
Effects of Panax notoginseng Water Extract on Immune Responses and Digestive Enzymes in White Shrimp Litopenaeus vannamei


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Litopenaeus vannamei
2.2. Preparation of PNWE
2.3. Experimental Design
2.4. Measurement of Immune Parameters
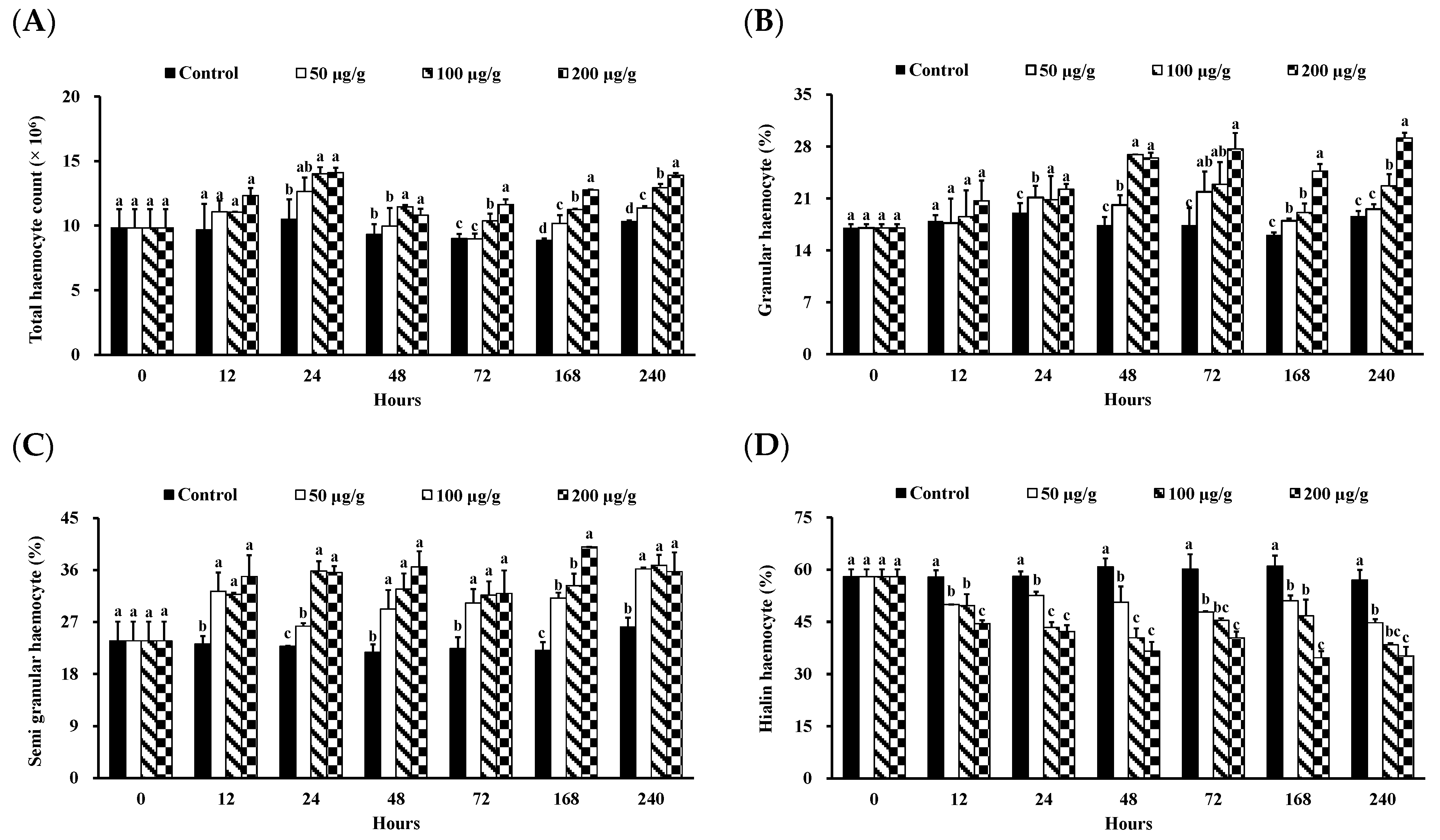
2.4.1. Measurement of THC
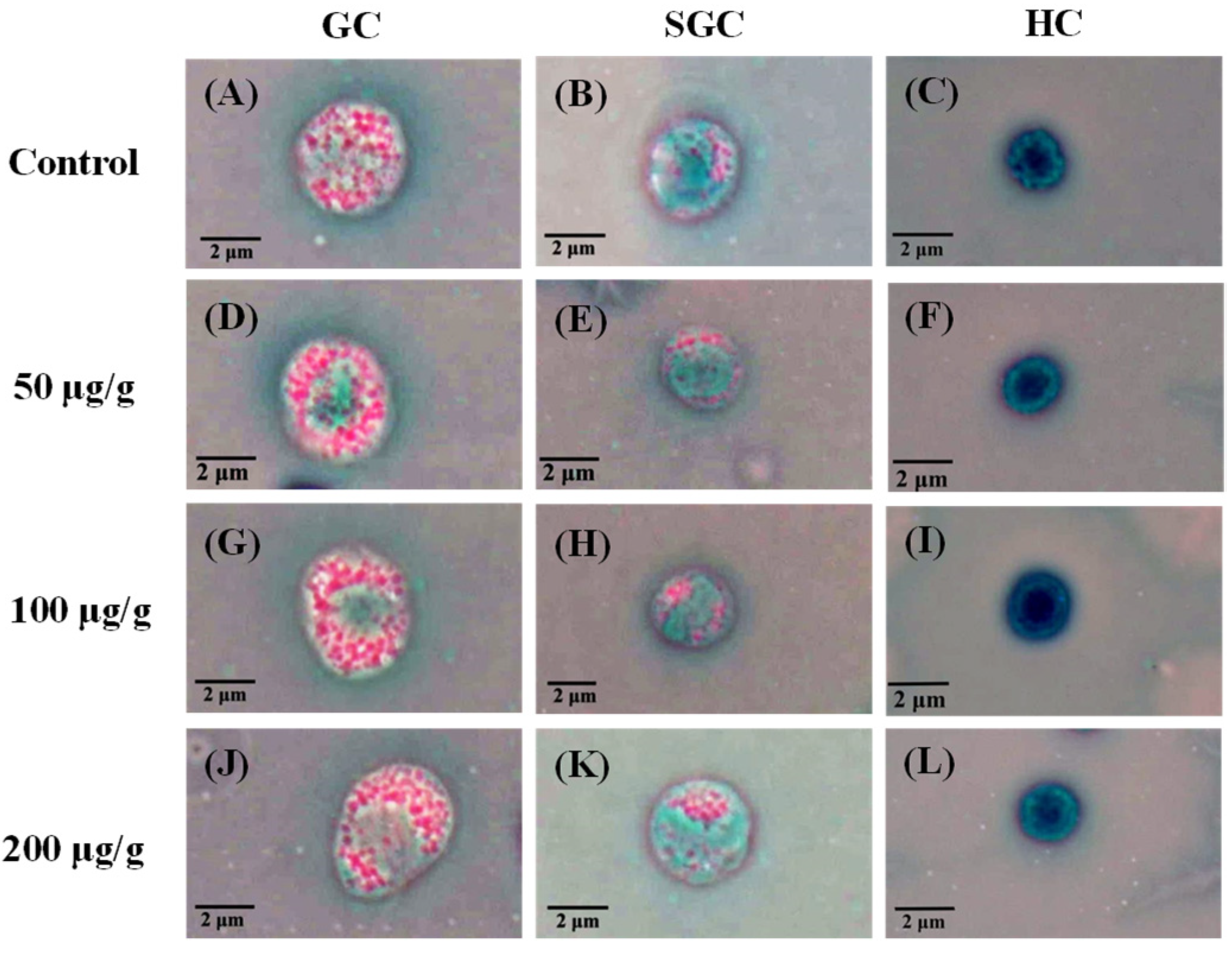
2.4.2. Measurement of DHC
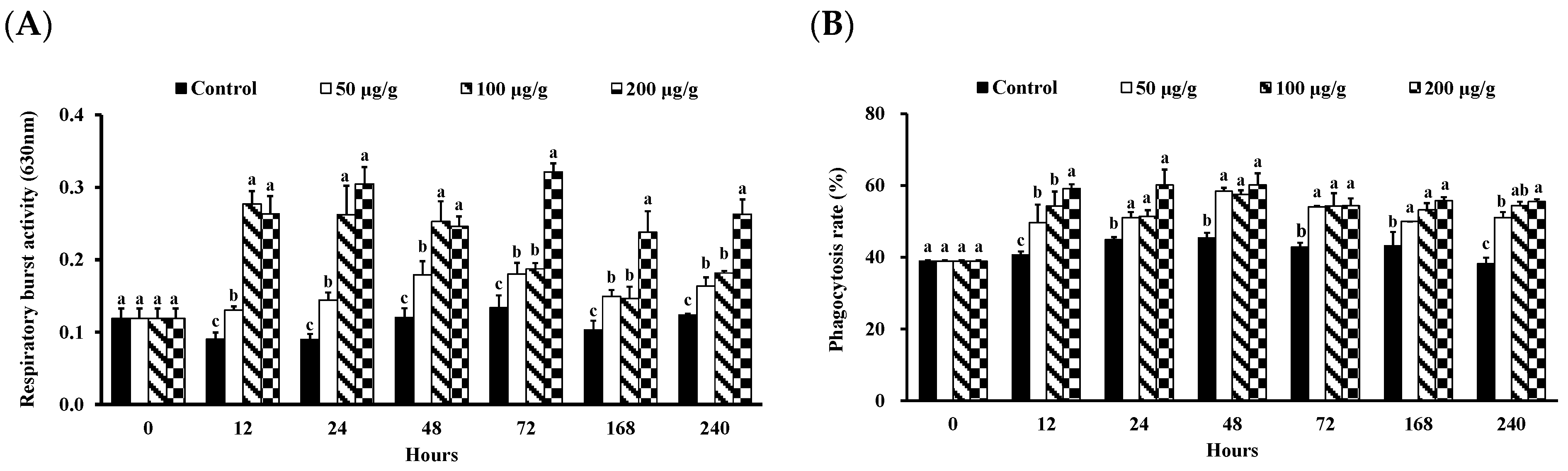
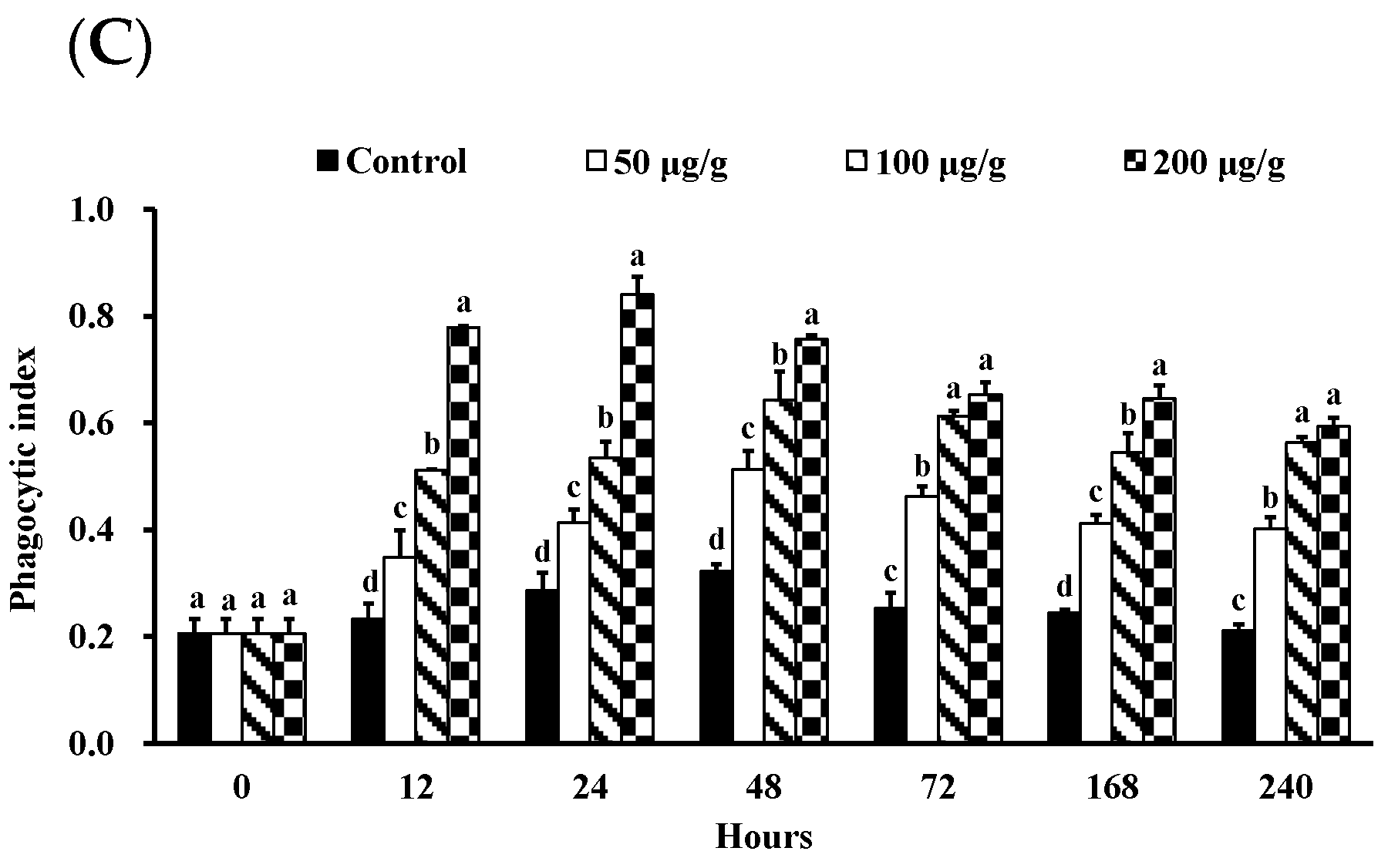
2.4.3. Measurement of Phagocytic Activity
2.4.4. Measurement of RB Activity
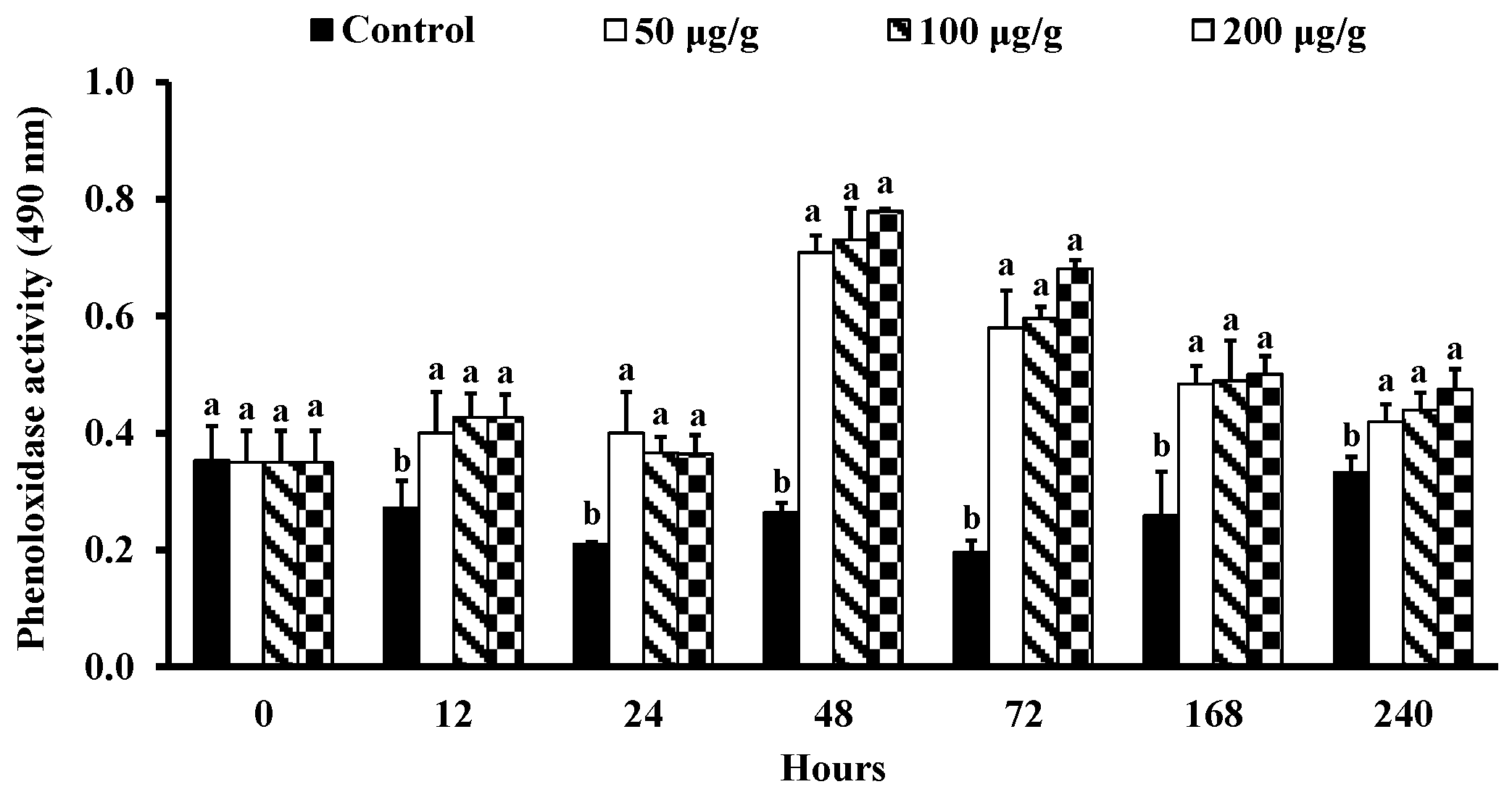
2.4.5. Measurement of PO Activity
2.5. Analysis of Digestive Enzymes
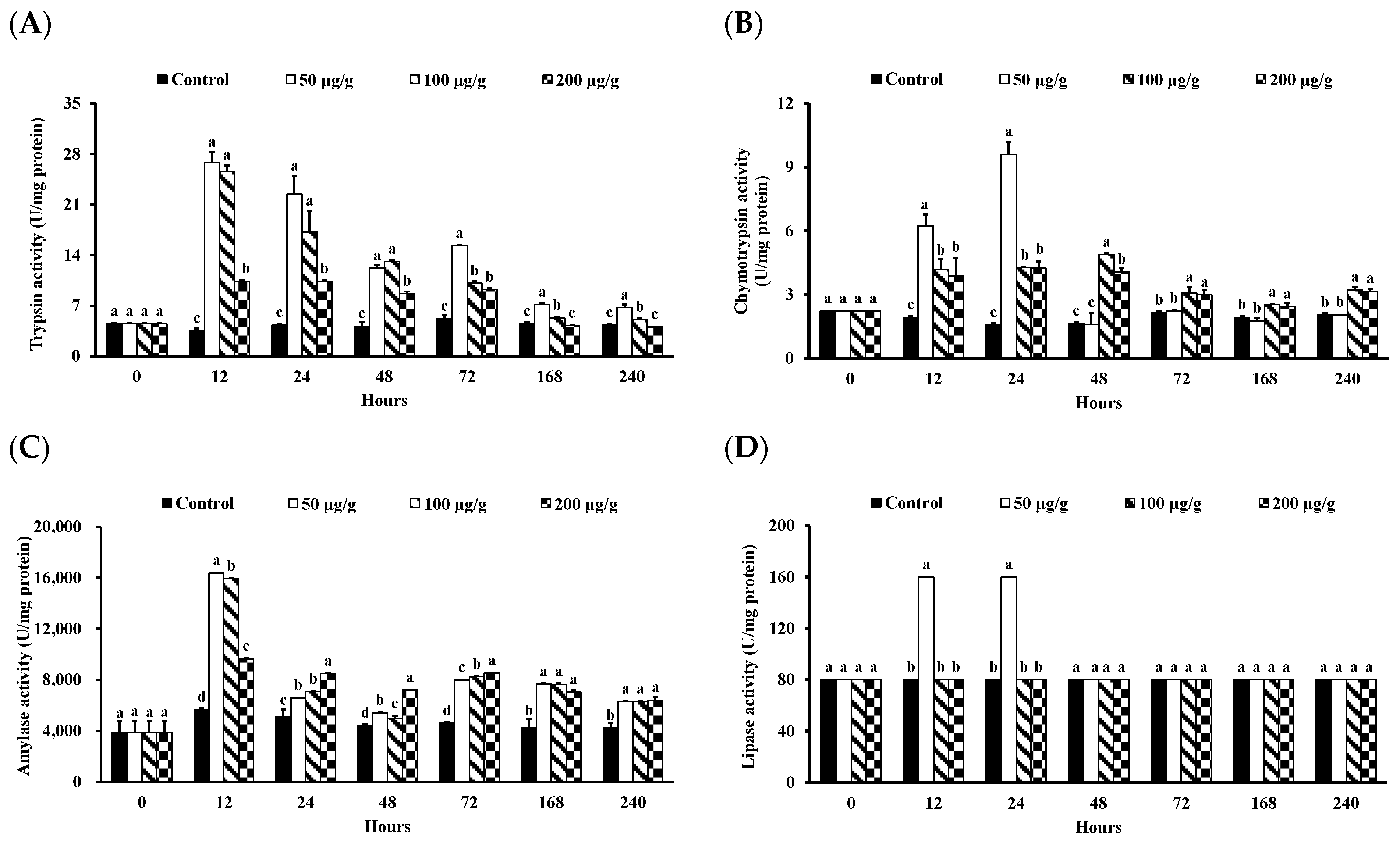
2.5.1. Measurement of Trypsin Activity
2.5.2. Measurement of Chymotrypsin Activity
2.5.3. Measurement of Amylase Activity
2.5.4. Measurement of Lipase Activity
2.6. Analysis of Vibrio Number
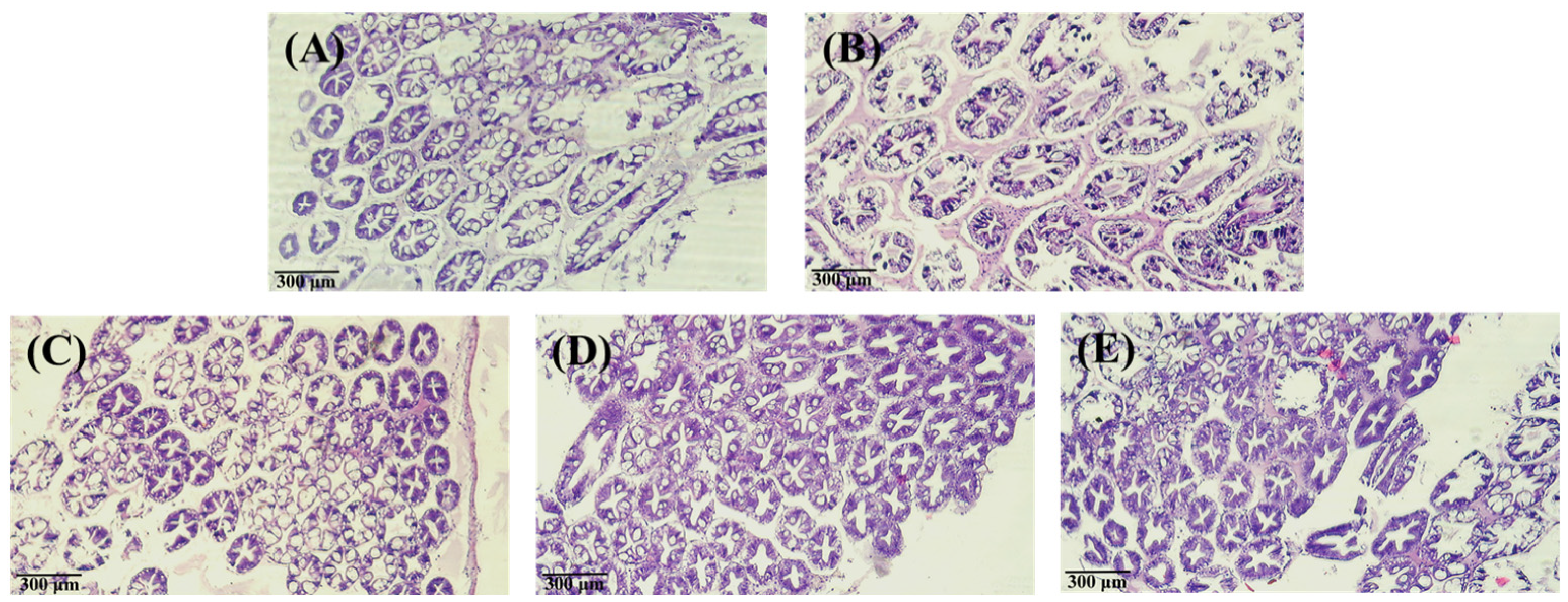
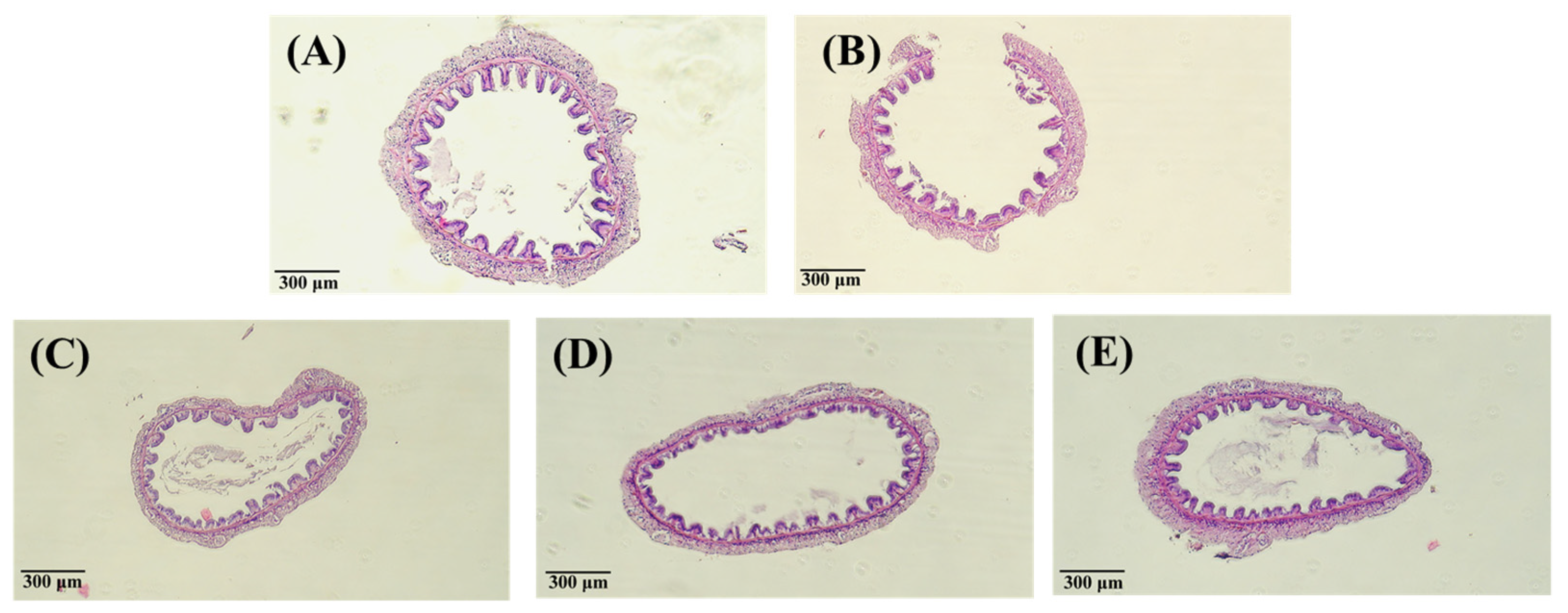
2.7. Analysis of Tissue Staining with Hematoxylin-Eosin
2.8. Statistical Analysis
3. Results
3.1. Effect of the PNWE on the Immune Response of L. vannamei
3.2. Effect of PNWE on the Digestive Enzyme Functions of L. vannamei
3.3. Effect of PNWE on the Variation in Vibrio Number and Histologic Patterns of L. vannamei
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Xia, Y.T.; Cheng, E.H.C.; Zheng, B.Z.Y.; Wu, Q.Y.; Dong, T.T.X.; Duan, R.; Qin, Q.W.; Wang, W.X.; Tsim, K.W.K. Feeding containing the aerial part of Scutellaria baicalensis promotes the growth and nutritive value of rabbit fish Siganus fuscescens. Food Sci. Nutr. 2021, 9, 487–4838. [Google Scholar] [CrossRef]
