
Luteolin: A Phytochemical to Mitigate S. Typhimurium Flagellin-Induced Inflammation in a Chicken In Vitro Hepatic Model
 , , , , , and
, , , , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell isolation and Establishment of Cell Cultures
2.2. Treatment of Cell Cultures
2.3. Measurements
2.3.1. Metabolic Activity
2.3.2. LDH Activity
2.3.3. IFN-α, IFN-γ, IL-10 Concentration
2.3.4. IL-6 and IL-8 Concentration
2.3.5. H2O2 Level
2.3.6. Malondialdehyde Concentration
2.4. Statistical Analysis
3. Results
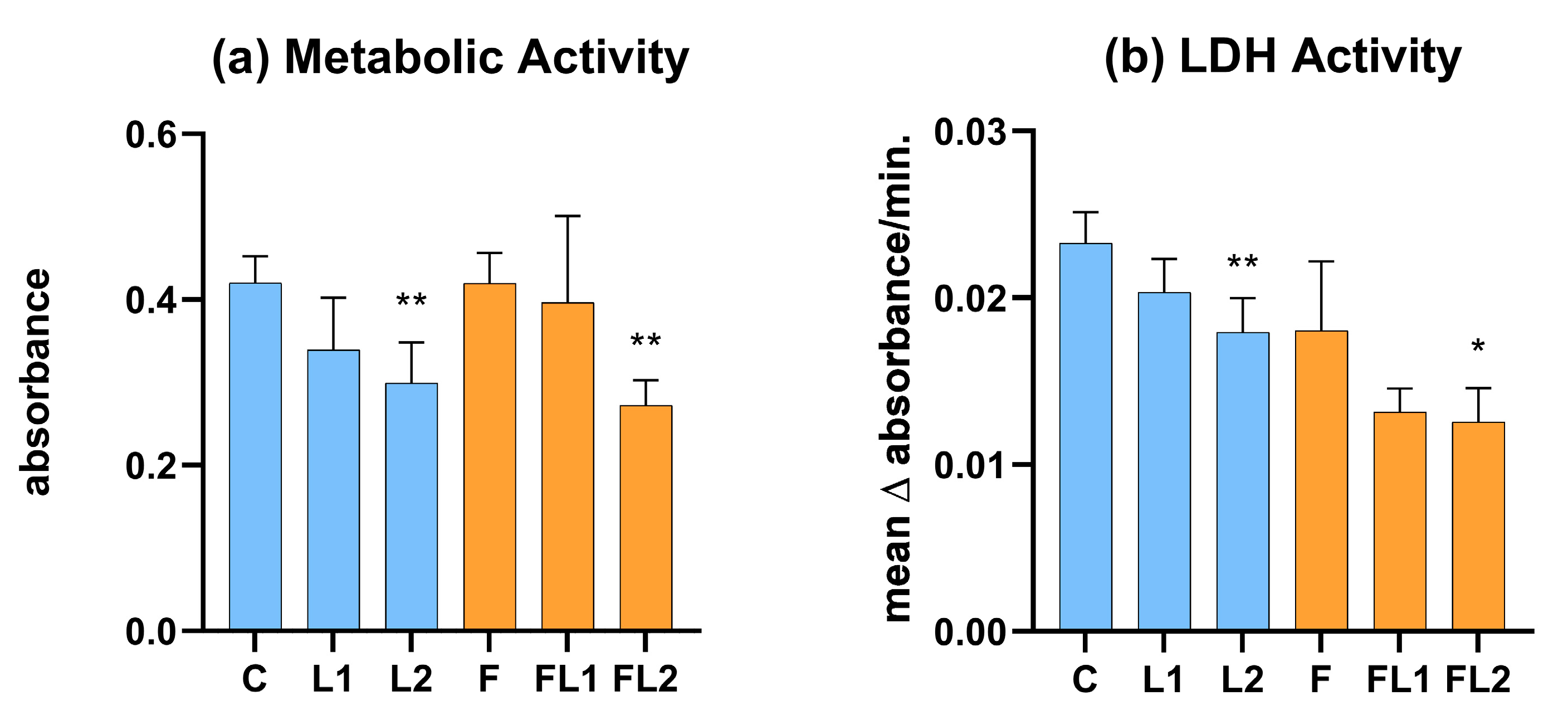
3.1. Metabolic Activity and LDH Activity
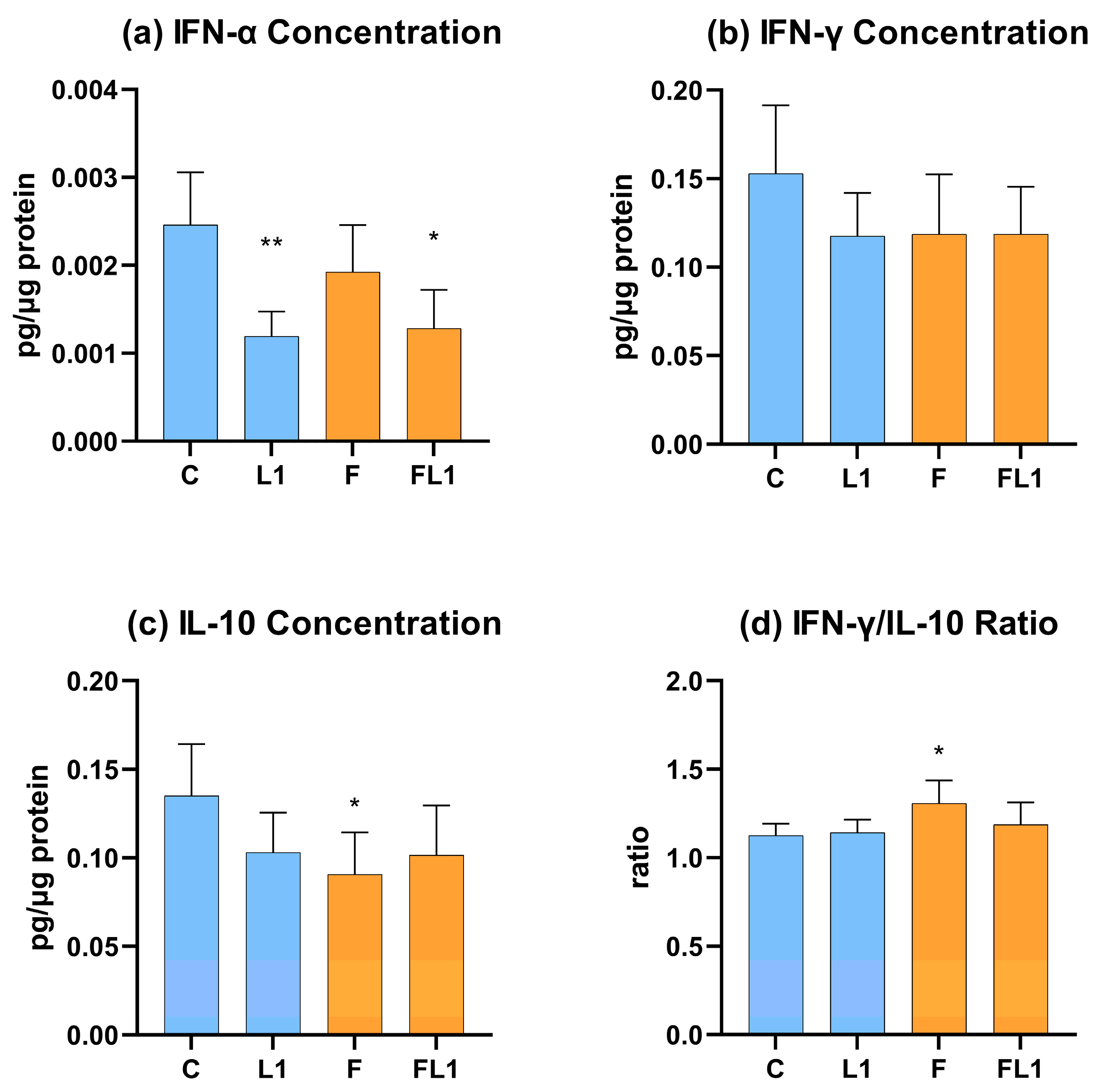
3.2. IFN-α, IFN-γ, IL-10 Concentration and IFN-γ/IL-10 Ratio
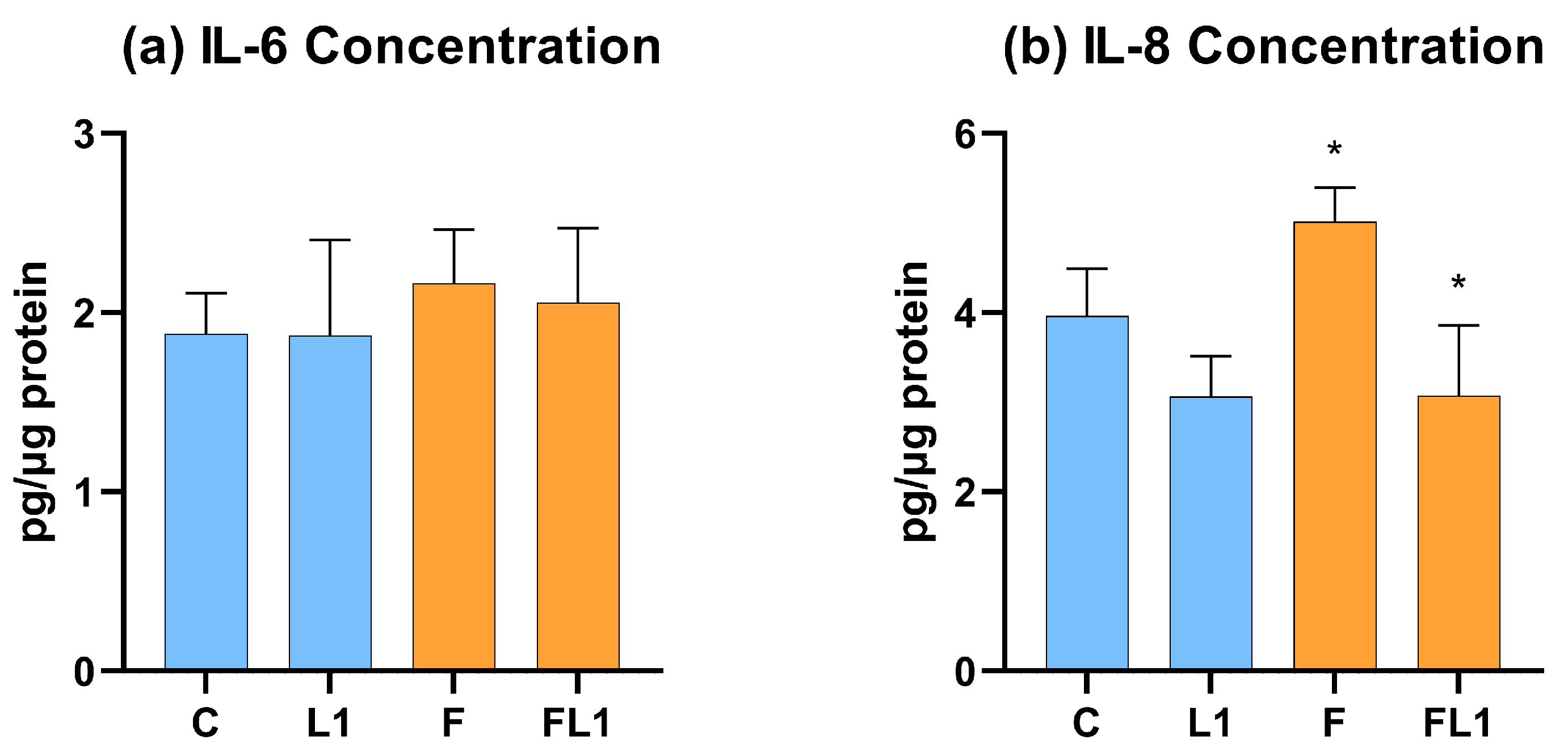
3.3. IL-6 and IL-8 Concentration
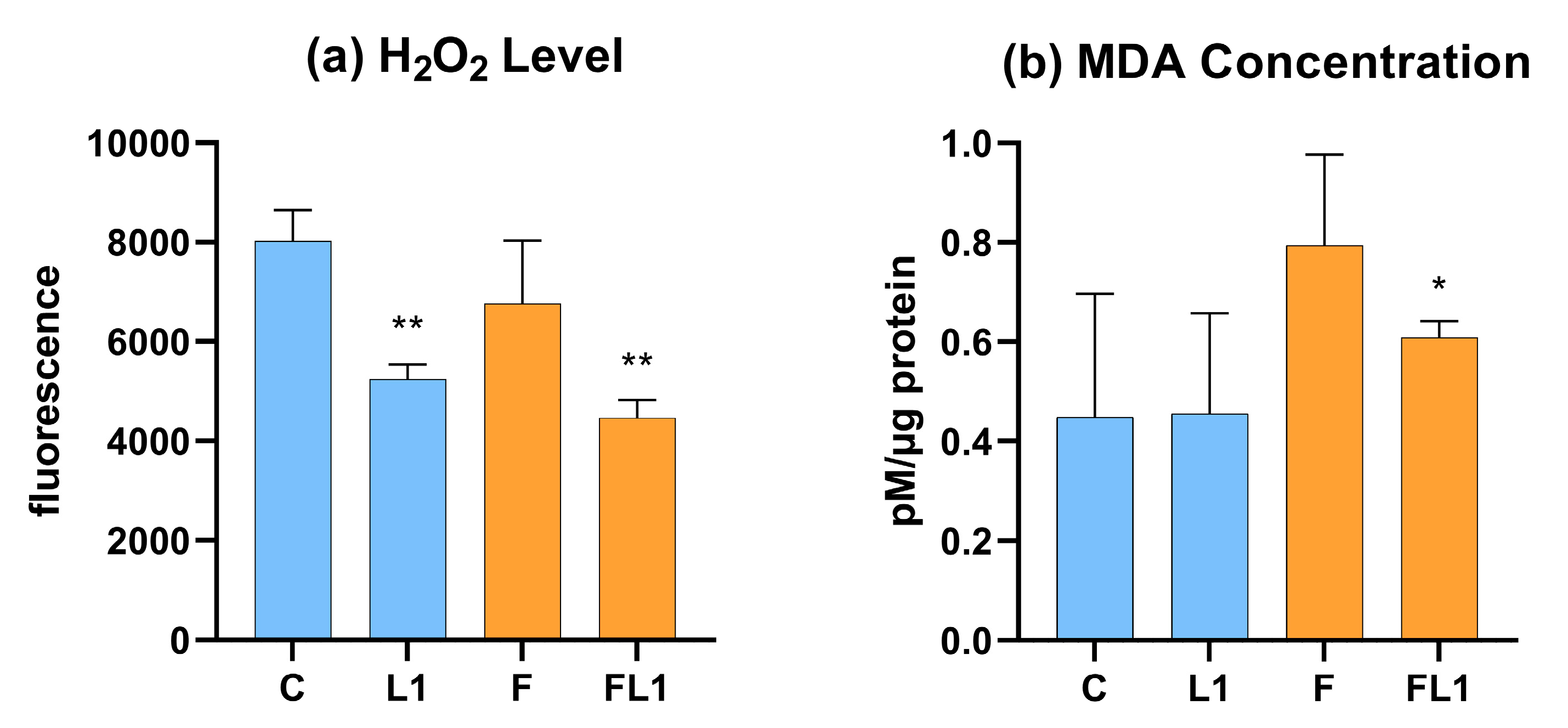
3.4. H2O2 Level and Malondialdehyde Concentration
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gao, B.; Jeong, W.-I.; Tian, Z. Liver: An Organ with Predominant Innate Immunity. Hepatology 2008, 47, 729–736. [Google Scholar] [CrossRef]
- Ishibashi, H.; Nakamura, M.; Komori, A.; Migita, K.; Shimoda, S. Liver Architecture, Cell Function, and Disease. Semin. Immunopathol. 2009, 31, 399. [Google Scholar] [CrossRef] [PubMed]
- Hendriksen, R.S.; Vieira, A.R.; Karlsmose, S.; Lo Fo Wong, D.M.A.; Jensen, A.B.; Wegener, H.C.; Aarestrup, F.M. Global Monitoring of Salmonella Serovar Distribution from the World Health Organization Global Foodborne Infections Network Country Data Bank: Results of Quality Assured Laboratories from 2001 to 2007. Foodborne Pathog. Dis. 2011, 8, 887–900. [Google Scholar] [CrossRef]
- Beal, R.K.; Wigley, P.; Powers, C.; Hulme, S.D.; Barrow, P.A.; Smith, A.L. Age at Primary Infection with Salmonella enterica Serovar Typhimurium in the Chicken Influences Persistence of Infection and Subsequent Immunity to Re-Challenge. Vet. Immunol. Immunopathol. 2004, 100, 151–164. [Google Scholar] [CrossRef]
- Hugas, M.; Beloeil, P.A. Controlling Salmonella along the Food Chain in the European Union—Progress over the Last Ten Years. Eurosurveillance 2014, 19, 20804. [Google Scholar] [CrossRef]
