1. Introduction
Pearl gentian grouper (
Epinephelus fuscoguttatus ♀ ×
Epinephelus lanceolatus ♂), with the heterosis of fast growth and strong disease resistance, is farmed in large quantities in China [
1]. It is a predatory fish which requires about 45% fishmeal in the diet [
2]. Fishmeal is an expensive ingredient with limited supply. There have been many studies to assess the feasibility of replacing fishmeal with other ingredients [
3].
Single-cell protein generally refer to the biological proteins processed by microalgae, fungi, and bacteria, which high contents of crude protein and are rich in amino acids, various vitamins, trace elements, and bioactive substances [
4]. Single-cell protein can be produced from agricultural by-products, oil industry waste gas [
5], or through further processing of fermentation residues from the brewing and monosodium glutamate manufacturing industries [
6]. The full utilization of single-cell protein can lead to resource conservation, and in recent years, it has been widely studied as a promising alternative to fishmeal substitute in aquafeed [
7].
Clostridium autoethanogenum is a strictly anaerobic, gram-positive, rod-like, motile bacterium first reported in 1994 by Abrini et al. [
8]. It is capable of producing ethanol using CO as a carbon source and is therefore used to treat exhaust gases from the steel industry and to produce ethanol [
9]. The mash produced during this fermentation process is extracted and then processed to produce a protein ingredient, called
Clostridium autoethanogenum protein (CAP). CAP crude protein content is higher than 80%, amino acid composition is similar to fishmeal, and no anti-nutritional factors have been detected [
5]. Moreover, the results of genomic sequences showed that
Clostridium autoethanogenum had no virulence genes [
10], so CAP is considered a viable alternative to fishmeal. It is worth noting that CAP lacks crude fat compared to fishmeal [
11] and also lacks phosphorus when used in shrimp feed [
12], which needs to be compensated by additional supplementation. In previous studies, the proportion of CAP substituted fishmeal can reach 63% [
13], 50% [
14,
15], or 42.8% [
16] in
Micropterus salmoides, 58.0% in
Acanthopagrus schlegelii [
5], 30% in
Larimichthys crocea [
17], and 30% in
Litopenaeus vannamei [
18], while higher levels of substitution would significantly reduce the growth and had varying impacts on the liver or intestinal health of aquatic animals. In one of the studies on
M. salmoides, it was shown that 75% CAP substitution level causes swelling of hepatocytes [
13], while according to another study, more than 57.14% CAP substitution levels caused morphological atrophy of the intestine in
M. salmoides. CAP substitution levels above 60% also cause upregulation of
tumor necrosis factor α (
tnf-α) and downregulation of
interleukin 10 (
il-10) in the intestine of
L. crocea [
17].
Up to now, the application of CAP as a kind of ingredient for grouper has not been reported. This study aimed to investigate the feasibility and suitable substitution amount of fishmeal in the feed of pearl gentian grouper by replacing it with CAP. In addition to growth performance, physiological and biochemical indices, gene expression, and morphology of the tissues of grouper were monitored and evaluated comprehensively in this study.
2. Materials and Methods
2.1. Diets and Experimental Design
The control group feed was made with gluten (wheat, crude protein: 73.75%; crude fat: 0.11%), soybean meal (crude protein: 68.21%; crude fat: 9.00%), brown fishmeal (crude protein: 68.21%, crude fat: 9.00%) as protein ingredients and wheat flour (crude protein: 9.19%, crude fat: 0.38%) as the main binder, which was named FM. Four experimental diets were formulated to be isonitrogenous and isolipidic with 15%, 30%, 45%, and 60% fishmeal replaced by CAP (crude protein: 84.14%, crude fat: 0.19%, produced by Shoulang Biotechnology Co., Ltd. Beijing, China), named CAP15, CAP30, CAP45 and CAP60, respectively. Methionine and arginine were added in the experimental diets to the same level as the control group. The nutritional composition of CAP and fishmeal is shown in
supplementary material Table S1.
Except for choline chloride, all ingredients were thoroughly mixed after squeezing through a 60-mesh sieve and then mechanically mixed with choline chloride and water. The 2.5 mm diameter pellets were extruded using a twin-screw extruder (F-26, South China University of Technology, Guangzhou, China) then air-dried for 48 h, and finally, stored at −20 °C before use. The formulation and proximate composition of the experimental diets are shown in
Table 1.
2.2. Experimental Animal and Feeding Management
The experiment was carried out in the Zhanjiang Hi-Tech Park of Ocean (Zhanjiang, China). The juvenile pearl gentian grouper for the experiment were obtained from a local breeding factory (Zhanjiang, China). A total of 500 fish (18.01 ± 0.82 g) were randomly distributed into 20 tanks (0.3 m3) at a density of 25 fish per tank, and each diet was assigned to four tanks. Fish were fed to satiation at 8:00 and 17:00 each day. Continuous aeration was provided to each tank. The water was changed once a day, the temperature was 26 to 30 °C, and the salinity was 24 to 32%. Dead fish were weighed and the mortalities were recorded. The feeding trial lasted for 8 weeks.
