
Transcriptomics and Proteomics of Haemonchus contortus in Response to Ivermectin Treatment
 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Approval
2.2. Sample Collection
2.3. Ivermectin Treatment
2.4. Transcriptomics
2.4.1. RNA Extraction, Sequencing, and Identification of Differentially Expressed Transcripts (DETs)
2.4.2. Sequence Filtering, Functional Annotation, and Analysis of Differentially Expressed Genes
2.5. Quantitative Proteomics (iTRAQ)
2.5.1. Protein Extraction, Quantization, and SDS-PAGE Electrophoresis
2.5.2. Proteolysis and ITRAQ Labeling
2.5.3. Reversed-Phase Liquid Chromatography (RPLC)
2.5.4. LC-MS/MS Analysis and Data Processing
2.6. Validation of RNA-Seq Results Using Quantitative Real-Time PCR (q-PCR)
3. Results
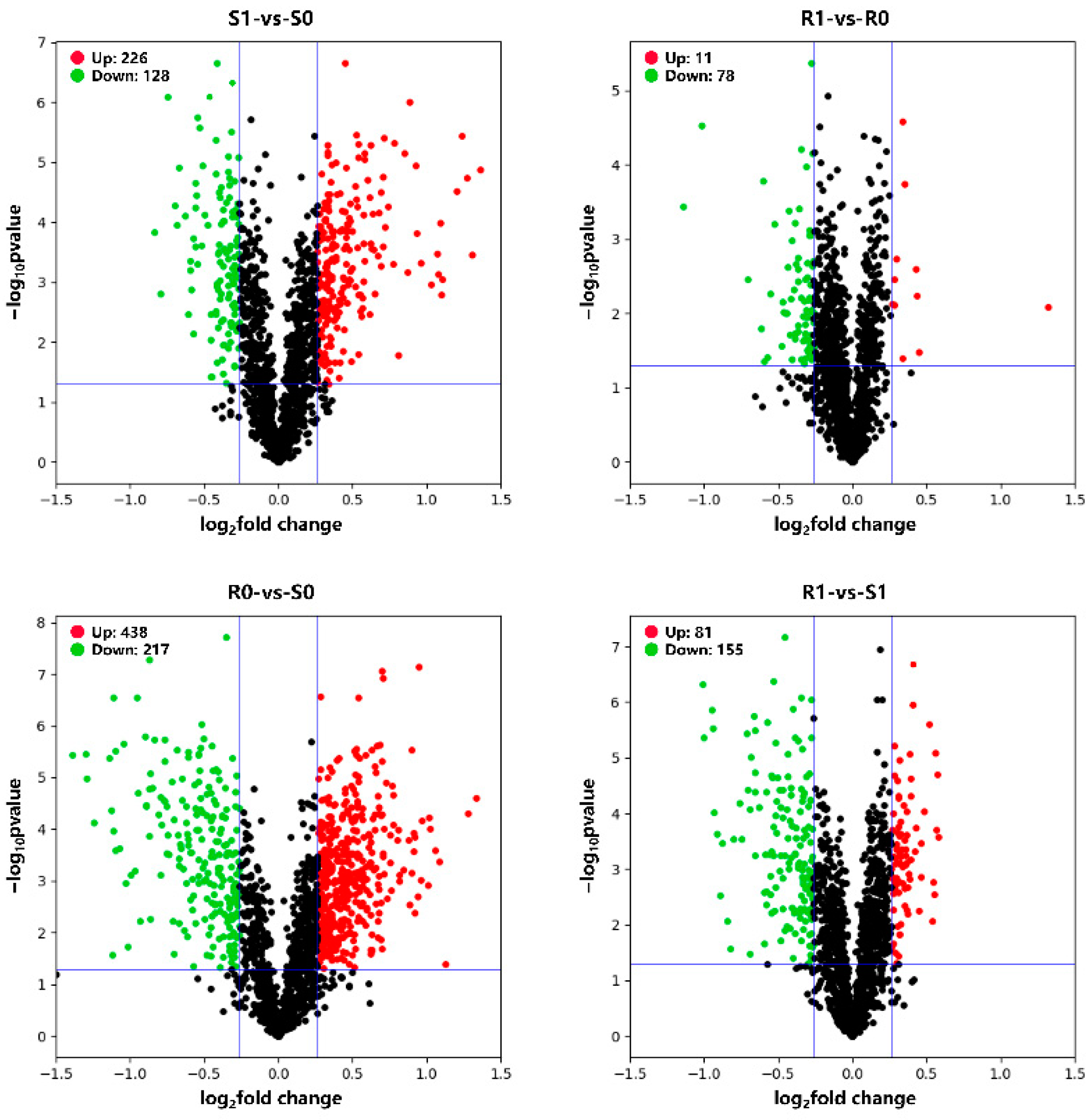
3.1. Differentially Expressed Genes (DEGs) and Differentially Expressed Proteins (DEPs)
3.2. Integrated Analysis of Transcriptome and Proteome
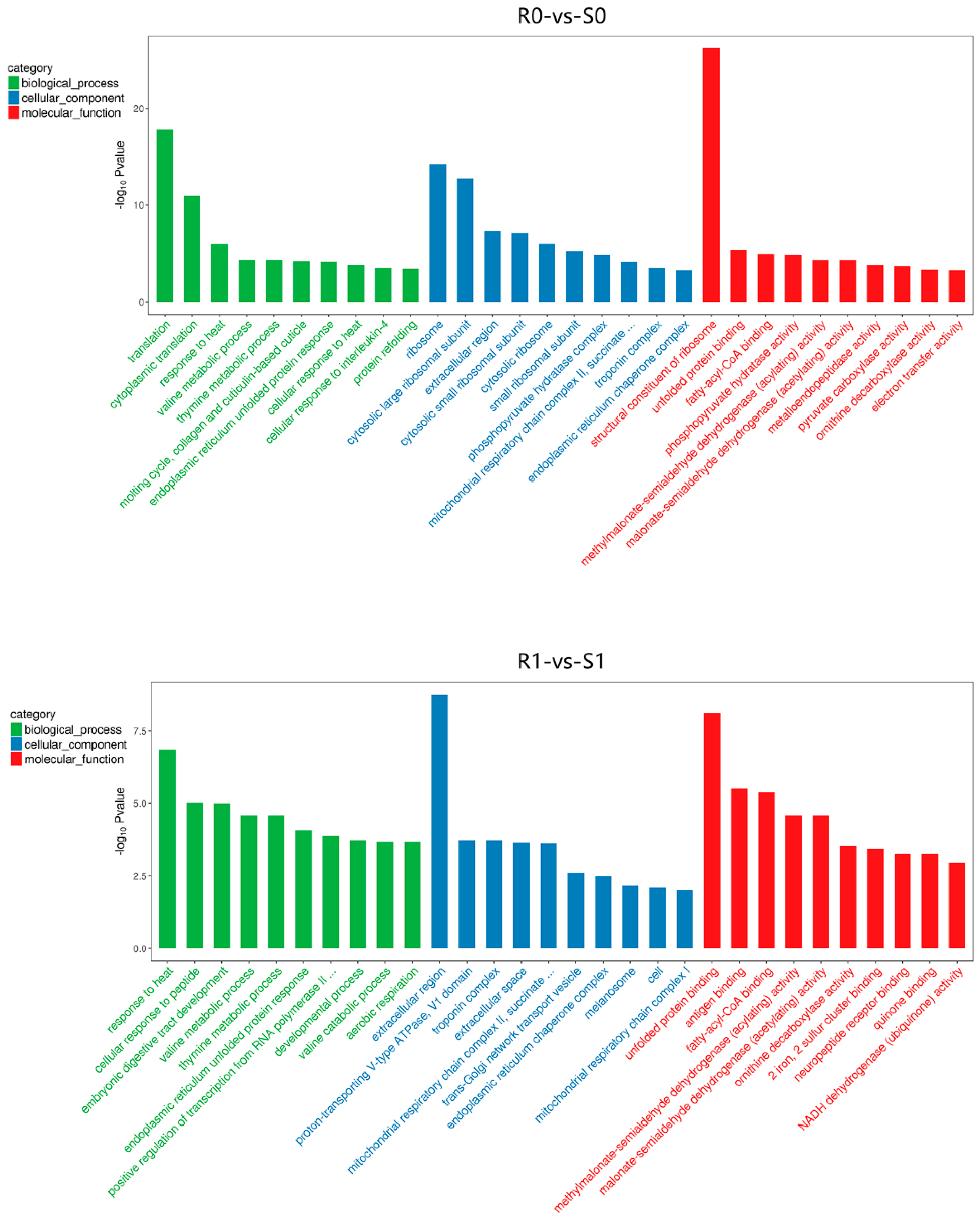
3.3. Gene Ontology (GO) Analysis of DEGs and DEPs
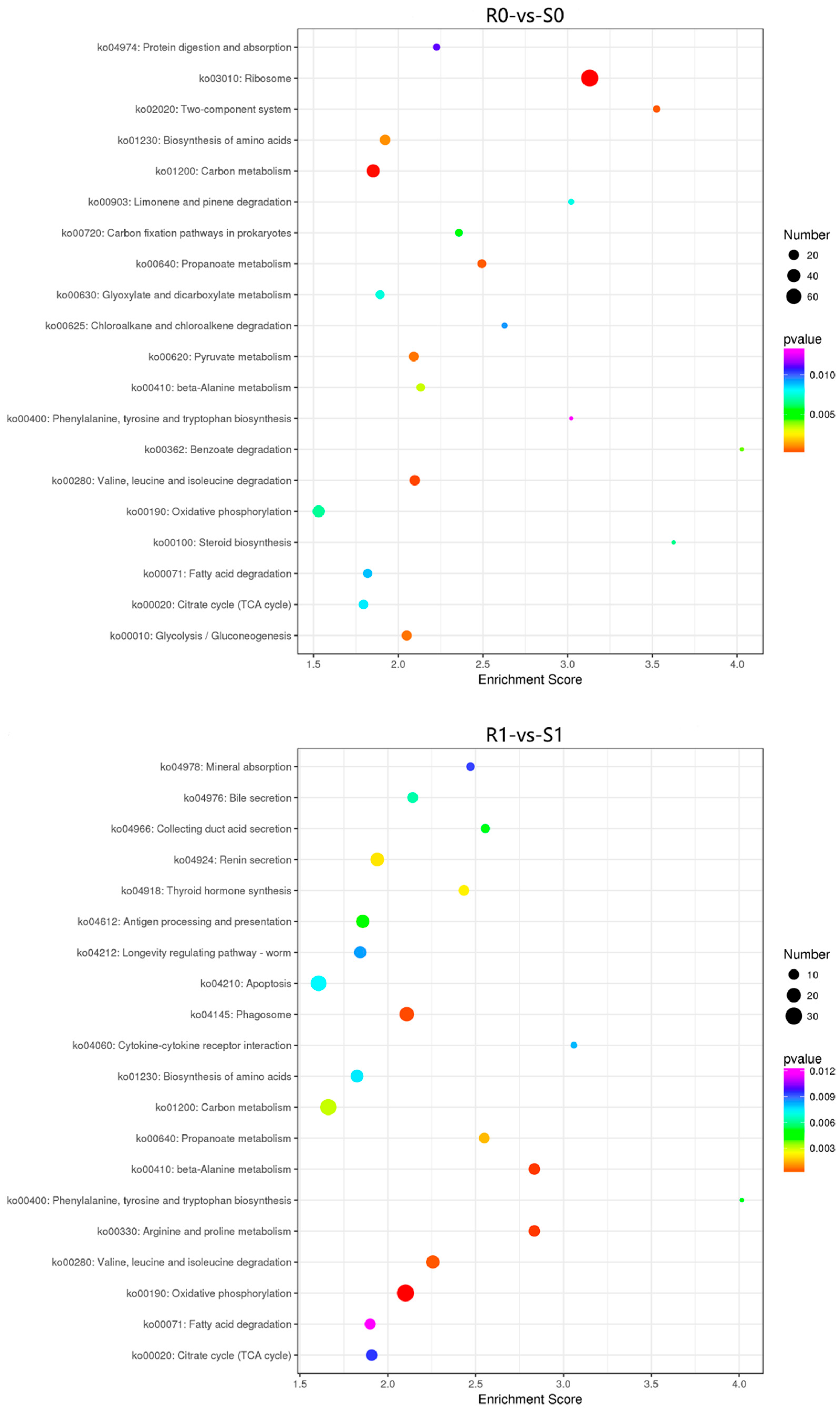
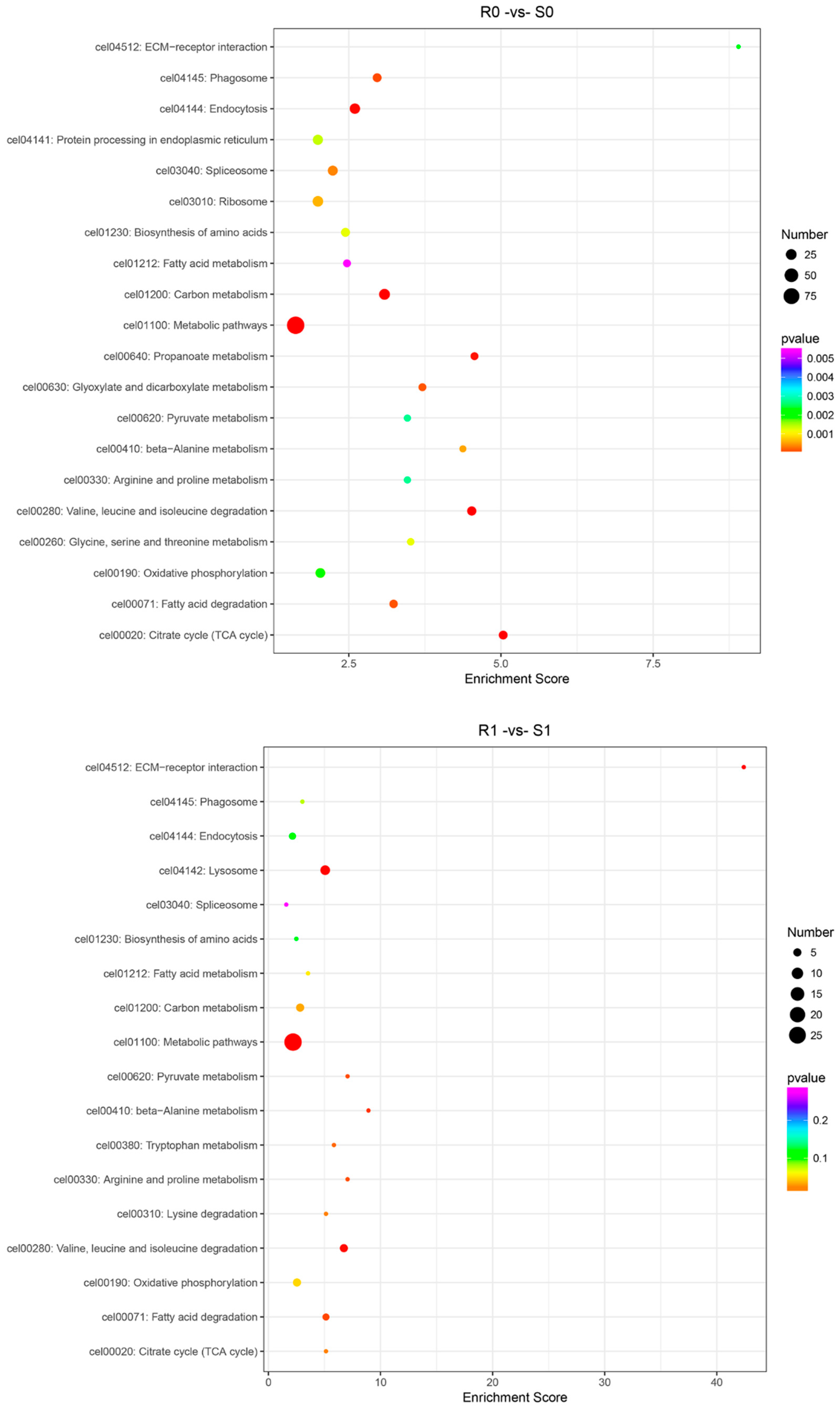
3.4. KEGG Pathway Analysis of the DEGs and DEPs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, C.; Li, F.; Zhang, Z.; Yang, X.; Ahmad, A.A.; Li, X.; Du, A.; Hu, M. Recent research progress in China on Haemonchus contortus. Front. Microbiol. 2017, 8, 1509. [Google Scholar] [CrossRef] [Green Version]
- James, C.E.; Davey, M.W. Increased expression of ABC transport proteins is associated with ivermectin resistance in the model nematode Caenorhabditis elegans. Int. J. Parasitol. 2009, 39, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Perry, B.D.; Randolph, T.F.; MyDermott, J.J.; Sones, K.R.; Thornton, P.K. Investing in Animal Health Research to Alleviate Poverty; International Livestock Research Institute (ILRI): Nairobi, Kenya, 2002. [Google Scholar]