- Barrow, P.A. The Paratyphoid Salmonellae. Rev. Sci. Tech. Int. Off. Epizoot. 2000, 19, 351–375. [Google Scholar] [CrossRef] [PubMed]
- Barrow, P.A.; Huggins, M.B.; Lovell, M.A.; Simpson, J.M. Observations on the Pathogenesis of Experimental Salmonella Typhimurium Infection in Chickens. Res. Vet. Sci. 1987, 42, 194–199. [Google Scholar] [CrossRef] [PubMed]
- Elkenany, R.M.; Eladl, A.H.; El-Shafei, R.A. Genetic Characterisation of Class 1 Integrons among Multidrug-Resistant Salmonella Serotypes in Broiler Chicken Farms. J. Glob. Antimicrob. Resist. 2018, 14, 202–208. [Google Scholar] [CrossRef]
- Bailey, M.; Taylor, R.; Brar, J.; Corkran, S.; Velásquez, C.; Novoa-Rama, E.; Oliver, H.F.; Singh, M. Prevalence and Antimicrobial Resistance of Salmonella from Antibiotic-Free Broilers During Organic and Conventional Processing. J. Food Prot. 2020, 83, 491–496. [Google Scholar] [CrossRef]
- Melendez, S.; Hanning, I.; Han, J.; Nayak, R.; Clement, A.; Wooming, A.; Hererra, P.; Jones, F.t.; Foley, S.l.; Ricke, S.C. Salmonella enterica Isolates from Pasture-Raised Poultry Exhibit Antimicrobial Resistance and Class I Integrons. J. Appl. Microbiol. 2010, 109, 1957–1966. [Google Scholar] [CrossRef]
- Zeng, H.; Carlson, A.Q.; Guo, Y.; Yu, Y.; Collier-Hyams, L.S.; Madara, J.L.; Gewirtz, A.T.; Neish, A.S. Flagellin Is the Major Proinflammatory Determinant of Enteropathogenic Salmonella. J. Immunol. 2003, 171, 3668–3674. [Google Scholar] [CrossRef] [PubMed]
- Gewirtz, A.T.; Yu, Y.; Krishna, U.S.; Israel, D.A.; Lyons, S.L.; Peek, R.M. Helicobacter Pylori Flagellin Evades Toll-like Receptor 5-Mediated Innate Immunity. J. Infect. Dis. 2004, 189, 1914–1920. [Google Scholar] [CrossRef]
- Kogut, M.H.; Genovese, K.J.; He, H.; Kaiser, P. Flagellin and Lipopolysaccharide Up-Regulation of IL-6 and CXCLi2 Gene Expression in Chicken Heterophils Is Mediated by ERK1/2-Dependent Activation of AP-1 and NF-KappaB Signaling Pathways. Innate Immun. 2008, 14, 213–222. [Google Scholar] [CrossRef]
- Kogut, M.H.; Iqbal, M.; He, H.; Philbin, V.; Kaiser, P.; Smith, A. Expression and Function of Toll-like Receptors in Chicken Heterophils. Dev. Comp. Immunol. 2005, 29, 791–807. [Google Scholar] [CrossRef]
