
Effect of Detergents Based on Sodium Dodecyl Sulfate on Functional Metrics of Frozen–Thawed Collared Peccary (Pecari tajacu) Semen
 , , , ,
, , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Ethics and Husbandry
2.2. Experimental Design
2.3. Semen Collection and Initial Evaluation
2.4. Semen Freezing–Thawing Procedures
2.5. Computer-Aided Semen Analysis
2.6. Sperm Membrane Functionality
2.7. Plasma Membrane Integrity and Mitochondrial Activity
2.8. Sperm Binding Ability Assay
2.9. Statistical Analysis
3. Results
3.1. Fresh Semen Parameters
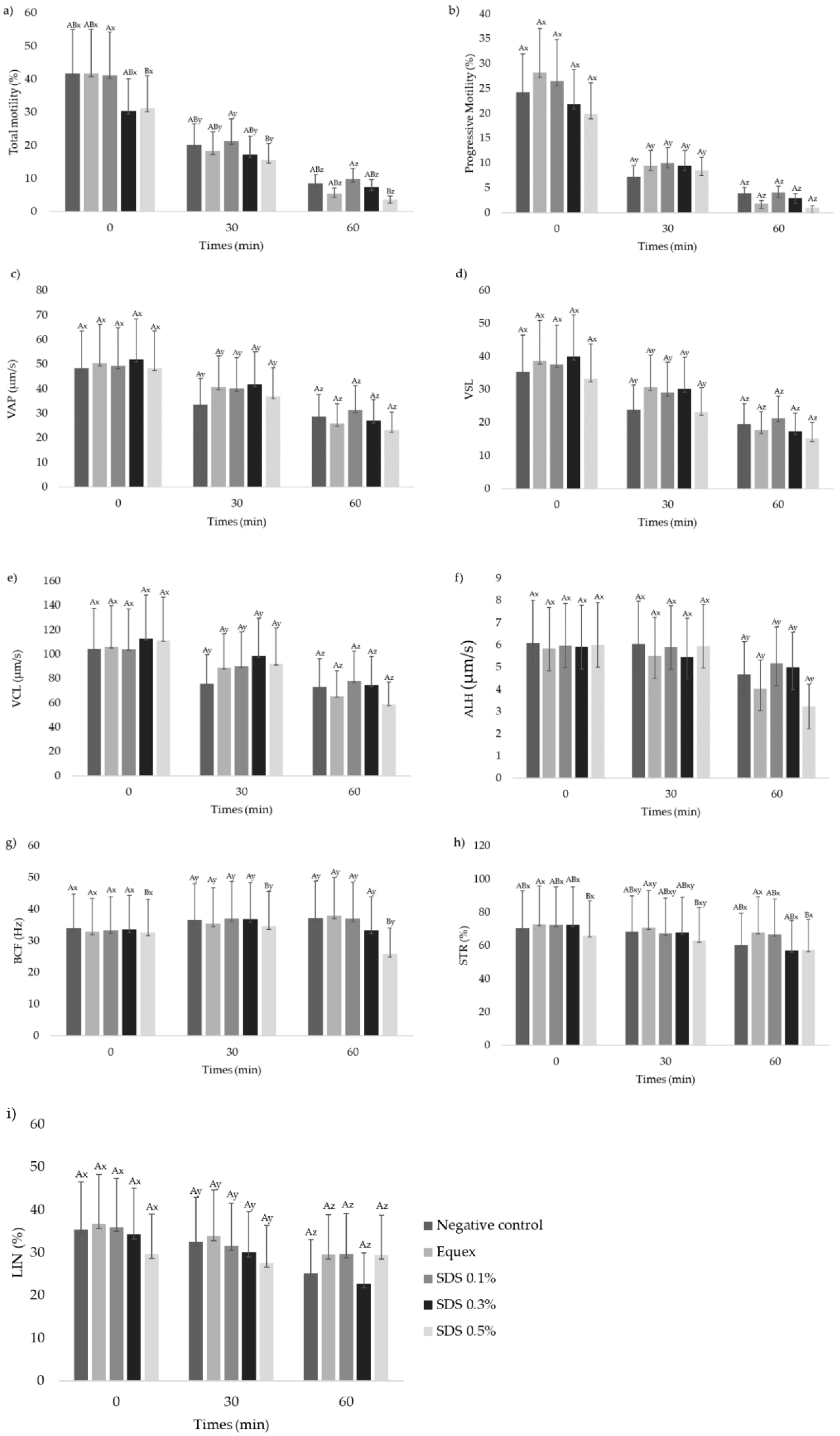
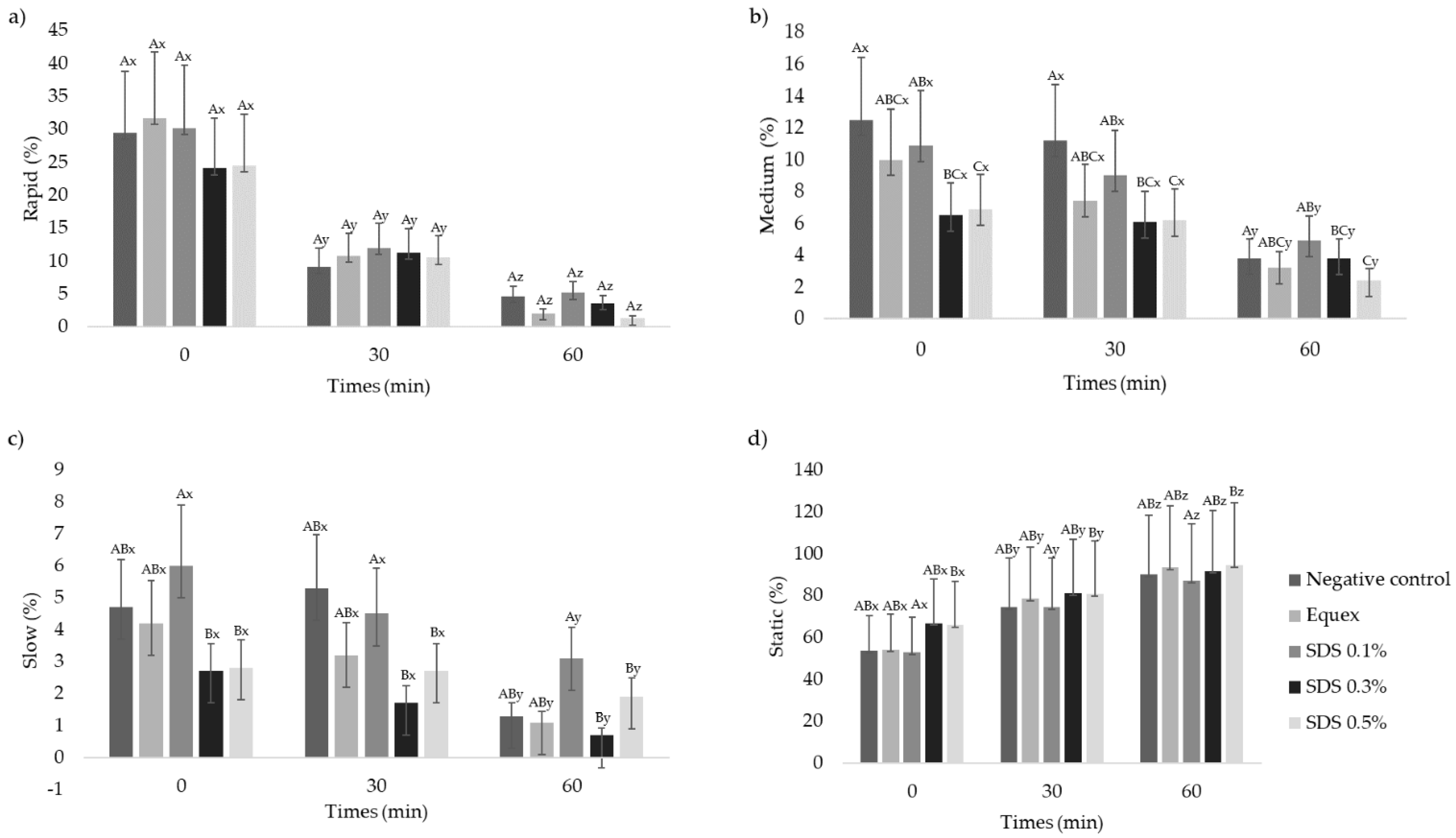
3.2. Sperm Motility Characteristics, Kinetic Parameters and Subpopulations
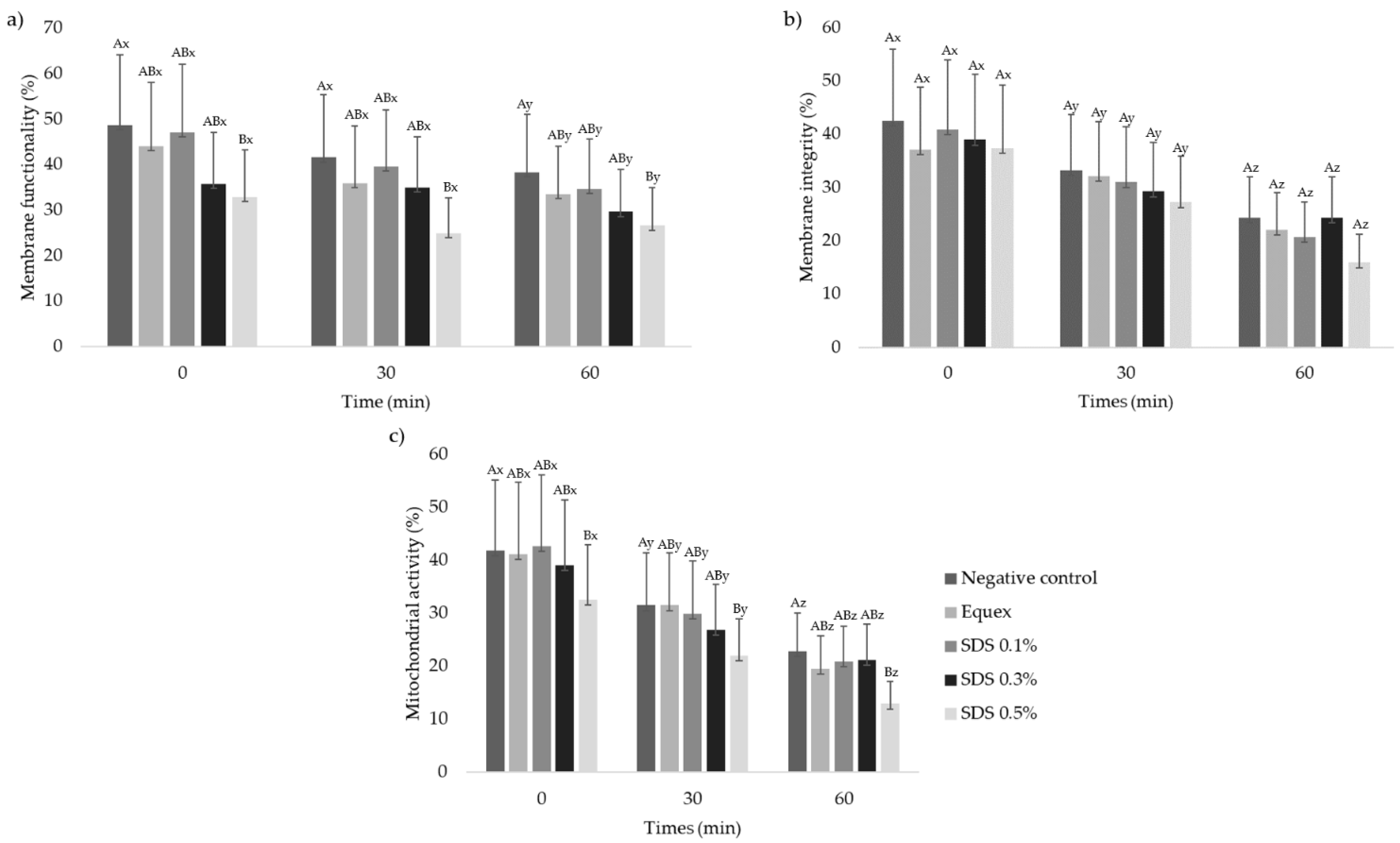
3.3. Sperm Membrane Functionality and Integrity and Mitochondrial Activity
3.4. Sperm-Binding Ability
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Comizzoli, P. Biobanking efforts and new advances in male fertility preservation for rare and endangered species. Asian J. Androl. 2015, 17, 640–645. [Google Scholar] [CrossRef] [PubMed]
- Gongora, J.; Reyna-Hurtado, R.; Beck, H.; Taber, A.; Altrichter, M.; Keuroghlian, A. Pecari tajacu. IUCN Red List. Threat. Species 2011, e.T41777A10562361. [Google Scholar] [CrossRef]
- Castelo, T.S.; Bezerra, F.S.B.; Lima, G.L.; Alves, H.M.; Oliveira, I.R.S.; Santos, E.A.A.; Peixoto, G.C.X.; Silva, A.R. Effect of centrifugation and sugar supplementation on the semen cryopreservation of captive collared peccaries (Tayassu tajacu). Cryobiology 2010, 61, 275–279. [Google Scholar] [CrossRef] [PubMed]