- Peter, J.W.; Chandrawathani, P. Haemonchus contortus: Parasite problem No. 1 from tropics—Polar Circle. Problems and prospects for control based on epidemiology. Trop. Biomed. 2005, 22, 131–137. [Google Scholar] [PubMed]
- Luo, X.; Shi, X.; Yuan, C.; Ai, M.; Ge, C.; Hu, M.; Feng, X.; Yang, X. Genome-wide SNP analysis using 2b-RAD sequencing identifies the candidate genes putatively associated with resistance to ivermectin in Haemonchus contortus. Parasites Vectors 2017, 10, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartley, D.J.; McAllister, H.; Bartley, Y.; Dupuy, J.; Menez, C.; Alvinerie, M.; Jackson, F.; Lespine, A. P-glycoprotein interfering agents potentiate ivermectin susceptibility in ivermectin sensitive and resistant isolates of Teladorsagia circumcincta and Haemonchus contortus. Parasitology 2009, 136, 1081–1088. [Google Scholar] [CrossRef] [PubMed]
- Õmura, S.; Crump, A. The life and times of ivermectin—A success story. Nat. Rev. Microbiol. 2004, 2, 984–989. [Google Scholar] [CrossRef]
- Geary, T.G. Ivermectin 20 years on: Maturation of a wonder drug. Trends. Parasitol. 2005, 21, 530–532. [Google Scholar] [CrossRef]
- Kaplan, R.M. Drug resistance in nematodes of veterinary importance: A status report. Trends. Parasitol. 2004, 20, 477–481. [Google Scholar] [CrossRef]
- Kaplan, R.M.; Vidyashankar, A.N. An inconvenient truth: Global worming and anthelmintic resistance. Vet. Parasitol. 2012, 186, 70–78. [Google Scholar] [CrossRef]
- Kotze, A.C.; Hunt, P.W.; Skuce, P.; von Samson-Himmelstjerna, G.; Martin, R.J.; Sager, H.; Krücken, J.; Hodgkinson, J.; Lespine, A.; Jex, A.R.; et al. Recent advances in candidate-gene and whole-genome approaches to the discovery of anthelmintic resistance markers and the description of drug/receptor interactions. Int. J. Parasitol. Drugs Drug Resist. 2014, 4, 164–184. [Google Scholar] [CrossRef] [Green Version]