- Van Doan, H.; Hoseinifar, S.H.; Chitmanat, C.; Jaturasitha, S.; Paolucci, M.; Ashouri, G.; Dawood, M.A.O.; Esteban, M.A. The effects of Thai ginseng, Boesenbergia rotunda powder on mucosal and serum immunity, disease resistance, and growth performance of Nile tilapia (Oreochromis niloticus) fingerlings. Aquaculture 2019, 513, 734388. [Google Scholar] [CrossRef]
- Yin, X.; Zhuang, X.; Liao, M.; Cui, Q.; Yan, C.; Huang, J.; Jiang, Z.; Huang, L.; Luo, Y.; Wang, W. Andrographis paniculata improves growth and non-specific immunity of shrimp Litopenaeus vannamei, and protects it from Vibrio alginolyticus by reducing oxidative stress and apoptosis. Dev. Comp. Immunol. 2023, 139, 104542. [Google Scholar] [CrossRef]
- Ghosh, A.K.; Panda, S.K.; Luyten, W. Anti-vibrio and immune-enhancing activity of medicinal plants in shrimp: A comprehensive review. Fish Shellfish Immunol. 2021, 117, 192–210. [Google Scholar] [CrossRef]
- Xu, C.; Wang, W.; Wang, B.; Zhang, T.; Cui, X.; Pu, Y.; Li, N. Analytical methods and biological activities of Panax notoginseng saponins: Recent trends. J. Ethnopharmacol. 2019, 236, 443–465. [Google Scholar] [CrossRef]
- Zhang, Y.; Bian, Y.; Cui, Q.; Yuan, C.; Lin, Y.; Li, J.; Meng, P. Effect of dietary complex Chinese herbal medicine on growth performance, digestive enzyme activities in tissues and expression of genes involved in the digestive enzymes and antioxidant enzymes and bacterial challenge in Litopenaeus vannamei. Aquac. Res. 2021, 52, 6741–6750. [Google Scholar] [CrossRef]
- Yang, X.; Xiong, X.; Wang, H.; Wang, J. Protective effects of Panax notoginseng saponins on cardiovascular diseases: A comprehensive overview of experimental studies. Evid.-Based Complement. Altern. Med. 2014, 2014, 204840. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Han, Z.; Li, G.; Zhang, S.; Luo, Y. Therapeutic potential and cellular mechanisms of Panax notoginseng on prevention of aging and cell senescence-associated diseases. Aging Dis. 2017, 8, 721–739. [Google Scholar] [CrossRef] [Green Version]