2.3. Sample Collection
Fish from each tank were starved for 24 h, then counted and weighed. Three fish per tank were randomly selected to record body weight, visceral weight, and hepatic weight to calculate the VSI and HSI. Six fish per tank were randomly selected for blood sampling from the caudal vein. The blood samples were kept in the ice box. The front positions of liver and part of the distal intestines taken from two fish per tank were cut off and stored in 4% formaldehyde solution for hematoxylin-eosin (H&E). For FM, CAP30, and CAP60 groups, the other part of the distal intestines was cut to 1 mm in size, and stored in 2.5% glutaraldehyde solution for transmission electron microscopy (TEM) samples preparation. The liver, dorsal muscle, and distal intestine samples from four fish per tank were collected, immediately frozen in liquid nitrogen, and then stored at −80 °C for analysis of the biochemical indices, enzyme activities, and gene expression. Three fish were taken from each tank and used for proximate composition assay.
2.4. Proximate Composition Assay
Proximate composition of experimental diets and whole fish samples were measured. Moisture was analyzed by drying the samples to constant weight at 105 °C. Crude lipid was measured by petroleum ether using the Soxhlet method. Crude protein was determined using the Dumas Nitrogen method by Primacs 100 analyzer (Skalar, Dutch) and estimated by multiplying nitrogen by 6.25.
2.5. Biochemical Indices and Enzyme Activities Assay
After 12 h of sampling, the blood samples were centrifuged (3500 rpm, 15 min, 4 °C), then serum was obtained. About 200 mg of tissue was weighed and then homogenized in 9:1 (saline solution: tissue) saline solution with ice bath. After centrifugation (2500 rpm, 10 min, 4 °C), supernatants were collected.
For serum samples, the contents of total protein (TP), triglyceride (TG), uric acid (UA), total cholesterol (T-CHO), malondialdehyde (MDA), glutathione (GSH), the activities of acid phosphatase (ACP), alanine aminotransferase (ALT), aspartate aminotransferase (AST), as well as total antioxidant capacity (T-AOC) were measured. For the treated liver samples, the contents of MDA, GSH, the activities of ACP, superoxide dismutase (SOD) and lysozyme, as well as total antioxidant capacity (T-AOC) were measured. For the treated distal intestine samples, the activities of trypsin, SOD, and lysozyme were measured.
Among the above indices, except for the trypsin, SOD and lysozyme were determined by enzyme-linked immunosorbent assay (ELISA) detection kits (Shanghai Enzyme-linked Biotechnology, Shanghai, China), following the instructions. All the other indices were determined by detection kits (Nanjing Jiancheng Bioengineering Institute, Nanjing, China), following the instructions.
2.6. Tissue Morphological Histology
According to the method of Chen et al. [
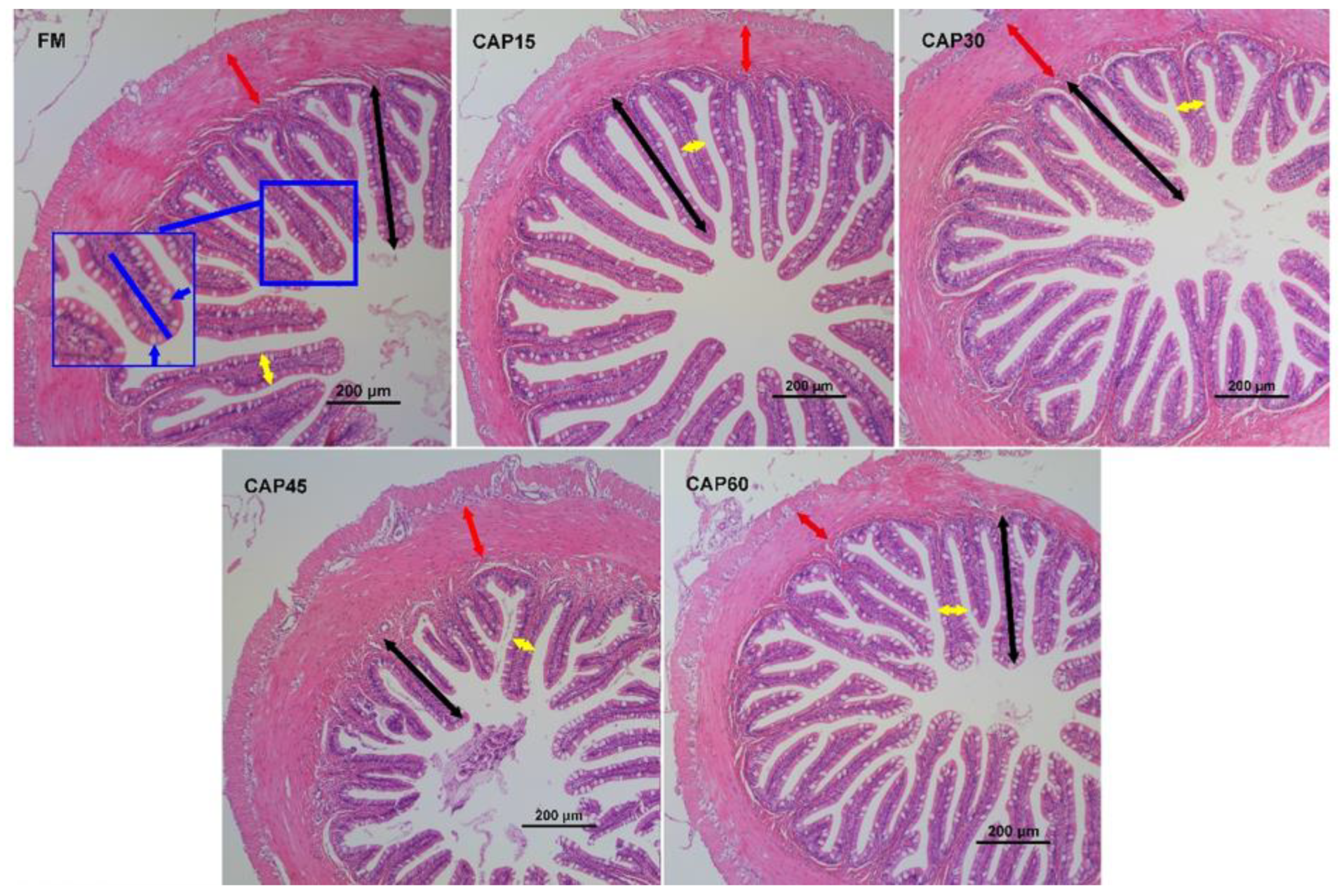
19], the samples for H&E stain were fixed with paraffin and sectioned, dewaxed, and stained with Hematoxylin solution, then were dehydrated and sealed with neutral gum. The histology of samples was photographed using an optical microscope (Nikon Ni-U, Japan). For the hepatic sections, nuclear excursion, inflammatory, and ballooning degenerated hepatocytes were assessed. For the distal intestines sections, ten mucosal folds were randomly selected, the height and width were measured, and the number of goblet cells within 200 µm from the top was counted. Ten measurement points were selected at equal intervals to measure the thickness of the muscularis.