- Gill, J.H. Avermectin/milbemycin resistance in trichostrongyloid nematodes. Int. J. Parasitol. 1998, 28, 863–877. [Google Scholar] [CrossRef] [PubMed]
- Martin, R.J.; Murray, I.; Robertson, A.P.; Bjorn, H.; Sangster, N. Anthelmintics and ion-channels: After a puncture, use a patch. Int. J. Parasitol. 1998, 28, 849–862. [Google Scholar] [CrossRef] [PubMed]
- Blackhall, W.J.; Poulet, J.F.; Prichard, R.K.; Beech, R.N. Haemonchus contortus: Selection at a glutamate-gated chloride channel gene in ivermectin- and moxidectin-selected strains. Exp. Parasitol. 1998, 90, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Blackhall, W.J.; Prichard, R.K.; Beech, R.N. Selection at a c-aminobutyric acid receptor gene in Haemonchus contortus resistant to avermectins/milbemycins. Mol. Biochem. Parasitol. 2003, 131, 137–145. [Google Scholar] [CrossRef]
- Kerboeuf, D.; Guégnard, F. Anthelmintics are substrates and activators of nematode P glycoprotein. Antimicrob. Agents Chemother. 2011, 55, 2224–2232. [Google Scholar] [CrossRef] [Green Version]
- Janssen, I.J.I.; Krücken, J.; Demeler, J.; von Samson-Himmelstjerna, G. Transgenically expressed Parascaris P-glycoprotein-11 can modulate ivermectin susceptibility in Caenorhabditis elegans. Int. J. Parasitol. Drugs Drug Resist. 2015, 5, 44–47. [Google Scholar] [CrossRef] [Green Version]
- Kaschny, M.; Demeler, J.; Janssen, I.J.; Kuzmina, T.A.; Besognet, B.; Kanellos, T.; Kerboeuf, D.; von Samson-Himmelstjerna, G.; Krücken, J. Macrocyclic Lactones Differ in Interaction with Recombinant P-Glycoprotein 9 of the Parasitic Nematode Cylicocylus elongatus and Ketoconazole in a Yeast Growth Assay. PLoS Pathog. 2015, 11, e1004781. [Google Scholar] [CrossRef] [Green Version]