- Fliegmann, J.; Felix, G. Immunity: Flagellin Seen from All Sides. Nat. Plants 2016, 2, 16136. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, F.d.O.; Freitas Neto, O.C.d.; Batista, D.F.A.; Almeida, A.M.d.; Rubio, M.d.S.; Alves, L.B.R.; Vasconcelos, R.d.O.; Barrow, P.A.; Berchier, A., Jr. Contribution of Flagella and Motility to Gut Colonisation and Pathogenicity of Salmonella Enteritidis in the Chicken. Braz. J. Microbiol. Publ. Braz. Soc. Microbiol. 2017, 48, 754–759. [Google Scholar] [CrossRef]
- Akira, S.; Uematsu, S.; Takeuchi, O. Pathogen Recognition and Innate Immunity. Cell 2006, 124, 783–801. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, M.D. Endotoxin and Kupffer Cell Activation in Alcoholic Liver Disease. Alcohol Res. Health 2003, 27, 300–306. [Google Scholar]
- Kiziltas, S. Toll-like Receptors in Pathophysiology of Liver Diseases. World J. Hepatol. 2016, 8, 1354–1369. [Google Scholar] [CrossRef]
- Jacobs, M.D.; Harrison, S.C. Structure of an IκBα/NF-ΚB Complex. Cell 1998, 95, 749–758. [Google Scholar] [CrossRef] [PubMed]
- Hong-Geller, E.; Chaudhary, A.; Lauer, S. Targeting Toll-Like Receptor Signaling Pathways for Design of Novel Immune Therapeutics. Curr. Drug Discov. Technol. 2008, 5, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Rathee, P.; Chaudhary, H.; Rathee, S.; Rathee, D.; Kumar, V.; Kohli, K. Mechanism of Action of Flavonoids as Anti-Inflammatory Agents: A Review. Inflamm. Allergy Drug Targets Former. Curr. Drug Targets Inflamm. Allergy 2009, 8, 229–235. [Google Scholar] [CrossRef]
- Kamboh, A.A.; Khan, M.A.; Kaka, U.; Awad, E.A.; Memon, A.M.; Saeed, M.; Korejo, N.A.; Bakhetgul, M.; Kumar, C. Effect of Dietary Supplementation of Phytochemicals on Immunity and Haematology of Growing Broiler Chickens. Ital. J. Anim. Sci. 2018, 17, 1038–1043. [Google Scholar] [CrossRef]
- Aziz, N.; Kim, M.-Y.; Cho, J.Y. Anti-Inflammatory Effects of Luteolin: A Review of In Vitro, In Vivo, and In Silico Studies. J. Ethnopharmacol. 2018, 225, 342–358. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Lazaro, M. Distribution and Biological Activities of the Flavonoid Luteolin. Mini Rev. Med. Chem. 2009, 9, 31–59. [Google Scholar] [CrossRef]