The electron microscopy samples were prepared and observed with reference to the method of Huang et al. [
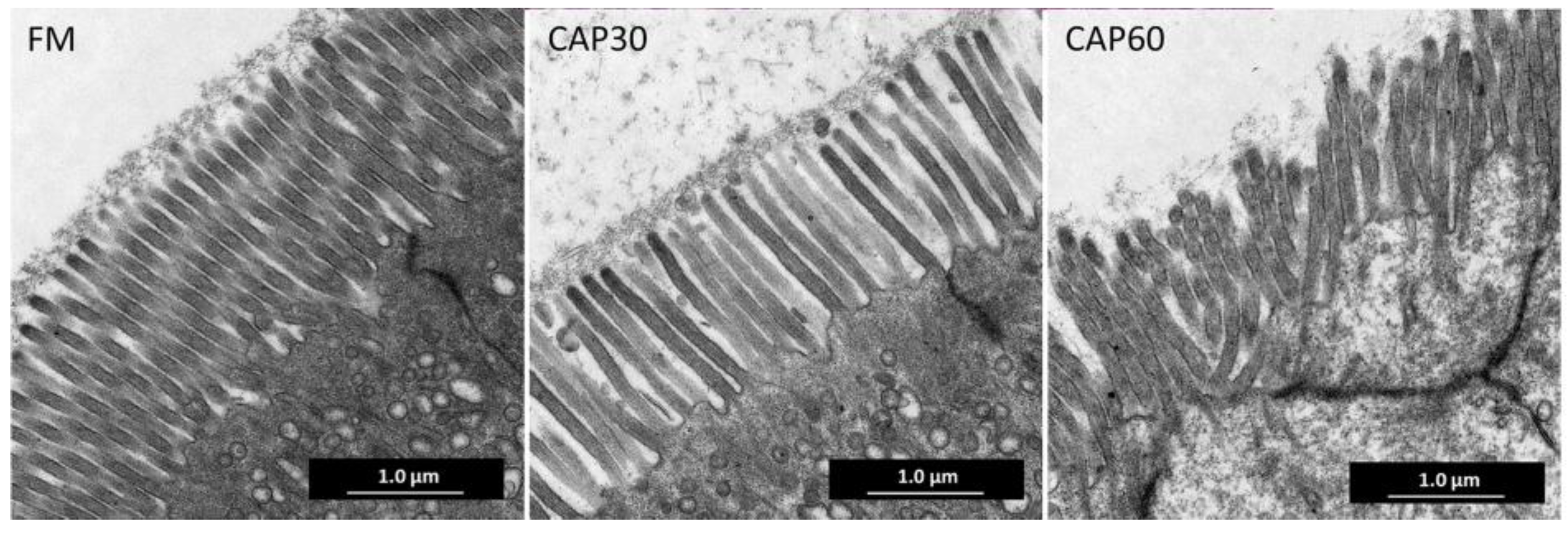
20]. Briefly, tissue samples were fixed in 2% osmium tetroxide phosphate buffer, then dehydrated in ethanol, an embedded in epoxy resin before being cut into ultrathin sections by an ultramicrotome (Leica EM UC7, Japan). Ultrathin sections were dyed by uranyl acetate and lead citrate, and photographed using a transmission electron microscope (HITACHI HT7600, Japan) at an accelerating voltage of 80 kV. Thirty intestinal microvillus were randomly selected in each sample to measure the length, and the number of microvillus within 10 μm was counted to calculate the density.
2.7. Real-Time Quantitative PCR Assay
Total RNA was extracted from tissue using an RNA extraction kit (Transgen Biotech, Beijing, China), and the concentration was detected with a NanoDrop 2000 (Thermo Fisher Scientific, Waltham, MA, USA) and adjusted to 600 ng μL−1 by adding ultrapure water. Evo M-MLV RT kit (Accurate Biology, Changsha, China) was used to synthesize cDNA. The methods and reactions followed the instructions.