- Ardelli, B.F.; Guerriero, S.B.; Prichard, R.K. Ivermectin imposes selection pressure on P-glycoprotein from Onchocerca volvulus: Linkage disequilibrium and genotype diversity. Parasitology 2006, 132, 375–386. [Google Scholar] [CrossRef] [Green Version]
- Pacheco, P.A.; Louvandini, H.; Giglioti, R.; Wedy, B.C.R.; Ribeiro, J.C.; Verissimo, C.J.; Ferreira, J.F.S.; Amarante, A.F.T.; Katiki, L.M. Phytochemical modulation of P-Glycoprotein and its gene expression in an ivermectin-resistant Haemonchus contortus isolate in vitro. Vet. Parasitol. 2022, 305, 109713. [Google Scholar] [CrossRef]
- Dicker, A.J.; Nisbet, A.J.; Skuce, P.J. Gene expression changes in a P-glycoprotein (Tci-pgp-9) putatively associated with ivermectin resistance in Teladorsagia circumcincta. Int. J. Parasitol. 2011, 41, 935–942. [Google Scholar] [CrossRef]
- Ardelli, B.F.; Prichard, R.K. Inhibition of P-glycoprotein enhances sensitivity of Caenorhabditis elegans to ivermectin. Vet. Parasitol. 2013, 191, 264–275. [Google Scholar] [CrossRef] [PubMed]
- Mate, L.; Ballent, M.; Cantón, C.; Lanusse, C.; Ceballos, L.; Alvarez L, L.I.; Liron, J.P. ABC-transporter gene expression in ivermectin-susceptible and resistant Haemonchus contortus isolates. Vet. Parasitol. 2022, 302, 109647. [Google Scholar] [CrossRef] [PubMed]
- Cwiklinski, K.; Merga, J.Y.; Lake, S.L.; Hartley, C.; Matthews, J.B.; Paterson, S.; Hodgkinson, J.E. Transcriptome analysis of a parasitic clade V nematode: Comparative analysis of potential molecular anthelmintic targets in Cylicostephanus goldi. Int. J. Parasitol. 2013, 43, 917–927. [Google Scholar] [CrossRef] [Green Version]