- Mackei, M.; Molnár, A.; Nagy, S.; Pál, L.; Kővágó, C.; Gálfi, P.; Dublecz, K.; Husvéth, F.; Neogrády, Z.; Mátis, G. Effects of Acute Heat Stress on a Newly Established Chicken Hepatocyte—Nonparenchymal Cell Co-Culture Model. Animals 2020, 10, 409. [Google Scholar] [CrossRef]
- Robinson, T.P.; Bu, D.P.; Carrique-Mas, J.; Fèvre, E.M.; Gilbert, M.; Grace, D.; Hay, S.I.; Jiwakanon, J.; Kakkar, M.; Kariuki, S.; et al. Antibiotic Resistance Is the Quintessential One Health Issue. Trans. R. Soc. Trop. Med. Hyg. 2016, 110, 377–380. [Google Scholar] [CrossRef]
- Manyi-Loh, C.; Mamphweli, S.; Meyer, E.; Okoh, A. Antibiotic Use in Agriculture and Its Consequential Resistance in Environmental Sources: Potential Public Health Implications. Molecules 2018, 23, 795. [Google Scholar] [CrossRef]
- Fair, R.J.; Tor, Y. Antibiotics and Bacterial Resistance in the 21st Century. Perspect. Med. Chem. 2014, 6, PMC.S14459. [Google Scholar] [CrossRef]
- Aljumaah, M.R.; Suliman, G.M.; Abdullatif, A.A.; Abudabos, A.M. Effects of Phytobiotic Feed Additives on Growth Traits, Blood Biochemistry, and Meat Characteristics of Broiler Chickens Exposed to Salmonella Typhimurium. Poult. Sci. 2020, 99, 5744–5751. [Google Scholar] [CrossRef]
- Buchanan, N.P.; Hott, J.M.; Cutlip, S.E.; Rack, A.L.; Asamer, A.; Moritz, J.S. The Effects of a Natural Antibiotic Alternative and a Natural Growth Promoter Feed Additive on Broiler Performance and Carcass Quality. J. Appl. Poult. Res. 2008, 17, 202–210. [Google Scholar] [CrossRef]
- Demir, E.; Sarica, Ş.; Özcan, M.A.; Suiçmez, M. The Use of Natural Feed Additives as Alternatives for an Antibiotic Growth Promoter in Broiler Diets. Br. Poult. Sci. 2003, 44, 44–45. [Google Scholar] [CrossRef]
- Alagawany, M.; Elnesr, S.S.; Farag, M.R.; Abd El-Hack, M.E.; Barkat, R.A.; Gabr, A.A.; Foda, M.A.; Noreldin, A.E.; Khafaga, A.F.; El-Sabrout, K.; et al. Potential Role of Important Nutraceuticals in Poultry Performance and Health—A Comprehensive Review. Res. Vet. Sci. 2021, 137, 9–29. [Google Scholar] [CrossRef] [PubMed]