- Sun, Z.; Tan, X.; Ye, H.; Zou, C.; Ye, C.; Wang, A. Effects of dietary Panax notoginseng extract on growth performance, fish composition, immune responses, intestinal histology and immune related genes expression of hybrid grouper (Epinephelus lanceolatus ♂ × Epinephelus fuscoguttatus ♀) fed high lipid diets. Fish Shellfish Immunol. 2018, 73, 234–244. [Google Scholar] [CrossRef]
- Johansson, M.W.; Keyser, P.; Sritunyalucksana, K.; Söderhäll, K. Crustacean haemocytes and haematopoiesis. Aquaculture 2000, 191, 45–52. [Google Scholar] [CrossRef]
- Johansson, M.W.; Söderhäll, K. Isolation and purification of a cell adhesion factor crayfish blood cells. J. Cell Biol. 1988, 106, 1795–1803. [Google Scholar] [CrossRef] [Green Version]
- Aspán, A.; Söderhäll, K. Purification prophenoloxidase from crayfish blood cells and its activation by an endogenous serine proteinase. Insect Biochem. 1991, 21, 363–373. [Google Scholar] [CrossRef]
- Moncada, S.; Palmer, R.M.J.; Higgs, E.A. Nitric oxide: Physiology, pathophysiology, and pharmacology. Pharmacol. Rev. 1991, 43, 109–142. [Google Scholar]
- Dong, J.; Zhao, Y.Y.; Yu, Y.H.; Sun, N.; Li, Y.D.; Wei, H.; Yang, Z.Q.; Li, X.D.; Li, L. Effect of stocking density on growth performance, digestive enzyme activities, and nonspecific immune parameters of Palaemonetes sinensis. Fish Shellfish Immunol. 2018, 73, 37–41. [Google Scholar] [CrossRef]
- Javahery, S.; Noori, A.; Hoseinifar, S.H. Growth performance, immune response, and digestive enzyme activity in Pacific white shrimp, Penaeus vannamei Boone, 1931, fed dietary microbial lysozyme. Fish Shellfish Immunol. 2019, 92, 528–535. [Google Scholar] [CrossRef]
