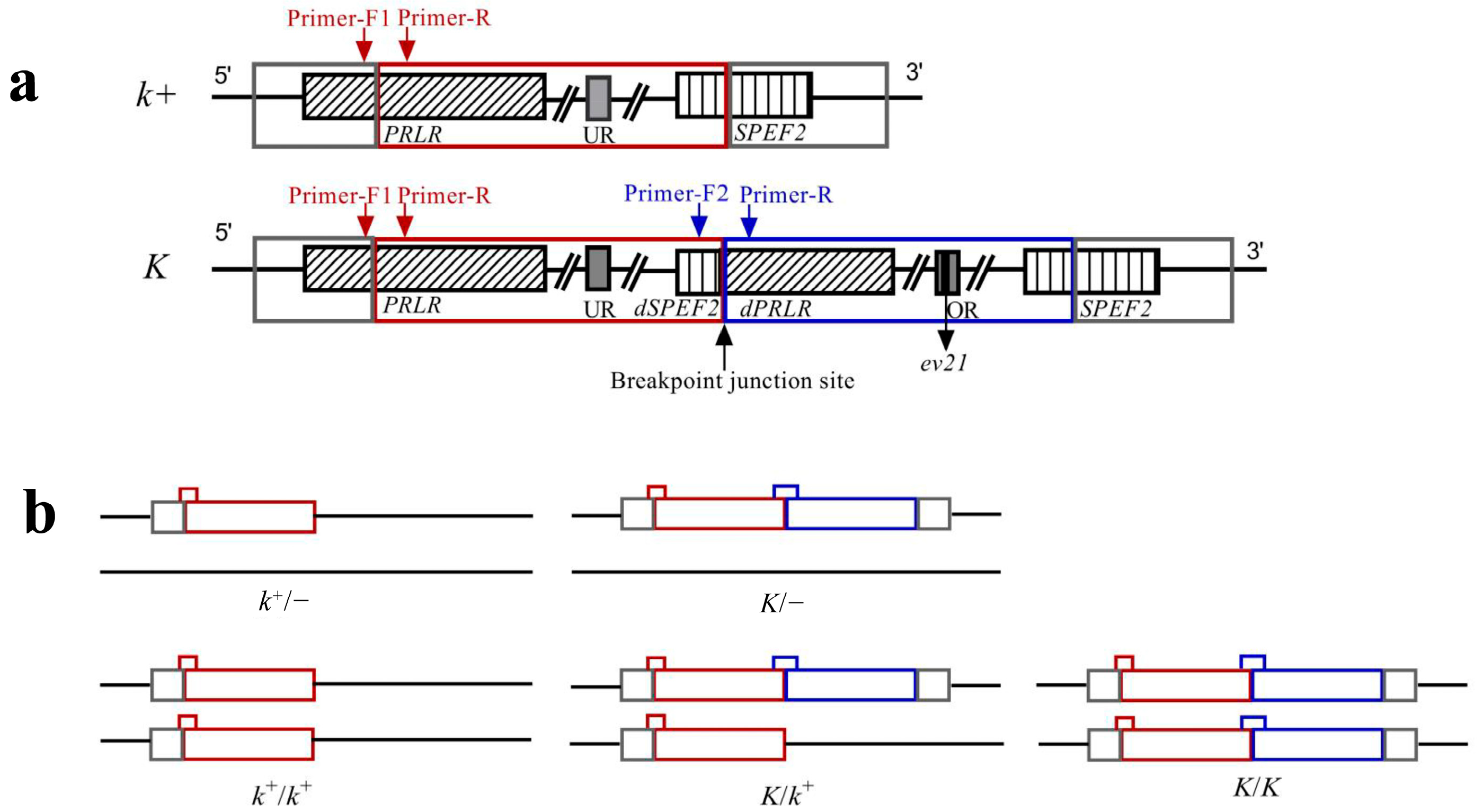
2.2. Primers
2.3. Optimization of Multiplex PCR Conditions
2.4. Genotyping and Verification of LF Males by Test Cross
3. Results
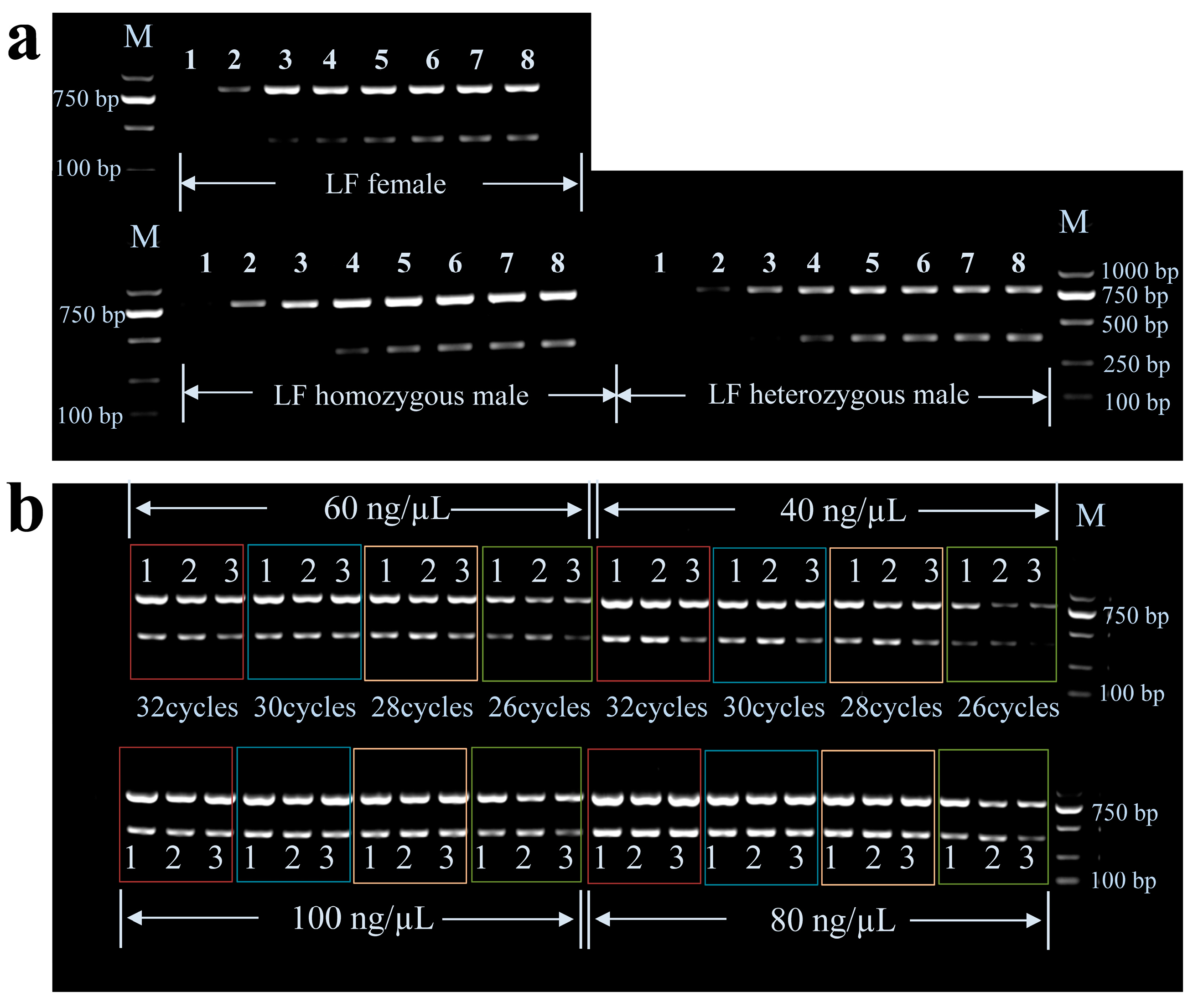
3.1. Optimization of Multiplex PCR Conditions
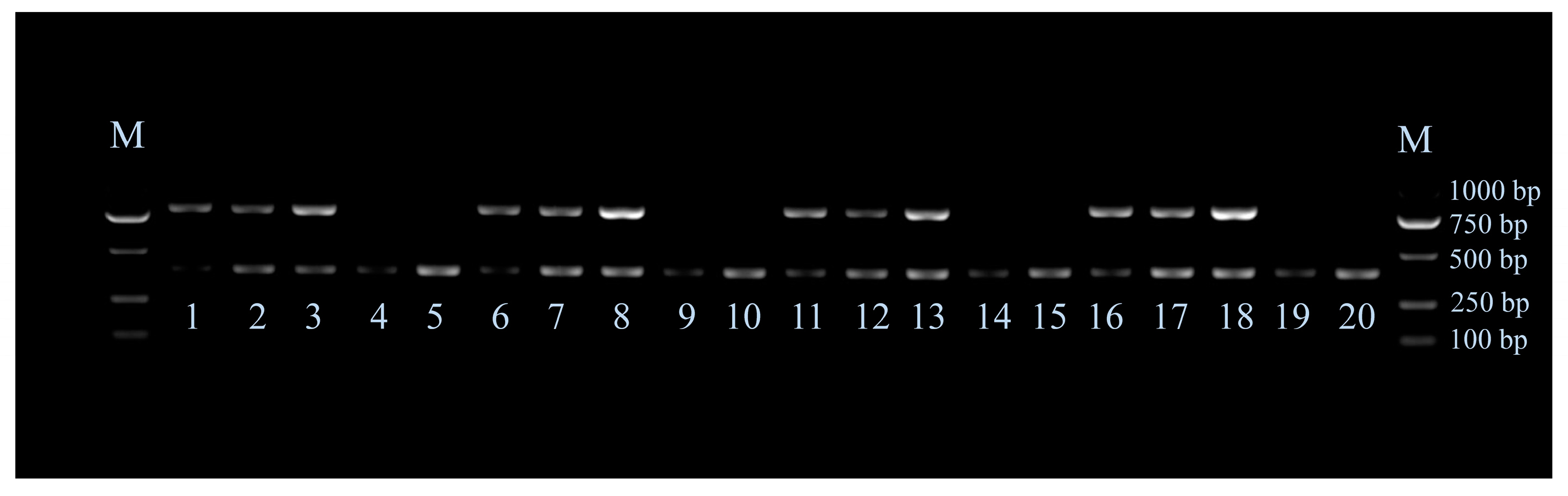
3.2. Genotyping by Multiplex PCR
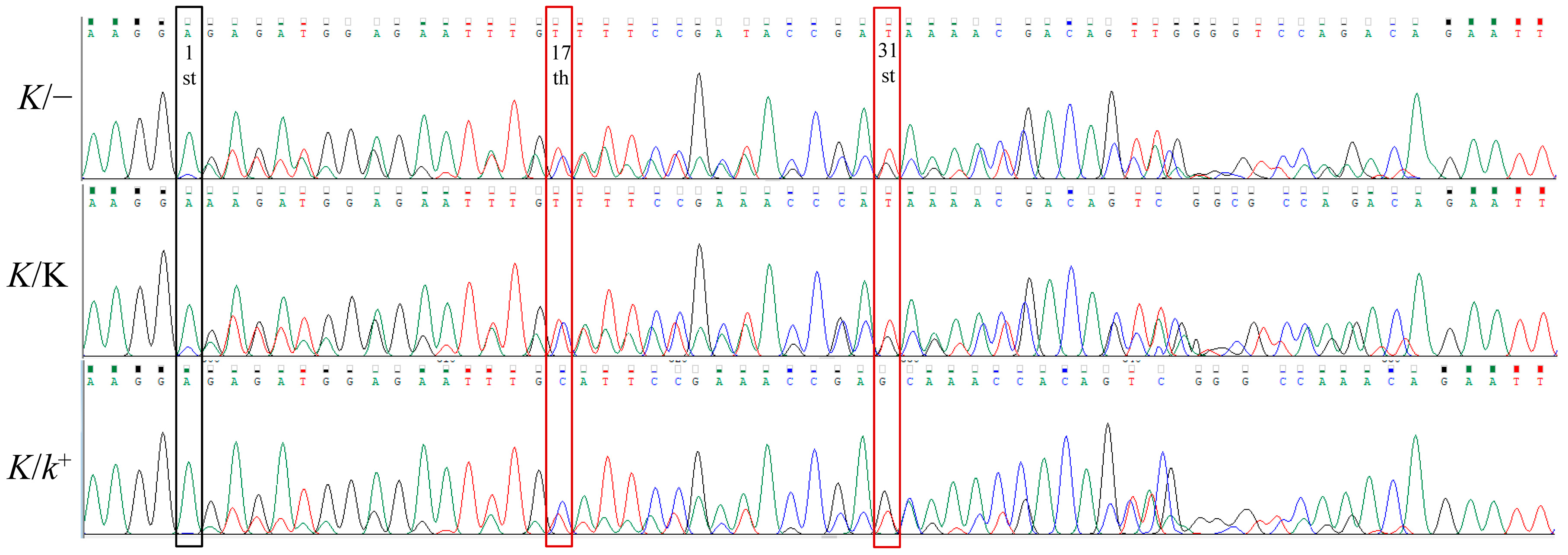
3.3. Verifying the Accuracy of the Genotyping Result of Late Feathering Roosters by Test Cross
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gryzinska, M.; Batkowska, J.; Andraszek, K.; Horecka, B.; Jezewska-Witkowska, G. Changes in plumage color and patterns in Polbar breed chicks (Polish conservative breed) during their first weeks after hatching. Eur. Poult. Sci. 2014, 78, 1–10. [Google Scholar] [CrossRef]
- Masui, K.; Hashimoto, J.J.S.B.C. Sexing Baby Chicks; Journal Printing Co., Limited: Vancouver, BC, Canada, 1933. [Google Scholar]
- Li, H.; Qiu, X.; Zeng, F.; Xie, H. Research on Feather Rate and Feather Type of Chicken in Different Strains and Their Offspring. J. Sichuan Agric. Univ. 1988, 3, 241–246. [Google Scholar]
- Serebrovsky, A.S. Crossing-over involving three sex-linked genes in chickens. Am. Nat. 1922, 56, 571–572. [Google Scholar] [CrossRef]
- Warren, D.C. Inheritance of rate of feathering in poultry. J. Hered. 1925, 16, 13–18. [Google Scholar] [CrossRef]
- Hertwig, P.; Rittershaus, T. Die Erbfaktoren der Haushühner. Z. Ver-Erbungslehre 1929, 51, 354–372. [Google Scholar] [CrossRef]