- Tráj, P.; Herrmann, E.M.; Sebők, C.; Vörösházi, J.; Mackei, M.; Gálfi, P.; Kemény, Á.; Neogrády, Z.; Mátis, G. Protective Effects of Chicoric Acid on Polyinosinic-Polycytidylic Acid Exposed Chicken Hepatic Cell Culture Mimicking Viral Damage and Inflammation. Vet. Immunol. Immunopathol. 2022, 250, 110427. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.K.; Deb, R.; Gaikwad, S.; Saravanan, R.; Mohan, C.M.; Dey, S. Recombinant Flagellin and Its Cross-Talk with Lipopolysaccharide—Effect on Pooled Chicken Peripheral Blood Mononuclear Cells. Res. Vet. Sci. 2013, 95, 930–935. [Google Scholar] [CrossRef] [PubMed]
- Pan, Z.; Fang, Q.; Geng, S.; Kang, X.; Cong, Q.; Jiao, X. Analysis of Immune-Related Gene Expression in Chicken Peripheral Blood Mononuclear Cells Following Salmonella enterica Serovar Enteritidis Infection In Vitro. Res. Vet. Sci. 2012, 93, 716–720. [Google Scholar] [CrossRef] [PubMed]
- Burdelya, L.G.; Brackett, C.M.; Kojouharov, B.; Gitlin, I.I.; Leonova, K.I.; Gleiberman, A.S.; Aygun-Sunar, S.; Veith, J.; Johnson, C.; Haderski, G.J.; et al. Central Role of Liver in Anticancer and Radioprotective Activities of Toll-like Receptor 5 Agonist. Proc. Natl. Acad. Sci. USA 2013, 110, E1857–E1866. [Google Scholar] [CrossRef] [PubMed]
- Sebők, C.; Tráj, P.; Vörösházi, J.; Mackei, M.; Papp, M.; Gálfi, P.; Neogrády, Z.; Mátis, G. Two Sides to Every Question: Attempts to Activate Chicken Innate Immunity in 2D and 3D Hepatic Cell Cultures. Cells 2021, 10, 1910. [Google Scholar] [CrossRef]
- Liu, M.; Cheng, C.; Li, X.; Zhou, S.; Hua, J.; Huang, J.; Li, Y.; Yang, K.; Zhang, P.; Zhang, Y.; et al. Luteolin Alleviates Ochratoxin A Induced Oxidative Stress by Regulating Nrf2 and HIF-1α Pathways in NRK-52E Rat Kidney Cells. Food Chem. Toxicol. 2020, 141, 111436. [Google Scholar] [CrossRef]
- Wang, X.; Wang, L.; Dong, R.; Huang, K.; Wang, C.; Gu, J.; Luo, H.; Liu, K.; Wu, J.; Sun, H.; et al. Luteolin Ameliorates LPS-Induced Acute Liver Injury by Inhibiting TXNIP-NLRP3 Inflammasome in Mice. Phytomedicine 2021, 87, 153586. [Google Scholar] [CrossRef]
- Zou, Y.; Luo, X.; Feng, Y.; Fang, S.; Tian, J.; Yu, B.; Li, J. Luteolin Prevents THP-1 Macrophage Pyroptosis by Suppressing ROS Production via Nrf2 Activation. Chem. Biol. Interact. 2021, 345, 109573. [Google Scholar] [CrossRef]