- Sangster, N.C. Pharmacology of anthelmintic resistance in cyathostomes: Will it occur with the avermectin/milbemycins? Vet. Parasitol. 1999, 85, 189–204. [Google Scholar] [CrossRef]
- Ménez, C.; Alberich, M.; Courtot, E.; Guegnard, F.; Blanchard, A.; Aguilaniu, H.; Lespine, A. The transcription factor NHR-8: A new target to increase ivermectin efficacy in nematodes. PLoS Pathog. 2019, 15, e1007598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yilmaz, E.; Ramünke, S.; Demeler, J.; Krücken, J. Comparison of constitutive and thiabendazole-induced expression of five cytochrome P450 genes in fourth-stage larvae of Haemonchus contortus isolates with different drug susceptibility identifies one gene with high constitutive expression in a multi-resistant isolate. Int. J. Parasitol. Drugs Drug Resist. 2017, 7, 362–369. [Google Scholar]
- Liu, Y.; Luo, X.; Li, J.; Wang, P.; Teng, B.; Wang, R.; Wang, X.; Yang, X. Using feeding and motility patterns for ivermectin resistance detecting in Haemonchus contortus larvae. Exp. Parasitol. 2022, 238, 108230. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Trinity: Reconstructing a full-length transcriptome without a genome from RNA-Seq data. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Kanehisa, M.; Araki, M.; Goto, S.; Hattori, M.; Hirakawa, M.; Itoh, M.; Katayama, T.; Kawashima, S.; Okuda, S.; Tokimatsu, T.; et al. KEGG for linking genomes to life and the environment. Nucleic. Acids. Res. 2008, 36, D480–D484. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; Van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts, A.; Pachter, L. Streaming fragment assignment for real-time analysis of sequencing experiments. Nat. Methods 2013, 10, 71–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anders, S.; Huber, W. Differential Expression of RNA-Seq Data at the Gene Level—The DESeq Package; European Molecular Biology Laboratory: Heidelberg, Germany, 2012. [Google Scholar]