- Alves, H.M.; Oliveira, I.R.S.; Castelo, T.S.; Lima, G.L.; Souza, A.L.P.; Moreira, M.A.P.; Paula, V.V.; Silva, A.R. Comparison of different glycerol and egg yolk concentrations added to tris-based extender for the collared peccaries (Tayassu tajacu) semen freezing. Reprod. Domest. Anim. 2013, 48, 506–511. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.A.; Peixoto, G.C.X.; Lima, G.L.; Bezerra, J.A.B.; Campos, L.B.; Paiva, A.L.C.; Paula, V.V.; Silva, A.R. Cryopreservation of collared peccaries (Tayassu tajacu) semen using a powdered coconut water (ACP-116c) based extender plus various concentrations of egg yolk and glycerol. Theriogenology 2012, 78, 605–611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campos, L.B.; Silva, M.A.; Bezerra, J.A.B.; Castelo, T.S.; Peixoto, G.C.X.; Silva, A.R. Survival of collared peccaries (Pecari tajacu) sperm after freezing-thawing using different extenders. Acta Sci. Vet. 2014, 42, 1–7. [Google Scholar]
- Bezerra, L.G.P.; Souza, A.L.P.; Lago, A.E.A.; Campos, L.B.; Nunes, T.L.; Paula, V.V.; Oliveira, M.F.; Silva, A.R. Addition of Equex STM to extender improves post-thawing longevity of collared peccaries’ sperm. Biopresev. Biobank. 2019, 17, 143–147. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Sharma, D.; Singhal, S.; Kumar, A.; Singh, A.K.; Honparkhe, M. Sodium dodecyl sulphate, N-octyl β-D glu-copyranoside and 4-methoxy phenyl β-D glucopyranoside effect on post-thaw sperm motion and viability traits of Mur-rah buffalo (Bubalus bubalis) bulls. Cryobiology 2022, 107, 1–12. [Google Scholar] [CrossRef]
- Peña, A.I.; Lugilde, L.L.; Barrio, M.; Herradón, P.G.; Quintela, L.A. Effects of Equex from different sources on post-thaw survival, longevity and intracellular Ca2+ concentration of dog spermatozoa. Theriogenology 2003, 59, 1725–1739. [Google Scholar] [CrossRef] [PubMed]
- Zambelli, D.; Iacono, E.; Raccagni, R.; Merlo, B. Quality and fertilizing ability of electroejaculated cat spermatozoa frozen with or without Equex STM Paste. Theriogenology 2010, 73, 886–892. [Google Scholar] [CrossRef]