- Alhoshy, M.; Shehata, A.I.; Habib, Y.J.; Abdel-Latif, H.M.R.; Wang, Y.; Zhang, Z. Nutrigenomics in crustaceans: Current status and future prospects. Shellfish Immunol. 2022, 129, 1–12. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations (FAO). The State of World Fisheries and Aquaculture; FAO: Rome, Italy, 2022. [Google Scholar]
- Kumar, S.; Verma, A.K.; Singh, S.P.; Awasthi, A. Immunostimulants for shrimp aquaculture: Paving pathway towards shrimp sustainability. Environ. Prog. Sustain. Energy 2022, 13, 25325–25343. [Google Scholar] [CrossRef]
- Yeh, S.T.; Liu, C.H.; Chen, J.C. Effect of copper sulfate on the immune response and susceptibility to Vibrio alginolyticus in the white shrimp Litopenaeus vannamei. Fish Shellfish Immunol. 2004, 17, 437–446. [Google Scholar] [CrossRef]
- Maftuch; Prasetio, E.; Sudianto, A.; Rozik, M.; Nurdiyani, R.; Sanusi, E.; Nursyam, H.; Fariedah, F.; Marsoedi; Murachman. Improvement of innate immune responses and defense activity in tiger shrimp (Penaeus monodon Fab.) by intramuscular administration of the outer membrane protein Vibrio alginolyticus. SpringerPlus 2013, 2, 2–8. [Google Scholar] [CrossRef] [Green Version]
- Jian, J.; Wu, Z. Influences of traditional Chinese medicine on non-specific immunity of Jian Carp (Cyprinus carpio var. Jian). Fish Shellfish Immunol. 2004, 16, 185–191. [Google Scholar] [CrossRef]
- Trenzado, C.E.; Hidalgo, F.; Villanueva, D.; Furné, M.; Díaz Casado, M.E.; Merino, R.; Sanz, A. Study of the enzymatic digestive profile in three species of Mediterranean sea urchins. Aquaculture 2012, 344–349, 174–180. [Google Scholar] [CrossRef]