- Smith, P.K.; Krohn, R.I.; Hermanson, G.T.; Mallia, A.K.; Gartner, F.H.; Provenzano, M.; Fujimoto, E.K.; Goek, N.M.; Olson, B.J.; Klenk, D.C. Measurement of protein using bicinchoninic acid. Anal. Biochem. 1985, 150, 76–85. [Google Scholar] [CrossRef]
- Candiano, G.; Bruschi, M.; Musante, L.; Santucci, L.; Ghiggeri, G.M.; Carnemolla, B.; Orecchia, P.; Zardi, L.; Righetti, P.G. Blue silver: A very sensitive colloidal Coomassie G-250 staining for proteome analysis. Electrophoresis 2004, 25, 1327–1333. [Google Scholar] [CrossRef] [PubMed]
- Wiśniewski, J.R.; Zougman, A.; Nagaraj, N.; Mann, M. Universal sample preparation method for proteome analysis. Nat. Methods 2009, 6, 359–362. [Google Scholar] [CrossRef] [PubMed]
- Gilleard, J.S. Understanding anthelmintic resistance: The need for genomics and genetics. Int. J. Parasitol. 2006, 36, 1227–1239. [Google Scholar] [CrossRef]
- Gilleard, J.S. Haemonchus contortus as a paradigm and model to study anthelmintic drug resistance. Parasitology 2013, 140, 1506–1522. [Google Scholar] [CrossRef]
- Kotze, A.C.; Prichard, R.K. Anthelmintic resistance in Haemonchus contortus: History, mechanisms and diagnosis. Adv. Parasitol. 2016, 93, 397–428. [Google Scholar]
- Gasser, R.B.; Schwarz, E.M.; Korhonen, P.K.; Young, N.D. Understanding Haemonchus contortus better through genomics and transcriptomics. Adv. Parasitol. 2016, 93, 519–567. [Google Scholar] [PubMed]
- Chen, Y.; McCarthy, D.; Robinson, M.; Smyth, G.K. edgeR: Differential Expression Analysis of Digital Gene Expression Data User’s Guide. Bioconductor User’s Guide. Available online: http://www.bioconductor.org/packages/release/bioc/vignettes/edgeR/inst/doc/edgeRUsersGuide.pdf (accessed on 17 September 2008).