- Pursel, V.G.; Schulman, L.L.; Johnson, L.A. Effect of Orvus ES Paste on acrosome morphology, motility and fertilizing capacity of frozen-thawed boar sperm. J. Anim. Sci. 1978, 47, 198–202. [Google Scholar] [CrossRef]
- Silva, M.H.M.; Valadão, L.; Silva, F.M. Effect of the tensioactive Agent SDS on extender’s boar semen viability after thawing. Am. J. App. Sci. 2021, 18, 9–14. [Google Scholar] [CrossRef]
- Souza, A.L.P.; Lima, G.L.; Peixoto, C.G.X.; Silva, A.M.; Oliveira, M.F.; Silva, A.R. Evaluation of anesthetic protocol for the collection of semen from captive collared peccaries (Tayassu tajacu) by electroejaculation. Anim. Reprod. Sci. 2009, 116, 370–375. [Google Scholar] [CrossRef]
- Silva, A.M.; Peixoto, G.C.X.; Bezerra, J.A.B.; Castelo, T.S.; Santos, E.A.A.; Silva, A.R. Relationship between Neubauer counting chamber and spectrophotometer used for the sperm concentration determination in collared peccaries (Pecari tajacu). Ciência Rural 2014, 44, 1494–1498. [Google Scholar] [CrossRef] [Green Version]
- Moreira, S.S.J.; Santos, C.S.; Castelo, T.S.; Bezerra, L.G.P.; Praxedes, E.C.G.; Matos, T.M.; Souza-Junior, J.B.F.; Feijó, F.M.C.; Comizzoli, P.; Silva, A.R. Investigating the need for antibiotic supplementation to the extender used for semen cryopreservation in collared peccaries. Front. Vet. Sci. 2022, 9, 954921. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.A.; Peixoto, G.C.; Castelo, T.S.; Lima, G.L.; Silva, A.M.; Oliveira, M.F.; Silva, A.R. Cryopreservation of collared peccary (Pecari tajacu) semen using different freezing curves, straw sizes, and thawing rates. Cryobiology 2013, 67, 50–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Souza, A.L.P.; Lima, G.L.; Peixoto, G.C.X.; Silva, A.M.; Oliveira, M.F.; Silva, A.R. Use of Aloe vera-based extender for chilling and freezing collared peccary (Pecari tajacu) semen. Theriogenology 2016, 85, 1432–1438. [Google Scholar] [CrossRef]