- Holmblad, T.; Söderhäll, K. Cell adhension molecules and antioxidative enzymes in a crustacean, possible role in immunity. Aquaculture 1999, 172, 111–123. [Google Scholar] [CrossRef]
- Rodríguez, J.; Le Moullac, G. State of the art of immunological tools and health control of penaeid shrimp. Aquaculture 2000, 191, 109–119. [Google Scholar] [CrossRef]
- Pu, M.; Wu, S. The growth performance, body composition and nonspecific immunity of white shrimps (Litopenaeus vannamei) affected by dietary Astragalus membranaceus polysaccharide. J. Biol. Macromol. 2022, 209, 162–165. [Google Scholar] [CrossRef]
- Munaeni, W.; Yuhana, M.; Setiawati, M.; Wahyudi, A.T. Effect in white shrimp Litopenaeus vannamei of Eleutherine bulbosa (Mill.) Urb. Powder on immune genes expression and resistance against Vibrio parahaemolyticus infection. Fish Shellfish Immunol. 2020, 102, 218–227. [Google Scholar] [CrossRef]
- Abidin, Z.; Huang, H.T.; Hu, Y.F.; Chang, J.J.; Huang, C.Y.; Wu, Y.S.; Nan, F.H. Effect of dietary supplementation with Moringa oleifera leaf extract and Lactobacillus acidophilus on growth performance, intestinal microbiota, immune response, and disease resistance in whiteleg shrimp (Penaeus vannamei). Fish Shellfish Immunol. 2022, 127, 876–890. [Google Scholar] [CrossRef]
- Huang, H.T.; Lee, P.T.; Liao, Z.H.; Chen, H.Y.; Nan, F.H. Effects of Phyllanthus amarus extract on nonspecific immune responses, growth, and resistance to Vibrio alginolyticus in white shrimp Litopenaeus vannamei. Fish Shellfish Immunol. 2020, 107, 1–8. [Google Scholar] [CrossRef]
- Rahardjo, S.; Vauza, M.A.T.; Rukmono, D.; Wiradana, P.A. Supplementation of hairy eggplant (Solanum ferox) and bitter ginger (Zingiber zerumbet) extracts as phytobiotic agents on whiteleg shrimp (Litopenaeus vannamei). J. Adv. Vet. Anim. Res. 2022, 9, 78. [Google Scholar] [CrossRef]