- Shaheen, S.; Fawaz, F.; Shah, S.; Büsselberg, D. Differential expression and pathway analysis in drug-resistant triple-negative breast cancer cell lines using RNASeq analysis. Int. J. Mol. Sci. 2018, 19, 1810. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X. Screening of Genes Related to Albendazole Resistance of Haemonchus contortus and Study on Its Molecular Regulation. Master’s Thesis, Inner Mongolia Agricultural University, Hohhot, China, 2019. [Google Scholar]
- Qin, H.; Lo, N.W.S.; Loo, J.F.C.; Lin, X.; Yim, A.K.Y.; Tsui, S.K.W.; Lau, T.C.; Ip, M.; Chan, T.F. Comparative transcriptomics of multidrug-resistant Acinetobacter baumannii in response to antibiotic treatments. Sci. Rep. 2018, 8, 3515. [Google Scholar] [CrossRef] [Green Version]
- Nelson, D.R.; Kamataki, T.; Waxman, D.J.; Guengerich, F.P.; Estabrook, R.W.; Feyereise, R.; Gonzalez, F.J.; Coon, M.J.; Gunsalus, I.C.; Gotoh, O.; et al. The P450 superfamily: Update on new sequences, gene mapping, accession numbers, early trivial names of enzymes, and nomenclature. DNA Cell Biol. 1993, 12, 1–51. [Google Scholar] [CrossRef] [PubMed]
- Daborn, P.; Boundy, S.; Yen, J.; Pittendrigh, B. DDT resistance in Drosophila correlates with Cyp6g1 over-expression and confers cross-resistance to the neonicotinoid imidacloprid. Mol. Genet. Genom. 2001, 266, 556–563. [Google Scholar] [CrossRef]
- Schlenke, T.A.; Begun, D.J. Strong selective sweep associated with a transposon insertion in Drosophila simulans. Proc. Natl. Acad. Sci. USA 2004, 101, 1626–1631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amenya, D.A.; Naguran, R.; Lo, T.C.; Ranson, H.; Spillings, B.L.; Wood, O.R.; Brooke, B.D.; Coetzee, M.; Koekemoer, L.L. Over expression of a cytochrome P450 (CYP6P9) in a major African malaria vector, Anopheles funestus, resistant to pyrethroids. Insect Mol. Biol. 2008, 17, 9–25. [Google Scholar] [CrossRef]
- Menzel, R.; Bogaert, T.; Achazi, R. A systematic gene expression screen of Caenorhabditis elegans cytochrome P450 genes reveals CYP35 as strongly xenobiotic inducible. Arch. Biochem. Biophys. 2001, 395, 158–168. [Google Scholar] [CrossRef]
- Menzel, R.; Rodel, M.; Kulas, J.; Steinberg, C.E. CYP35: Xenobiotically induced gene expression in the nematode Caenorhabditis elegans. Arch. Biochem. Biophys. 2005, 438, 93–102. [Google Scholar] [CrossRef] [Green Version]
- Laing, S.T.; Ivens, A.; Laing, R.; Ravikumar, S.; Butler, V.; Woods, D.J.; Gilleard, J.S. Characterization of the xenobiotic response of Caenorhabditis elegans to the anthelmintic drug albendazole and the identification of novel drug glucoside metabolites. Biochem. J. 2010, 432, 505–516. [Google Scholar] [CrossRef] [Green Version]
- Kotze, A.C.; Dobson, R.J.; Chandler, D. Synergism of rotenone by piperonyl butoxide in Haemonchus contortus and Trichostrongylus colubriformis in vitro: Potential for drug-synergism through inhibition of nematode oxidative detoxification pathways. Vet. Parasitol. 2006, 136, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Kawalek, J.C.; Rew, R.S.; Heavner, J. Glutathione-S-transferase, a possible drug-metabolizing enzyme, in Haemonchus contortus: Comparative activity of a cambendazole-resistant and a susceptible strain. Int. J. Parasitol. 1984, 14, 173–175. [Google Scholar] [CrossRef] [PubMed]