- Sung, J.; Lee, J. Anti-Inflammatory Activity of Butein and Luteolin Through Suppression of NFκB Activation and Induction of Heme Oxygenase-1. J. Med. Food 2015, 18, 557–564. [Google Scholar] [CrossRef] [PubMed]
- Göbel, T.W.; Schneider, K.; Schaerer, B.; Mejri, I.; Puehler, F.; Weigend, S.; Staeheli, P.; Kaspers, B. IL-18 Stimulates the Proliferation and IFN-γ Release of CD4+ T Cells in the Chicken: Conservation of a Th1-Like System in a Nonmammalian Species1. J. Immunol. 2003, 171, 1809–1815. [Google Scholar] [CrossRef] [PubMed]
- Dai, M.; Xu, C.; Chen, W.; Liao, M. Progress on Chicken T Cell Immunity to Viruses. Cell. Mol. Life Sci. 2019, 76, 2779–2788. [Google Scholar] [CrossRef]
- Cardenas-Garcia, S.; Dunwoody, R.P.; Marcano, V.; Diel, D.G.; Williams, R.J.; Gogal, R.M., Jr.; Brown, C.C.; Miller, P.J.; Afonso, C.L. Effects of Chicken Interferon Gamma on Newcastle Disease Virus Vaccine Immunogenicity. PLoS ONE 2016, 11, e0159153. [Google Scholar] [CrossRef] [PubMed]
- Ingram, J.P.; Brodsky, I.E.; Balachandran, S. Interferon–γ in Salmonella Pathogenesis: New Tricks for an Old Dog. Cytokine 2017, 98, 27–32. [Google Scholar] [CrossRef]
- Stoycheva, M.; Murdjeva, M. Serum Levels of Interferon-γ, Interleukin-12, Tumour Necrosis Factor-α, and Interleukin-10, and Bacterial Clearance in Patients with Gastroenteric Salmonella Infection. Scand. J. Infect. Dis. 2005, 37, 11–14. [Google Scholar] [CrossRef]
- Pie, S.; Matsiota-Bernard, P.; Truffa-Bachi, P.; Nauciel, C. Gamma Interferon and Interleukin-10 Gene Expression in Innately Susceptible and Resistant Mice during the Early Phase of Salmonella Typhimurium Infection. Infect. Immun. 1996, 64, 849–854. [Google Scholar] [CrossRef]
- Beal, R.K.; Powers, C.; Wigley, P.; Barrow, P.A.; Kaiser, P.; Smith, A.L. A Strong Antigen-Specific T-Cell Response Is Associated with Age and Genetically Dependent Resistance to Avian Enteric Salmonellosis. Infect. Immun. 2005, 73, 7509–7516. [Google Scholar] [CrossRef]
- Hoerr, F.J. Clinical Aspects of Immunosuppression in Poultry. Avian Dis. 2010, 54, 2–15. [Google Scholar] [CrossRef]