- Pan, S.; Jiang, L.; Wu, S. Stimulating effects of polysaccharide from Angelica sinensis on the nonspecific immunity of white shrimps (Litopenaeus vannamei). Fish Shellfish Immunol. 2018, 74, 170–174. [Google Scholar] [CrossRef]
- Wu, Y.S.; Lee, M.C.; Huang, C.T.; Kung, T.C.; Huang, C.Y.; Nan, F.H. Effects of traditional medical herbs “minor bupleurum decoction” on the non-specific immune responses of white shrimp (Litopenaeus vannamei). Fish Shellfish Immunol. 2017, 64, 218–225. [Google Scholar] [CrossRef]
- Li, H.; Xu, C.; Zhou, L.; Dong, Y.; Su, Y.; Wang, X.; Qin, J.G.; Chen, L.; Li, E. Beneficial effects of dietary β-glucan on growth and health status of Pacific white shrimp Litopenaeus vannamei at low salinity. Fish Shellfish Immunol. 2019, 91, 315–324. [Google Scholar] [CrossRef]
- Abdel-Tawwab, M.; Selema, T.A.A.; Khalil, R.H.; El-Sabbagh, N.; Eldessouki, E.A.; Fawzy, R.M.; Abd El-Naby, A.S. The growth performance, antioxidant and immune responses, and disease resistance of Litopenaeus vannamei fed on diets supplemented with Indian ginseng (Withania somnifera). Fish Shellfish Immunol. 2022, 128, 19–27. [Google Scholar] [CrossRef]
- Chen, K.; Sagada, G.; Xu, B.; Liu, Y.; Zheng, L.; Tegomo, A.F.; Yang, Y.; Sun, Y.; Wang, J.; Longshaw, M.; et al. Evaluation of methanotroph (Methylococcus capsulatus, Bath) bacteria meal as an alternative protein source for growth performance, digestive enzymes, and health status of Pacific white shrimp (Litopenaeus vannamei). Aquac. Int. 2022, 30, 1693–1710. [Google Scholar] [CrossRef]
- Gong, H.; Qin, Z.; Chen, Z.; Li, J.; Chang, Z.; Li, J.; Chen, P. Effects of dietary tannic acid on growth, digestion, immunity and resistance to ammonia stress, and intestinal microbial community in Pacific white shrimp (Litopenaeus vannamei). Fishes 2022, 7, 327. [Google Scholar] [CrossRef]