- Labade, C.P.; Jadhav, A.R.; Ahire, M.; Zinjarde, S.S.; Tamhane, V.A. Role of induced glutathione-S-transferase from Helicoverpa armigera (Lepidoptera: Noctuidae) HaGST-8 in detoxification of pesticides. Ecotoxicol. Environ. Saf. 2018, 147, 612–621. [Google Scholar] [CrossRef] [PubMed]
- Pugazhendhi, A.; Dhanarani, S.; Shankar, C.; Prakash, P.; Ranganathan, K.; Saratale, R.G.; Thamaraiselvi, K. Electrophoretic pattern of glutathione S-transferase (GST) in antibiotic resistance Gram-positive bacteria from poultry litter. Microb. Pathog. 2017, 110, 285–290. [Google Scholar] [CrossRef]
- Reid, W.R.; Sun, H.; Becnel, J.J.; Clark, A.G.; Scott, J.G. Overexpression of a glutathione S-transferase (Mdgst) and a galactosyltransferase-like gene (Mdgt1) is responsible for imidacloprid resistance in house flies. Pest Manag. Sci. 2019, 75, 37–44. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, K.; Yamada, N. Identification of a diazinon-metabolizing glutathione S-transferase in the silkworm, Bombyx mori. Sci. Rep. 2016, 6, 30073. [Google Scholar] [CrossRef] [Green Version]
- Lumjuan, N.; Rajatileka, S.; Changsom, D.; Wicheer, J.; Leelapat, P.; Prapanthadara, L.; Somboon, P.; Lycett, G.; Ranson, H. The role of the Aedes aegypti Epsilon glutathione transferases in conferring resistance to DDT and pyrethroid insecticides. Insect Biochem. Mol. Biol. 2011, 41, 203–209. [Google Scholar] [CrossRef]
- Wang, Y.; Qiu, L.; Ranson, H.; Lumjuan, N.; Hemingway, J.; Setzer, W.N.; Meehan, E.J.; Chen, L. Structure of an insect epsilon class glutathione S-transferase from the malaria vector Anopheles gambiae provides an explanation for the high DDT-detoxifying activity. J. Struct. Biol. 2008, 164, 228–235. [Google Scholar] [CrossRef]
- Stasiuk, S.J.; MacNevin, G.; Workentine, M.L.; Gray, D.; Redman, E.; Bartley, D.; Morrison, A.; Sharma, N.; Colwell, D.; Ro, D.K.; et al. Similarities and differences in the biotransformation and transcriptomic responses of Caenorhabditis elegans and Haemonchus contortus to five different benzimidazole drugs. Int. J. Parasitol. Drugs Drug Resist. 2019, 11, 13–29. [Google Scholar] [CrossRef]
- Matoušková, P.; Lecová, L.; Laing, R.; Dimunová, D.; Vogel, H.; Stuchlíková, L.R.; Nguyen, L.T.; Kellerová, P.; Vokřál, I.; Lamka, J.; et al. UDP-glycosyltransferase family in Haemonchus contortus: Phylogenetic analysis, constitutive expression, sex-differences and resistance-related differences. Int. J. Parasitol. Drugs Drug Resist. 2018, 8, 420–429. [Google Scholar]












Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Wang, X.; Luo, X.; Wang, R.; Zhai, B.; Wang, P.; Li, J.; Yang, X. Transcriptomics and Proteomics of Haemonchus contortus in Response to Ivermectin Treatment. Animals 2023, 13, 919. https://doi.org/10.3390/ani13050919
Liu Y, Wang X, Luo X, Wang R, Zhai B, Wang P, Li J, Yang X. Transcriptomics and Proteomics of Haemonchus contortus in Response to Ivermectin Treatment. Animals. 2023; 13(5):919. https://doi.org/10.3390/ani13050919
Chicago/Turabian StyleLiu, Yang, Xiaomin Wang, Xiaoping Luo, Rui Wang, Bintao Zhai, Penglong Wang, Junyan Li, and Xiaoye Yang. 2023. "Transcriptomics and Proteomics of Haemonchus contortus in Response to Ivermectin Treatment" Animals 13, no. 5: 919. https://doi.org/10.3390/ani13050919