- Siegel, P.B.; Mueller, C.D.; Craig, J.V. Some Phenotypic Differences among Homozygous, Heterozygous, and Hemizygous Late Feathering Chicks. Poult. Sci. 1957, 36, 232–239. [Google Scholar] [CrossRef]
- Harris, D.L.; Garwood, V.A.; Lowe, P.C.; Hester, P.Y.; Crittenden, L.B.; Fadly, A.M. Influence of sex-linked feathering phenotypes of parents and progeny upon lymphoid leukosis virus infection status and egg production. Poult. Sci. 1984, 63, 401–413. [Google Scholar] [CrossRef] [PubMed]
- Smith, E.J.; Fadly, A.M. Influence of congenital transmission of endogenous virus-21 on the immune response to avian leukosis virus infection and the incidence of tumors in chickens. Poult. Sci. 1988, 67, 1674–1679. [Google Scholar] [CrossRef]
- Bacon, L.D.; Smith, E.; Crittenden, L.B.; Havenstein, G.B. Association of the Slow Feathering-(K) and an Endogenous Viral (Ev21) Gene on the Z-Chromosome of Chickens. Poult. Sci. 1988, 67, 191–197. [Google Scholar] [CrossRef]
- Smith, E.J.; Fadly, A.M. Male-Mediated Venereal Transmission of Endogenous Avian-Leukosis Virus. Poult. Sci 1994, 73, 488–494. [Google Scholar] [CrossRef]
- Iraqi, F.; Smith, E.J. Determination of the Zygosity of Ev21-K in Late-Feathering Male White Leghorns Using the Polymerase Chain-Reaction. Poult. Sci. 1994, 73, 939–946. [Google Scholar] [CrossRef] [PubMed]
- Tixierboichard, M.H.; Benkel, B.F.; Chambers, J.R.; Gavora, J.S. Screening Chickens for Endogenous Virus Ev21 Viral Element by the Polymerase Chain-Reaction. Poult. Sci. 1994, 73, 1612–1616. [Google Scholar] [CrossRef] [PubMed]
- Iraqi, F.; Smith, E.J. Organization of the Sex-Linked Late-Feathering Haplotype in Chickens. Anim. Genet. 1995, 26, 141–146. [Google Scholar] [CrossRef] [PubMed]