- Santos, E.A.A.; Sousa, P.C.; Peixoto, G.C.X.; Simao, B.R.; Oliveira, M.F.; Silva, A.R. Establishing the hypoosmotic swelling test for sperm analysis in collared peccaries (Pecari tajacu). Arq. Bras. Med. Vet. Zootec. 2013, 65, 1257–1260. [Google Scholar] [CrossRef] [Green Version]
- Sousa, P.C.; Santos, E.A.A.; Silva, A.M.; Bezerra, J.A.B.; Souza, A.L.P.; Lima, G.L.; Oliveira, M.F.; Silva, A.R. Identification of ultrastructural and functional damages in sperm from six-banded armadillos (Euphractus sexcinctus) due to cryopreservation. Pesq. Vet. Bras. 2016, 36, 767–774. [Google Scholar] [CrossRef]
- Campos, L.B.; Peixoto, G.C.X.; Silva, A.M.; Souza, A.L.P.; Castelo, T.S.; Maia, K.M.; Pereira, A.F.; Silva, A.R. Estimating the binding ability of collared peccary (Pecari tajacu Linnaeus, 1758) sperm using heterologous substrates. Theriogenology 2017, 92, 57–62. [Google Scholar] [CrossRef]
- Maia, M.S.; Azevedo, H.C.; Bicudo, S.D.; Sousa, D.B.; Rodello, L. Efeito da adição do equex-STM ao diluente tris-gema na motilidade do espermatozoide criopreservado de carneiro [Effect of adding equex-STM to the tris-yolk diluent on the motility of ram cryopreserved sperm]. Acta Sci. Vet. 2008, 33, 311. [Google Scholar]
- Verstegen, J.; Iguer-Ouada, M.; Onclin, K. Computer assisted semen analyzers in andrology research and veterinary practice. Theriogenology 2002, 57, 149–179. [Google Scholar] [CrossRef] [PubMed]