- Lee, A.J.; Ashkar, A.A. The Dual Nature of Type I and Type II Interferons. Front. Immunol. 2018, 9, 2061. [Google Scholar] [CrossRef] [PubMed]
- Garrido, D.; Alber, A.; Kut, E.; Chanteloup, N.K.; Lion, A.; Trotereau, A.; Dupont, J.; Tedin, K.; Kaspers, B.; Vervelde, L.; et al. The Role of Type I Interferons (IFNs) in the Regulation of Chicken Macrophage Inflammatory Response to Bacterial Challenge. Dev. Comp. Immunol. 2018, 86, 156–170. [Google Scholar] [CrossRef] [PubMed]
- Poh, T.Y.; Pease, J.; Young, J.R.; Bumstead, N.; Kaiser, P. Re-evaluation of chicken CXCR1 determines the true gene structure: CXCLi1 (K60) and CXCLi2 (CAF/interleukin-8) are ligands for this receptor. J. Biol. Chem. 2008, 283, 16408–16415. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-A.; Kim, D.-K.; Kang, O.-H.; Choi, Y.-A.; Park, H.-J.; Choi, S.-C.; Kim, T.-H.; Yun, K.-J.; Nah, Y.-H.; Lee, Y.-M. Inhibitory Effect of Luteolin on TNF-α-Induced IL-8 Production in Human Colon Epithelial Cells. Int. Immunopharmacol. 2005, 5, 209–217. [Google Scholar] [CrossRef]
- Wang, S.; Cao, M.; Xu, S.; Shi, J.; Mao, X.; Yao, X.; Liu, C. Luteolin Alters Macrophage Polarization to Inhibit Inflammation. Inflammation 2020, 43, 95–108. [Google Scholar] [CrossRef]
- Li, L.; Luo, W.; Qian, Y.; Zhu, W.; Qian, J.; Li, J.; Jin, Y.; Xu, X.; Liang, G. Luteolin Protects against Diabetic Cardiomyopathy by Inhibiting NF-ΚB-Mediated Inflammation and Activating the Nrf2-Mediated Antioxidant Responses. Phytomed. Int. J. Phytother. Phytopharm. 2019, 59, 152774. [Google Scholar] [CrossRef]
- Keestra, A.M.; de Zoete, M.R.; Bouwman, L.I.; Vaezirad, M.M.; van Putten, J.P.M. Unique Features of Chicken Toll-like Receptors. Dev. Comp. Immunol. 2013, 41, 316–323. [Google Scholar] [CrossRef]
- Ulfig, A.; Leichert, L.I. The Effects of Neutrophil-Generated Hypochlorous Acid and Other Hypohalous Acids on Host and Pathogens. Cell. Mol. Life Sci. 2021, 78, 385–414. [Google Scholar] [CrossRef]
- Thomas, D.C. The Phagocyte Respiratory Burst: Historical Perspectives and Recent Advances. Immunol. Lett. 2017, 192, 88–96. [Google Scholar] [CrossRef]