- Kansaku, N.; Guemene, D.; Nakamura, A.; Uchida, M. Sequence Characterization of K-gene Linked Region in Various Chicken Breeds. J. Poult. Sci. 2011, 48, 181–186. [Google Scholar] [CrossRef] [Green Version]
- Smith, E.J.; Levin, I. Application of a locus-specific DNA hybridization probe in the analysis of the slow-feathering endogenous virus complex of chickens. Poult. Sci. 1991, 70, 1957–1964. [Google Scholar] [CrossRef] [PubMed]
- Boulliou, A.; Le Pennec, J.P.; Hubert, G.; Donal, R.; Smiley, M. The endogenous retroviral ev21 locus in commercial chicken lines and its relationship with the slow-feathering phenotype (K). Poult. Sci. 1992, 71, 38–46. [Google Scholar] [CrossRef]
- Takenouchi, A.; Toshishige, M.; Ito, N.; Tsudzuki, M. Endogenous viral gene ev21 is not responsible for the expression of late feathering in chickens. Poult. Sci. 2018, 97, 403–411. [Google Scholar] [CrossRef]
- Elferink, M.G.; Vallee, A.A.A.; Jungerius, A.P.; Crooijmans, R.P.M.A.; Groenen, M.A.M. Partial duplication of the PRLR and SPEF2 genes at the late feathering locus in chicken. BMC Genom. 2008, 9, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Bu, G.X.; Huang, G.; Fu, H.; Li, J.; Huang, S.M.; Wang, Y.J. Characterization of the novel duplicated PRLR gene at the late-feathering K locus in Lohmann chickens. J. Mol. Endocrinol. 2013, 51, 261–276. [Google Scholar] [CrossRef] [Green Version]