- Gil, M.C.; García-Herreros, M.; Barón, F.J.; Aparicio, I.M.; Santos, A.J.; García-Marín, L.J. Morphometry of porcine spermatozoa and its functional significance in relation with the motility parameters in fresh semen. Theriogenology 2009, 71, 254–263. [Google Scholar] [CrossRef] [PubMed]
- Morris, G.I.; Morris, G.I.; Clarke, A. Effects of Low Temperature on Biological Membranes; Morris, G.I., Morris, G.I., Clarke, A., Eds.; Academic Press: New York, NY, USA, 1981; pp. 241–377. [Google Scholar]
- Schäfer-Somi, S.; Binder, C.; Burak, J.; Papadopoulos, N.; Ilas, J.; Boersma, A.; Aurich, C. Using egg yolk in a TRIS-Equex STM paste extender for freezing of dog semen is superior to egg yolk plasma, also after addition of lecithin and catalase. Cryobiology 2021, 100, 63–71. [Google Scholar] [CrossRef]
- Aboagla, E.M.E.; Terada, T. Effects of the supplementation of trehalose extender containing egg yolk with sodium do-decyl sulfate on the freezability of goat spermatozoa. Theriogenology 2004, 62, 809–818. [Google Scholar] [CrossRef] [PubMed]
- Peña, A.; Linde-Forsberg, C. Effects of Equex, one- or two-step dilution, and two freezing and thawing rates on post-thaw survival of dog spermatozoa. Theriogenology 2000, 54, 859–875. [Google Scholar] [CrossRef]
- Buranaamnuay, K.; Mahasawangkul, S.; Saikhun, K. The in vitro quality of frozen-thawed Asian elephant (Elephas maximus) spermatozoa in semen supplemented with Equex STM paste and oxytocin during and after cryopreservation. Reprod Biol. 2013, 13, 169–171. [Google Scholar] [CrossRef]
- De Mercado, E.; Rodríguez, A.; Gómez, E.; Sanz, E. Cryopreservation of Iberian pig spermatozoa. Comparison of different freezing extenders based on post-thaw sperm quality. Anim. Reprod. Sci. 2010, 118, 54–61. [Google Scholar] [CrossRef]