The amplification of real-time quantitative PCR was performed in a total volume of 10 μL, comprising 0.1 μL (5μM) of each primer, 1 μL of cDNA, 3.8 μL of sterilized double-distilled water, and 5 μL of 2X SYBR
® Green Pro Taq HS PremixII (Accurate Biotechnology, China), using a quantitative thermal cycler (Light Cycler 480, Roche Diagnostics, Switzerland). The target gene expression was calculated by the 2
−∆∆Ct method [
21] using
β-
actin as the reference gene [
22] and the FM group as the reference group. The information of primers was presented in
Table 2. Some of these primers were derived from the studies of Xu et al. [
23] and Yin et al. [
24], which were designed according to the full-length sequences from the sequenced transcriptome of the pearl gentian grouper (not published).
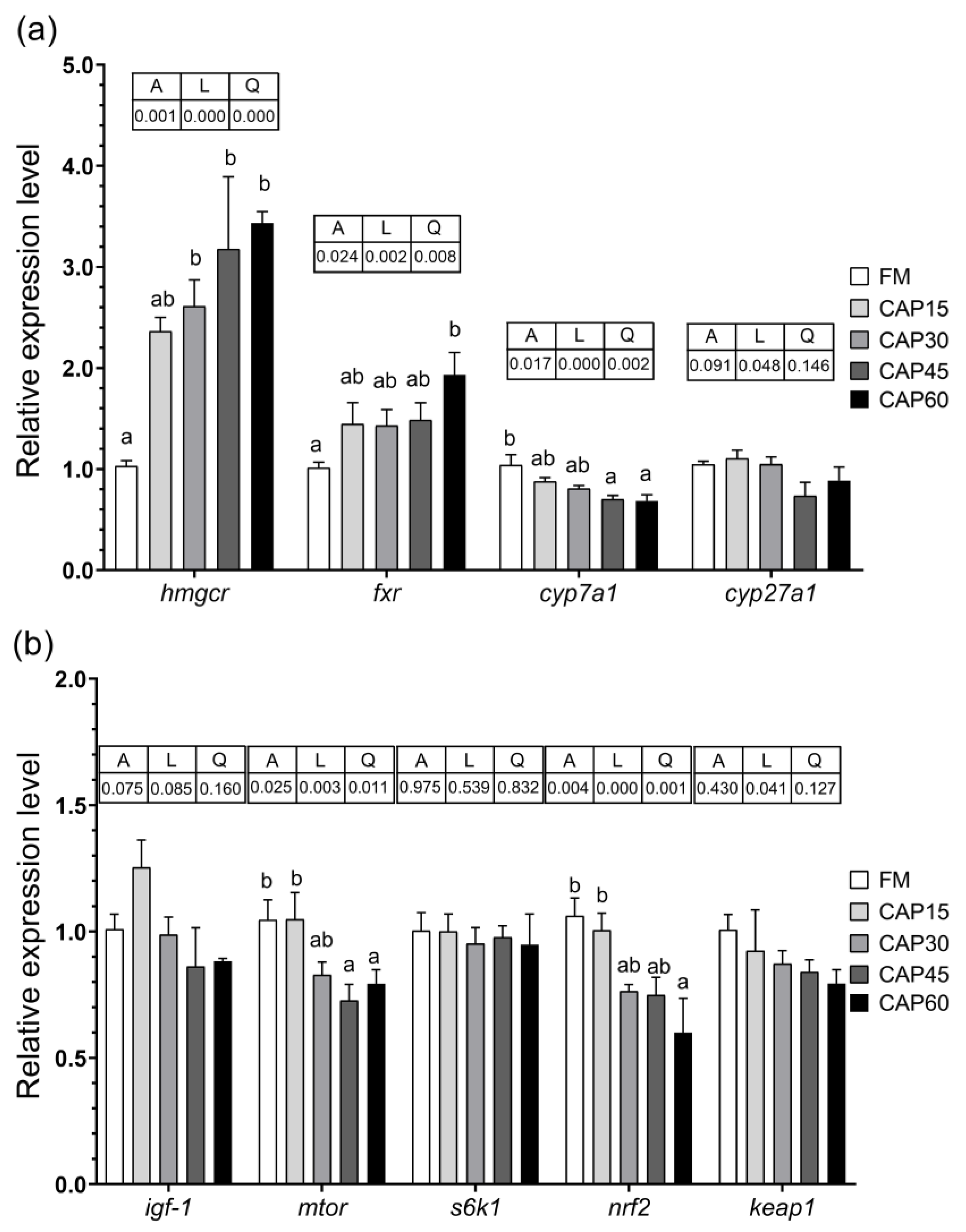
For the aspect of cholesterol bile acid metabolism, four genes, 3-hydroxy-3-methylglutaryl coenzyme A reductase (hmgcr), farnesoid X (fxr) and cholesterol 7α-hydroxylase (cyp7a1), cholesterol 27α-hydroxylase (cyp27a1) were selected. For the IGF-1/PI3K/AKT/mTOR signaling pathway, six genes, insulin-like growth factor (igf-1), serine/threonine-protein kinase (akt), mammalian target of rapamycin (mtor), eukaryotic translation initiation factor 4E-binding protein 1 (4e-bp1), and ribosomal protein S6 kinase 1 (s6k1) were selected. For the Nrf2-Keap1 signaling pathway, nuclear erythroid 2-related factor 2 (nrf2) and kelch-like ECH associating protein 1 (keap1) were selected.