- Bu, G.X.; Wang, C.Y.; Cai, G.Q.; Leung, F.C.; Xu, M.; Wang, H.N.; Huang, G.; Li, J.; Wang, Y.J. Molecular characterization of prolactin receptor (cPRLR) gene in chickens: Gene structure, tissue expression, promoter analysis, and its interaction with chicken prolactin (cPRL) and prolactin-like protein (cPRL-L). Mol. Cell. Endocrinol. 2013, 370, 149–162. [Google Scholar] [CrossRef]
- Zhao, J.; Yao, J.; Li, F.; Yang, Z.; Sun, Z.; Qu, L.; Wang, K.; Su, Y.; Zhang, A.; Montgomery, S.A.; et al. Identification of candidate genes for chicken early- and late-feathering. Poult. Sci. 2016, 95, 1498–1503. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Zuo, F.; Zhang, W.; Wu, Y.; Liu, A. A Kind of Application and Method of PRLR Gene. CN. Patent CN107937568B, 30 June 2020. [Google Scholar]
- Li, J.; Liu, X.; Bao, H.; Wu, C. A Method of Identification Slow Feathering and Early Feathering Genotype. CN. Patent CN110129453B, 7 August 2020. [Google Scholar]
- Markoulatos, P.; Siafakas, N.; Moncany, M. Multiplex polymerase chain reaction: A practical approach. J. Clin. Lab. Anal. 2002, 16, 47–51. [Google Scholar] [CrossRef] [PubMed]
- Edwards, M.C.; Gibbs, R.A.J.G.R. Multiplex PCR: Advantages, development, and applications. Genome Res. 1994, 3, S65–S75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, Z.; Wang, J.; Zhang, S. Advances in multiple PCR technology studies. Chin. J. Biotechnol. 2020, 36, 171–179. [Google Scholar] [CrossRef]
- Zimmermann, K.; Schogl, D.; Plaimauer, B.; Mannhalter, J.W. Quantitative multiple competitive PCR of HIV-1 DNA in a single reaction tube. Biotechniques 1996, 21, 480–484. [Google Scholar] [CrossRef] [Green Version]