- Awadin, W.F.; Eladl, A.H.; El-Shafei, R.A.; El-Adl, M.A.; Aziza, A.E.; Ali, H.S.; Saif, M.A. Effect of Omega-3 Rich Diet on the Response of Japanese Quails (Coturnix coturnix japonica) Infected with Newcastle Disease Virus or Avian Influenza Virus H9N2. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2020, 228, 108668. [Google Scholar] [CrossRef]
- Harmon, B. Avian Heterophils in Inflammation and Disease Resistance. Poult. Sci. 1998, 77, 972–977. [Google Scholar] [CrossRef] [PubMed]
- Surai, P.F.; Kochish, I.I.; Fisinin, V.I.; Kidd, M.T. Antioxidant Defence Systems and Oxidative Stress in Poultry Biology: An Update. Antioxidants 2019, 8, 235. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q. Role of Nrf2 in Oxidative Stress and Toxicity. Annu. Rev. Pharmacol. Toxicol. 2013, 53, 401–426. [Google Scholar] [CrossRef]
- Cai, Q.; Rahn, R.O.; Zhang, R. Dietary Flavonoids, Quercetin, Luteolin and Genistein, Reduce Oxidative DNA Damage and Lipid Peroxidation and Quench Free Radicals. Cancer Lett. 1997, 119, 99–107. [Google Scholar] [CrossRef]
- Kovács, D.; Karancsi, Z.; Farkas, O.; Jerzsele, Á. Antioxidant Activity of Flavonoids in LPS-Treated IPEC-J2 Porcine Intestinal Epithelial Cells and Their Antibacterial Effect against Bacteria of Swine Origin. Antioxidants 2020, 9, 1267. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tráj, P.; Sebők, C.; Mackei, M.; Kemény, Á.; Farkas, O.; Kákonyi, Á.; Kovács, L.; Neogrády, Z.; Jerzsele, Á.; Mátis, G. Luteolin: A Phytochemical to Mitigate S. Typhimurium Flagellin-Induced Inflammation in a Chicken In Vitro Hepatic Model. Animals 2023, 13, 1410. https://doi.org/10.3390/ani13081410
Tráj P, Sebők C, Mackei M, Kemény Á, Farkas O, Kákonyi Á, Kovács L, Neogrády Z, Jerzsele Á, Mátis G. Luteolin: A Phytochemical to Mitigate S. Typhimurium Flagellin-Induced Inflammation in a Chicken In Vitro Hepatic Model. Animals. 2023; 13(8):1410. https://doi.org/10.3390/ani13081410
Chicago/Turabian StyleTráj, Patrik, Csilla Sebők, Máté Mackei, Ágnes Kemény, Orsolya Farkas, Ákos Kákonyi, László Kovács, Zsuzsanna Neogrády, Ákos Jerzsele, and Gábor Mátis. 2023. "Luteolin: A Phytochemical to Mitigate S. Typhimurium Flagellin-Induced Inflammation in a Chicken In Vitro Hepatic Model" Animals 13, no. 8: 1410. https://doi.org/10.3390/ani13081410