2.8. Calculations and statistical analysis
The calculation formulas were as follows:
Weight gain (WG,%) = 100 × (Wt − Wi)/Wi;
Specific growth rate (SGR,%) = 100 × (lnWt − lnWi)/t;
Survival rate (SR,%) = 100 × Nt/Ni;
Feed conversion ratio (FCR) = Wfi/(Wt − Wi)
Daily Feed intake ratio (DFI,%/day) = 100 × Wfi/((Wg − Wi)/2 × t)
Condition factor (CF, kg/cm3 %) = 100 × Body weight (g)/Body length (cm3)
Hepatosomatic index (HSI,%) = 100 × Hepatosomatic weight (g)/Body weight (g)
Viscerasomatic index (VSI,%) = 100 × Viscerasomatic weight (g)/Body weight (g)
where Wt is final body weight (g); Wi, initial body weight (g); Wfi, Total feed intake (g); Nt, the final fish number; Ni, the initial fish number; t, the duration of experiment days.
All data were subjected to statistical verification using one-way analysis of variance (ANOVA) with SPSS 25.0 (International Business Machines Corp, Armonk, NY, USA). When there were significant differences between groups, Tukey’s multiple comparison test was performed with differences significant at
p < 0.05. Further, a follow-up trend analysis was performed using orthogonal polynomial comparison, to determine whether the effects were linear or quadratic [
17].
4. Discussion
The results of this study indicate that CAP is a feasible alternative to fishmeal in the feed of pearl gentian grouper. Even a 45% replacement level does not significantly affect the growth of grouper. Similar results have been shown in other studies of marine fish. In a study on
A. schlegelii, Chen et al. [
5] found that CAP could replace 58% of fishmeal without significantly affecting growth indices, while in a study on
L. crocea, Wu et al. [
17] found that CAP could replace 30% of fishmeal in the feed. The studies on
A. schlegelii have found that replacing fishmeal in feed with high levels of CAP reduced the palatability of the feed [
5]. In this trial, the mean value of DFI in the CAP30 group was even higher than that in the control group, which may indicate that feeds with appropriate CAP substitution levels have better feeding attraction. However, when 45% or more fishmeal was replaced in the diet, the DFI of grouper was reduced compared with the control group, but the difference was not significant, indicating that the palatability of CAP is still relatively suitable for grouper. Therefore, the significant reduction of growth performance in the CAP60 group may be influenced by other factors.
ACP is a hydrolase that destroys the structure of pathogens by hydrolyzing phosphate esters to enhance immunity [
25]. In this experiment, the activity of ACP in serum and liver increased with the increasing dietary CAP levels, which was contrary to the findings of Wu et al. on
L. crocea [
17], probably due to the difference of species. Another study indicated that ACP activity within the serum and liver of hybrid grouper was increased by negative stress [
26], suggesting that the effects of excessive levels in the diet of CAP on grouper may negatively affect the immunity of grouper.
The serum activities of AST and ALT are low in normal conditions, but when liver damage occurs, these two transaminases escape from the hepatocytes, causing a high level of their activities in the serum [
27]. GSH is an important antioxidant substance in cells and is essential for maintaining the stability of the intracellular environment [
28]. Therefore, when lipid peroxidation increases, the GSH content decreases, in addition to an increase in MDA content [
29]. In this experiment, the change of activity of two transaminases, the contents of MDA, GSH in the serum and the changes of MDA, GSH, T-AOC in the liver illustrated the oxidative damage to the organism by excessive substitution of fishmeal by CAP. In a study on
M. salmoides, Yang et al. [
16] also found that fishmeal in CAP replacement feed significantly increased serum MDA levels, similar to this experiment. However, in the study of
A. schlegelii, 58% CAP substitution levels in diet did not significantly affect the antioxidant capacity of the liver [
5]. It is possible that these results are caused by differences in the experimental species.
The Nrf2-Keap1 signaling pathway is an important endogenous signaling pathway affecting the resistance to oxidative stress [
30]. Among them,
nrf2 is a transcription factor that can upregulate the transcription of antioxidant-related genes to maintain cellular defense [
31]. When the intracellular oxidative damage situation normalizes,
keap1 expression is upregulated and Keap1 enters the nucleus to bind with Nrf2 and exit the nucleus, returning to its steady-state [
32]. In studies on
Sillago sihama [
33] and
Pseudosciaena crocea [
34],
keap1 expression level in the liver was positively correlated with
nrf2. Similar results were presented in this study, and the changes in the expression of these two genes also suggested that the level of CAP substitution to fishmeal in the feed affected the antioxidant capacity of the grouper liver.