- Elnifro, E.M.; Ashshi, A.M.; Cooper, R.J.; Klapper, P.E. Multiplex PCR: Optimization and application in diagnostic virology. Clin. Microbiol. Rev. 2000, 13, 559–570. [Google Scholar] [CrossRef]
- Lee, G.-Y.; Kim, W.-K.; Cho, S.; Park, K.; Kim, J.; Lee, S.-H.; Lee, J.; Lee, Y.-S.; Kim, J.H.; Byun, K.S.J.M. Genotyping and Molecular Diagnosis of Hepatitis A Virus in Human Clinical Samples Using Multiplex PCR-Based Next-Generation Sequencing. Microorganisms 2022, 10, 100. [Google Scholar] [CrossRef]
- Henegariu, O.; Heerema, N.A.; Dlouhy, S.R.; Vance, G.H.; Vogt, P.H. Multiplex PCR: Critical parameters and step-by-step protocol. Biotechniques 1997, 23, 504–511. [Google Scholar] [CrossRef]
- Sint, D.; Raso, L.; Traugott, M. Advances in multiplex PCR: Balancing primer efficiencies and improving detection success. Methods Ecol. Evol. 2012, 3, 898–905. [Google Scholar] [CrossRef]
- Luo, C.L.; Shen, X.; Rao, Y.S.; Xu, H.P.; Tang, J.; Sun, L.; Nie, Q.H.; Zhang, X.Q. Differences of Z chromosome and genomic expression between early- and late-feathering chickens. Mol. Biol. Rep. 2012, 39, 6283–6288. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Breeds | Abbreviations | Phenotypes | Sex | Numbers of Chickens |
|---|---|---|---|---|
| Shouguang chicken | SG | LF | male | 48 |
| EF | male | 34 | ||
| LF | female | 76 | ||
| EF | female | 74 | ||
| Beijing You chicken | BY | LF | male | 59 |
| EF | male | 6 | ||
| Lvke chicken | LK | LF | male | 17 |
| Tibetan chicken | TB | LF | male | 27 |
| EF | male | 15 | ||
| Huxv chicken | HX | EF | male | 40 |
| Luxi gamecocks | LXG | EF | male | 16 |