- Xu, A.; Shang-Guan, J.; Li, Z.; Gao, Z.; Huang, Y.C.; Chen, Q. Effects of dietary Chinese herbal medicines mixture on feeding attraction activity, growth performance, nonspecific immunity and digestive enzyme activity of Japanese seabass (Lateolabrax japonicus). Aquac. Res. 2020, 17, 100304. [Google Scholar] [CrossRef]
- Wangkahart, E.; Wachiraamonloed, S.; Lee, P.T.; Subramani, P.A.; Qi, Z.; Wang, B. Impacts of Aegle marmelos fruit extract as a medicinal herb on growth performance, antioxidant and immune responses, digestive enzymes, and disease resistance against Streptococcus agalactiae in Nile tilapia (Oreochromis niloticus). Fish Shellfish Immunol. 2022, 120, 402–410. [Google Scholar] [CrossRef]
- Hong, S.J.; Wan, J.B.; Zhang, Y.; Hu, G.; Lin, H.C.; Seto, S.W.; Kwan, Y.W.; Lin, Z.X.; Wang, Y.T.; Lee, S.M.Y. Angiogenic effect of saponin extract from Panax notoginseng on HUVECs in vitro and zebrafish in vivo. Phytother. Res. 2009, 23, 677–686. [Google Scholar] [CrossRef]
- Yang, B.R.; Hong, S.J.; Lee, S.M.Y.; Cong, W.H.; Wan, J.B.; Zhang, Z.R.; Zhang, Q.W.; Zhang, Y.; Wang, Y.T.; Lin, Z.X. Pro-angiogenic activity of notoginsenoside R1 in human umbilical vein endothelial cells in vitro and in a chemical-induced blood vessel loss model of zebrafish in vivo. Chin. J. Integr. Med. 2016, 22, 420–429. [Google Scholar] [CrossRef]
- Fan, Y.; Wang, X.; Wang, Y.; Liu, H.; Yu, X.; Li, L.; Ye, H.; Wang, S.; Gai, C.; Xu, L.; et al. Potential effects of dietary probiotics with Chinese herb polysaccharides on the growth performance, immunity, disease resistance, and intestinal microbiota of rainbow trout (Oncorhynchus mykiss). J. World Aquac. Soc. 2021, 52, 1194–1208. [Google Scholar] [CrossRef]