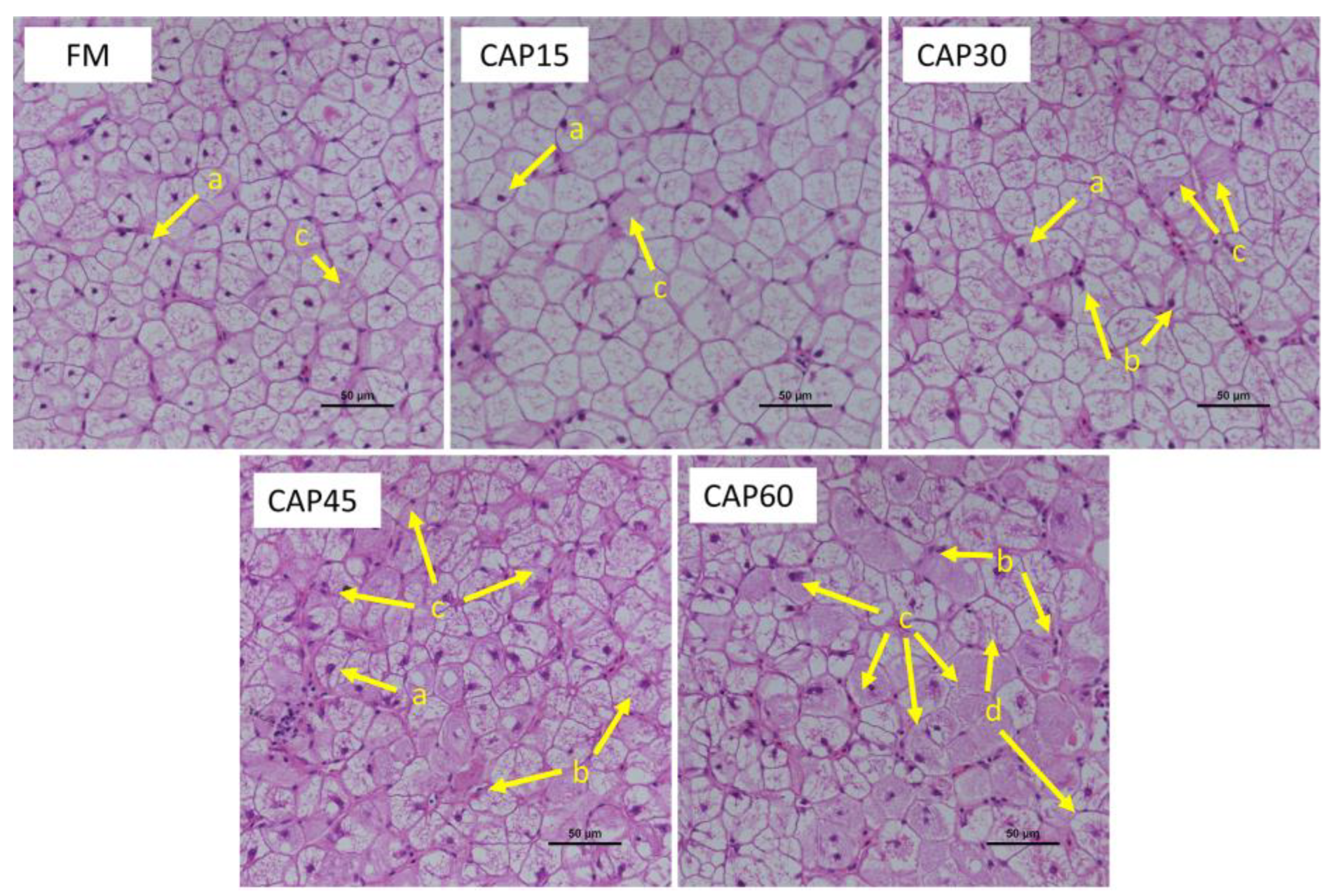
In addition to changes in biochemical indicators, the morphological structure of liver cells can be significantly influenced by diet [
35]. In histopathology, the position of nuclei can reflect the health of the hepatocytes [
36]. The ground-glass microstructure of the hepatocytes indicates hypertrophy of the smooth endoplasmic reticulum, a sign of inflammation [
37], while ballooning degeneration of the hepatocytes is often considered a form of apoptosis [
38]. This result revealed that substitution of fishmeal in feed with high level CAP induced increased hepatocytes inflammation.
Cholesterol is an important component of cell membranes and a precursor of bile acids and steroid hormone. Since fish can synthesize cholesterol, it is usually considered a non-essential nutrient [
39]. Liver is the principal site of cholesterol synthesis, as well as the place of producing bile acids by cholesterol, where HMGCR and CYP7A1, CP27A1 are the key enzymes involved in cholesterol synthesis and bile acid production, respectively [
40]. The farnesol X receptor (FXR) is the bile acid receptor that inhibits the expression of
cyp7a1 to negatively regulate the production of bile acids [
23]. Although we designed the feed formulation with fish oil to equalize the crude fat in each group, the results showed that the CAP high level replacement group showed differences in cholesterol metabolism compared to the FM group. According to the study by Wu et al. [
41], the addition of cholesterol in the diet of giant grouper (
Epinephelus lanceolatus) resulted in higher blood cholesterol content than the control group, but the expression of
hmgcr was downregulated as a result. Similarly, in this study with increasing dietary CAP replacement level, the expression level of
hmgcr was significantly upregulated, while the serum T-CHO content in CAP45 and CAP60 groups was significantly lower than the FM group. Furthermore, the change in the relative expression of
fxr,
cyp7a1, and
cyp27a1 indicates that bile acid synthesis is inhibited when high level of fishmeal was replaced by CAP, which may be due to the deficiency of cholesterol. However, more in-depth verification is needed to further investigated.