- Dorado, J.; Hidalgo, M.; Acha, D.; Ortiz, I.; Bottrel, M.; Azcona, F.; Carrasco, J.J.; Gómez-Arrones, V.; Demyda-Peyrás, S. Cryopreservation of Andalusian donkey (Equus asinus) spermatozoa: Use of alternative energy sources in the freezing extender affects post-thaw sperm motility patterns but not DNA stability. Anim. Reprod. Sci. 2019, 208, 106126. [Google Scholar] [CrossRef]
- Malo, C.; De Blas, I.; Skidmore, J.A. Interaction among extenders, cryoprotectants and Orvus Es Paste supplementation and freezing rate for sperm cryopreservation in the dromedary camel. Andrologia 2021, 53, e14224. [Google Scholar] [CrossRef]
- Maia, K.M.; Peixoto, G.C.X.; Campos, L.B.; Bezerra, J.A.B.; Ricarte, A.R.F.; Moreira, N.; Oliveira, M.F.; Silva, A.R. Estrus cycle monitoring of captive collared peccaries (Pecari tajacu) in semiarid conditions. Pesq. Vet. Bras. 2014, 34, 1115–1120. [Google Scholar] [CrossRef] [Green Version]
- Peixoto, G.C.X.; Lima, G.L.; Maia, K.M.; Souza, A.L.P.; Castelo, T.S.; Paiva, A.L.C.; Paula, V.V.; Oliveira, M.F.; Brito, A.B.; Domingues, S.F.S.; et al. Single injection of eCG/hCG leads to successful estrous synchronization in the collared peccary (Pecari tajacu Linnaeus, 1758). Anim. Reprod. Sci. 2019, 208, 106112. [Google Scholar] [CrossRef] [PubMed]
- Anel, L.; Gomes-Alves, S.; Alvarez, M.; Borragan, S.; Anel, E.; Nicolas, M.; Martinez-Pastor, F.; de Paz, P. Effect of basic factors of extender composition on post-thawing quality of brown bear electroejaculated spermatozoa. Theriogenology 2010, 74, 643–651. [Google Scholar] [CrossRef] [PubMed]
- Strzezek, J.; Glogowski, J.; Margierska, E.; Luberda, Z.; Jablonowska, C.Z. Some aspects of cryobiochemistry of boar semen. In Proceedings of the Tenth International Congress on Animal Reproduction and Artificial Insemination, University of Illinois, Urbana-Champaign, IL, USA, 10–14 June 1984; Volume 2, pp. 244–246. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Sperm Kinetic Motility Patterns | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Treatments | Time | Total Motility (%) | Progressive Motility (%) | VAP (µm/s) | VSL (µm/s) | VCL (µm/s) | ALH (µm/s) | BCF (Hz) | STR (%) | LIN (%) |
| Fresh | 87.8 ± 4.4 | 71.2 ± 4.6 | 57.4 ± 5.1 | 45.0 ± 4.4 | 110.0 ± 8.8 | 5.6 ± 0.3 | 33.4 ± 1.0 | 76.1 ± 1.1 | 41.3 ± 1.7 | |
| Number of Bound Sperm | Frozen–Thawed Semen | ||||
|---|---|---|---|---|---|
| Negative Control | Equex | SDS 0.1% | SDS 0.3% | SDS 0.5% | |
| Mean ± SE | 138.1 ± 10.8 A | 126.1 ± 9.0 A | 131.8 ± 8.0 A | 144.3 ± 13.6 A | 131.6 ± 9.3 A |
| Min | 84.4 | 74.3 | 110.0 | 103.4 | 81.0 |
| Max | 188.7 | 174.7 | 193.6 | 147.0 | 180.2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moreira, S.S.J.; Silva, A.M.; Pereira, A.G.; Santos, R.P.; Dantas, M.R.T.; Souza-Júnior, J.B.F.; Snoeck, P.P.N.; Silva, A.R. Effect of Detergents Based on Sodium Dodecyl Sulfate on Functional Metrics of Frozen–Thawed Collared Peccary (Pecari tajacu) Semen. Animals 2023, 13, 451. https://doi.org/10.3390/ani13030451
Moreira SSJ, Silva AM, Pereira AG, Santos RP, Dantas MRT, Souza-Júnior JBF, Snoeck PPN, Silva AR. Effect of Detergents Based on Sodium Dodecyl Sulfate on Functional Metrics of Frozen–Thawed Collared Peccary (Pecari tajacu) Semen. Animals. 2023; 13(3):451. https://doi.org/10.3390/ani13030451
Chicago/Turabian StyleMoreira, Samara S. J., Andréia M. Silva, Ana G. Pereira, Romário P. Santos, Maiko R. T. Dantas, João B. F. Souza-Júnior, Paola P. N. Snoeck, and Alexandre R. Silva. 2023. "Effect of Detergents Based on Sodium Dodecyl Sulfate on Functional Metrics of Frozen–Thawed Collared Peccary (Pecari tajacu) Semen" Animals 13, no. 3: 451. https://doi.org/10.3390/ani13030451