| LF | male | 1 |
| Primers | Sequence(5′–3′) | Product Size |
|---|---|---|
| F1 | GTTTGACCTGTGCTGTGGTTTGCT | F1-R:386 bp |
| R | CTGTGCCCTTCCATCAGTGCTTC | |
| F2 | GCCATCAGCCAGATCCGTCAG | F2-R:786 bp |
| Breeds | Phenotypes | Sex | Numbers of Chickens | Numbers of Chickens Present of Tandem Duplication | Numbers of Chickens Absent of Tandem Duplication |
|---|---|---|---|---|---|
| SG | LF | male | 48 | 48 | |
| EF | male | 34 | 34 | ||
| LF | female | 76 | 76 | ||
| EF | female | 74 | 74 | ||
| BY | LF | male | 59 | 59 | |
| EF | male | 6 | 6 | ||
| LK | LF | male | 17 | 17 | |
| TB | LF | male | 27 | 27 | |
| EF | male | 15 | 15 | ||
| HX | EF | male | 40 | 40 | |
| LXG | EF | male | 16 | 16 | |
| LF | male | 1 | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shen, Q.; Li, J.; Bao, H.; Wu, C. Identification of Duplication Genotypes of the Feathering Rate Gene in Chicken by a Multiplex PCR Following Electrophoresis and/or Sanger Sequencing. Animals 2023, 13, 1091. https://doi.org/10.3390/ani13061091
Shen Q, Li J, Bao H, Wu C. Identification of Duplication Genotypes of the Feathering Rate Gene in Chicken by a Multiplex PCR Following Electrophoresis and/or Sanger Sequencing. Animals. 2023; 13(6):1091. https://doi.org/10.3390/ani13061091
Chicago/Turabian StyleShen, Qingmiao, Junying Li, Haigang Bao, and Changxin Wu. 2023. "Identification of Duplication Genotypes of the Feathering Rate Gene in Chicken by a Multiplex PCR Following Electrophoresis and/or Sanger Sequencing" Animals 13, no. 6: 1091. https://doi.org/10.3390/ani13061091