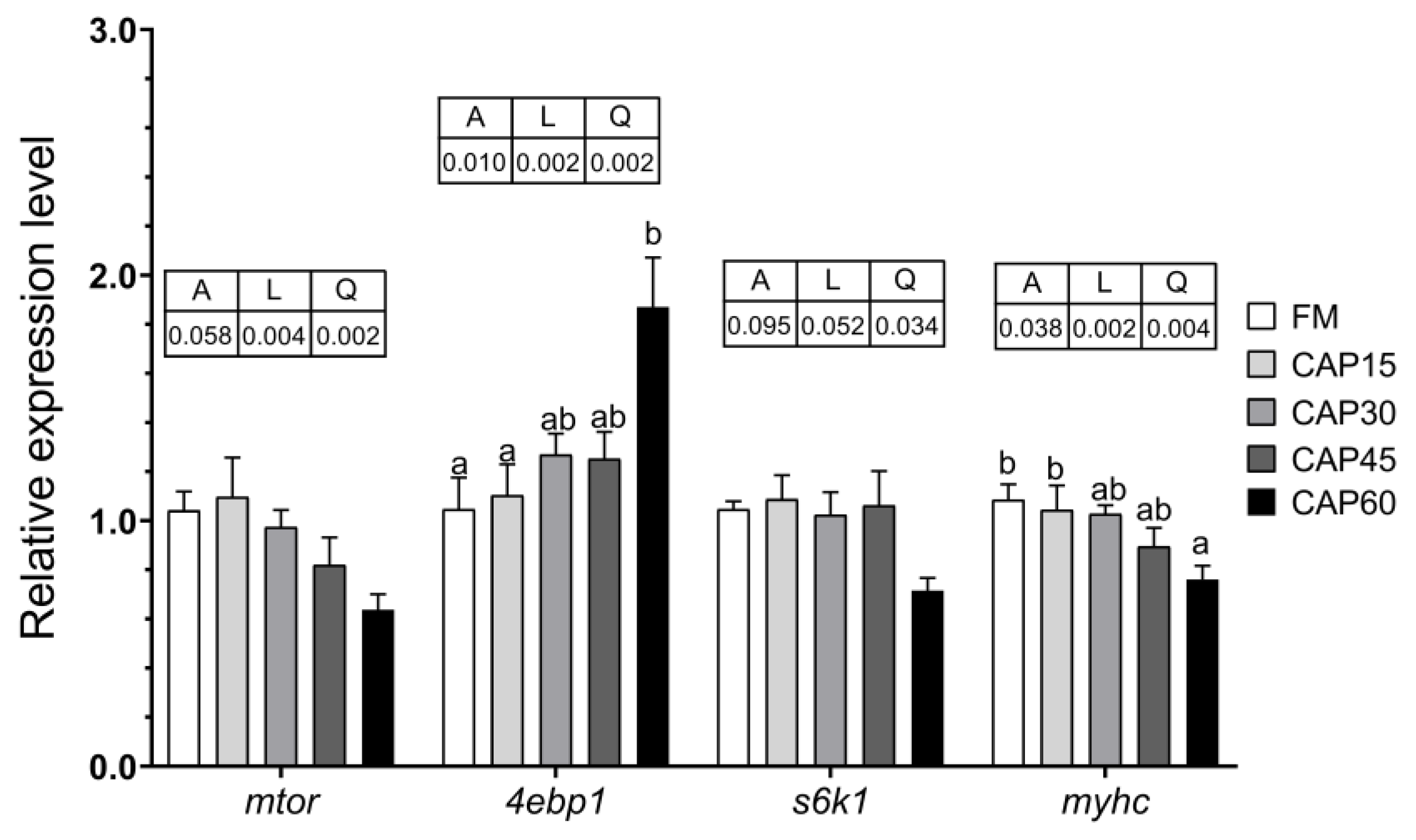
The mTOR is the signaling pathway involved in regulating the metabolism of nutrients such as proteins, lipids, and nucleic acids in the organism to preserve normal cell growth and proliferation [
24]. In the IGF-1/PI3K/AKT/mTOR signaling pathway, IGF-1 activates the PI3K/AKT pathway, which in turn activates the formation of mTOR, phosphorylates S6K1, and inhibits 4EBP1 activity [
42]. The phosphorylation of S6K1 promotes the initiation of translation, while 4E-BP1 is a negative regulator of the translation process [
43].
igf-1 is significantly highly expressed in fish liver [
44]. Although the expression of
igf-1 and
s6k1 in liver was not significantly affected in this experiment, the difference in
mtor still indicated that the substitution of CAP for fishmeal would still affect the IGF-1/PI3K/AKT/mTOR signaling pathway to some extent. It has been reported that replacing fishmeal with blended alternatives leads to down-regulation of the expression of
mtor,
s6k1 in grouper [
45], which is similar to the present study. In addition, the expression of these two genes in grouper is also affected by certain amino acid deficiencies [
46]. Subject to IGF-1 mediated effects, changes in the expression level of
myhc result in changes in the content of myosin [
44]. In a previous study on pearl gentian grouper, Yang et al. [
47] used its expression as an index to evaluate muscle protein synthesis capacity. In this study, the expression level of
mtor,
4ebp1,
s6k1, and
myhc in dorsal muscles all showed to varying degrees that CAP has a negative effect on muscle protein synthesis at higher levels of pair substitution. After all, CAP has not yet been found to contain active small peptides or unknown growth factors, which are nutrients that are abundant in fishmeal [
2].