- Ping, T.N.; Hsieh, S.L.; Wang, J.J.; Chen, J.B.; Wu, C.C. Panax notoginseng suppresses bone morphogenetic protein-2 expression in EA. hy926 endothelial cells by inhibiting the noncanonical NF-κB and Wnt/β-Catenin signaling pathways. Plants 2022, 11, 3265. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group (µg (g shrimp)−1) | Vibrio Numbers (×107) after Time Elapsed | ||||||
|---|---|---|---|---|---|---|---|
| 0 | 12 | 24 | 48 | 72 | 168 | 240 | |
| 0 | 12.23 ± 2.56 a | 10.14 ± 0.43 a | 11.26 ± 0.16 a | 10.13 ± 0.36 a | 10.48 ± 5.59 a | 10.25 ± 3.25 b | 10.16 ± 0.21 a |
| 50 | 12.23 ± 2.56 a | 8.30 ± 1.38 a | 7.03 ± 0.96 a | 7.68 ± 1.27 ab | 3.66 ± 1.17 b | 2.64 ± 1.15 b | 4.73 ± 1.68 b |
| 100 | 12.23 ± 2.56 a | 8.07 ± 0.30 a | 6.78 ± 1.52 a | 4.17 ± 0.29 c | 3.69 ± 0.36 b | 1.70 ± 0.06 b | 2.92 ± 0.30 b |
| 200 | 12.23 ± 2.56 a | 10.55 ± 3.97 a | 8.05 ± 2.57 a | 6.56 ± 1.51 bc | 2.27 ± 0.74 b | 1.20 ± 0.10 b | 2.56 ± 0.56 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.-T.; Kuo, C.-L.; Wu, C.-C.; Liu, C.-H.; Hsieh, S.-L. Effects of Panax notoginseng Water Extract on Immune Responses and Digestive Enzymes in White Shrimp Litopenaeus vannamei. Animals 2023, 13, 1131. https://doi.org/10.3390/ani13071131
Chen Y-T, Kuo C-L, Wu C-C, Liu C-H, Hsieh S-L. Effects of Panax notoginseng Water Extract on Immune Responses and Digestive Enzymes in White Shrimp Litopenaeus vannamei. Animals. 2023; 13(7):1131. https://doi.org/10.3390/ani13071131
Chicago/Turabian StyleChen, Ya-Ting, Chia-Ling Kuo, Chih-Chung Wu, Chun-Hung Liu, and Shu-Ling Hsieh. 2023. "Effects of Panax notoginseng Water Extract on Immune Responses and Digestive Enzymes in White Shrimp Litopenaeus vannamei" Animals 13, no. 7: 1131. https://doi.org/10.3390/ani13071131