The height of the intestinal mucosal folds, as well as the length and density of the microvilli determine the absorption surface area of the intestine together [
20,
48]. In this study, microvilli in the CAP60 group were disorganized, sparse, and short; intestinal folds and muscle layer thickness were also decreased with the increasing CAP levels, similar to the results of studies in
M. salmoides [
16], reflects the damage of the intestinal. In addition, this result of decreased trypsin activity further suggests that the absorption capacity of the intestine would be affected. Based on the results of
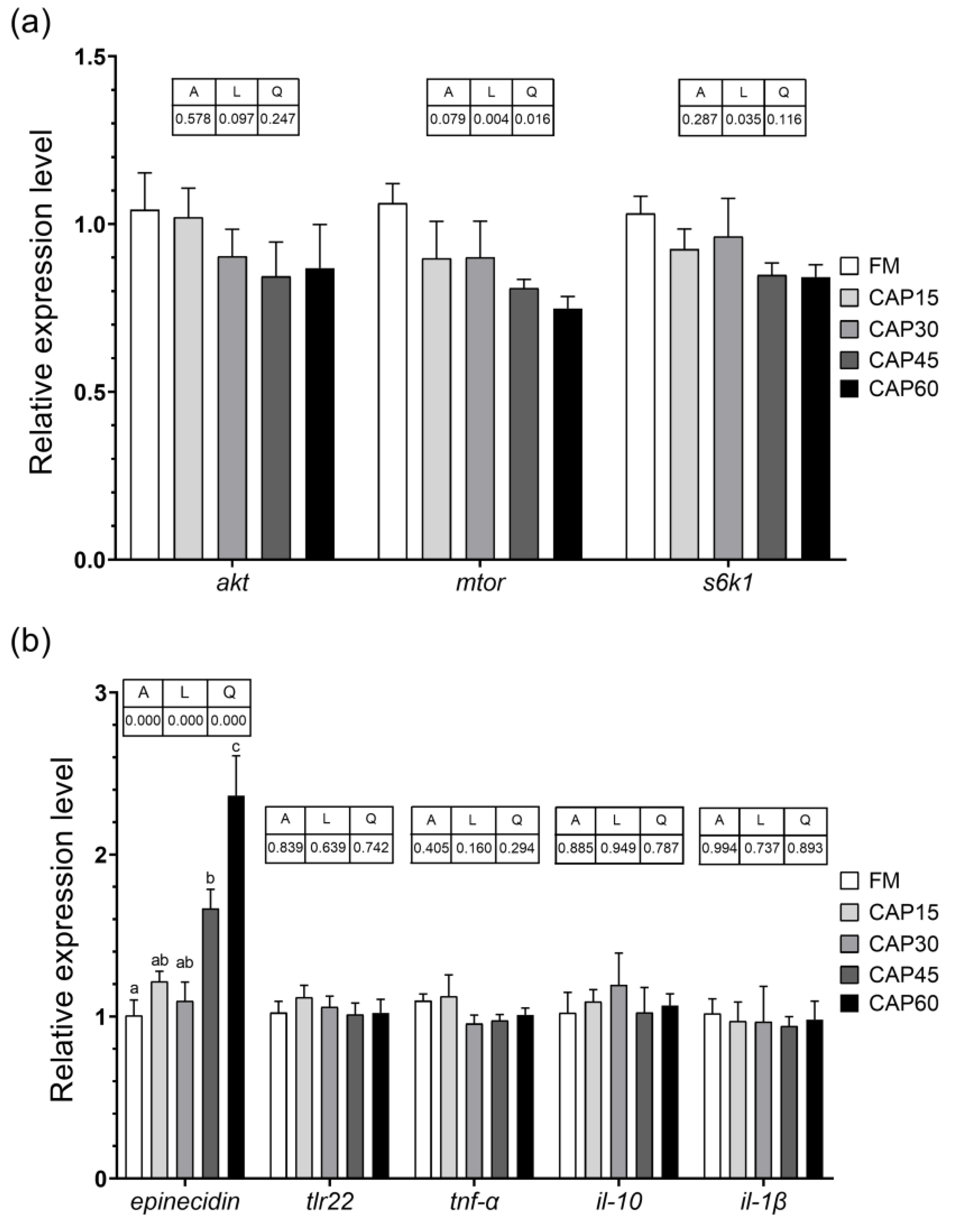
akt,
mtor, and
s6k1 gene expression assays, it can be further inferred that high levels of CAP substitution have a negative impact on intestinal development. Deficiencies in intestinal digestive and absorptive capacity were important in affecting growth performance in this experiment.
The mucus layer of the intestine has a barrier role in protecting the intestine, and its main component, mucoglycoprotein, is secreted by the goblet cells [
49]. Goblet cells are highly differentiated columnar epithelial cells [
50] that appear vacuolated after HE staining treatment. In this study, the high number of goblet cells in the CAP15 group versus the control group suggests that there is some improvement in intestinal barrier function by CAP, but further, the high levels of substitution can diminish intestinal barrier function. It has also been reported that exposure to oxidative damage leads to a reduction of intestinal goblet cells in pearl gentian grouper [
51]. In this study, the activities of SOD were decreased in the high-level CAP substitution group. All these results suggest that a diet with high levels of CAP substitution for fishmeal also causes some degree of oxidative damage to the distal intestines of pearl gentian grouper. Epinecidin has the ability to kill microorganisms directly or inhibit their growth [
52]. In previous studies on grouper, it has been found that the expression level of epinecidin-related genes in the distal intestine can be affected by the vitamin and probiotic preparations added to the feed [
53,
54]. Combined with the result that the relative expression of
epinecidin mRNA in distal intestine was upregulated with the increase in feed CAP substitution in this experiment, it is hypothesized that CAP, as a single-cell protein, contains unknown factors that stimulate the distal intestine leading to an increase in epinecidin expression, the mechanism of which needs to be further investigated.









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}