
Follicular Atresia, Cell Proliferation, and Anti-Mullerian Hormone in Two Neotropical Primates (Aotus nancymae and Sapajus macrocephalus)
 , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Sites and Animals
2.2. Histological Procedure and Follicular Quantification
2.3. Immunohistochemistry for Caspase-3, BCL-2, PCNA, and AMH
2.4. Hormonal, Proliferative and Cell Apoptotic Quantification
- number of immunostained follicles/total follicles counted × 100;
- number of immunostained granulosa cells/total of 1000 granulosa cells analyzed;
- number of immunostained oocytes/number of oocytes counted × 100.
2.5. Statistical Analysis
3. Results
3.1. Follicular Quantification
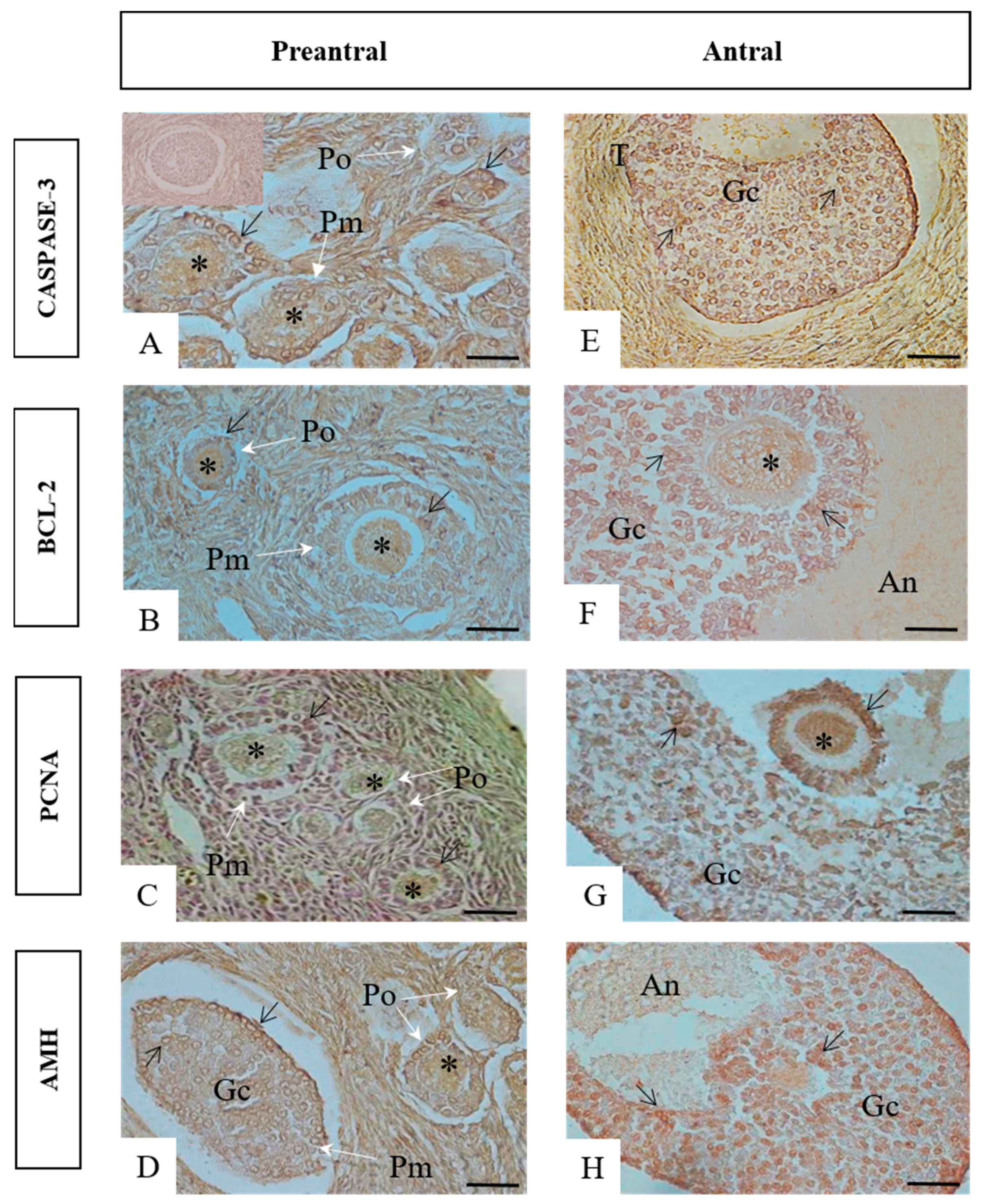
3.2. Immunohistochemistry for Cleaved Caspase-3, BCL-2, PCNA, and AMH
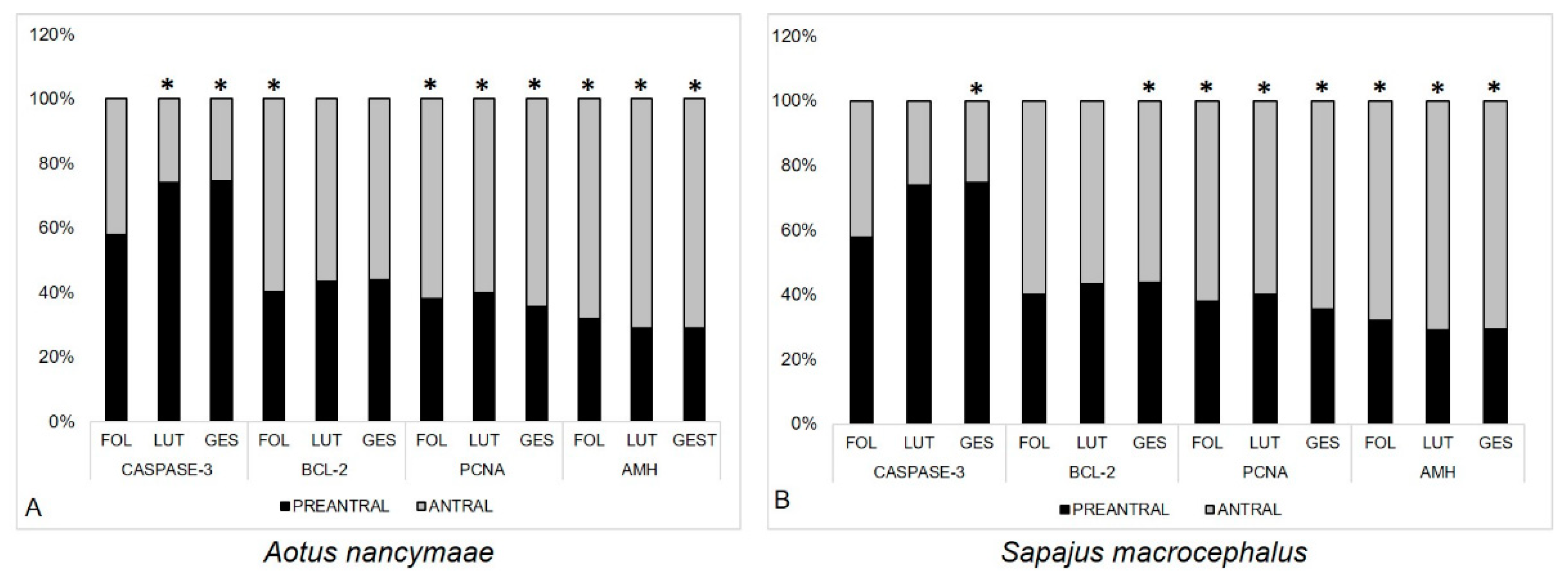
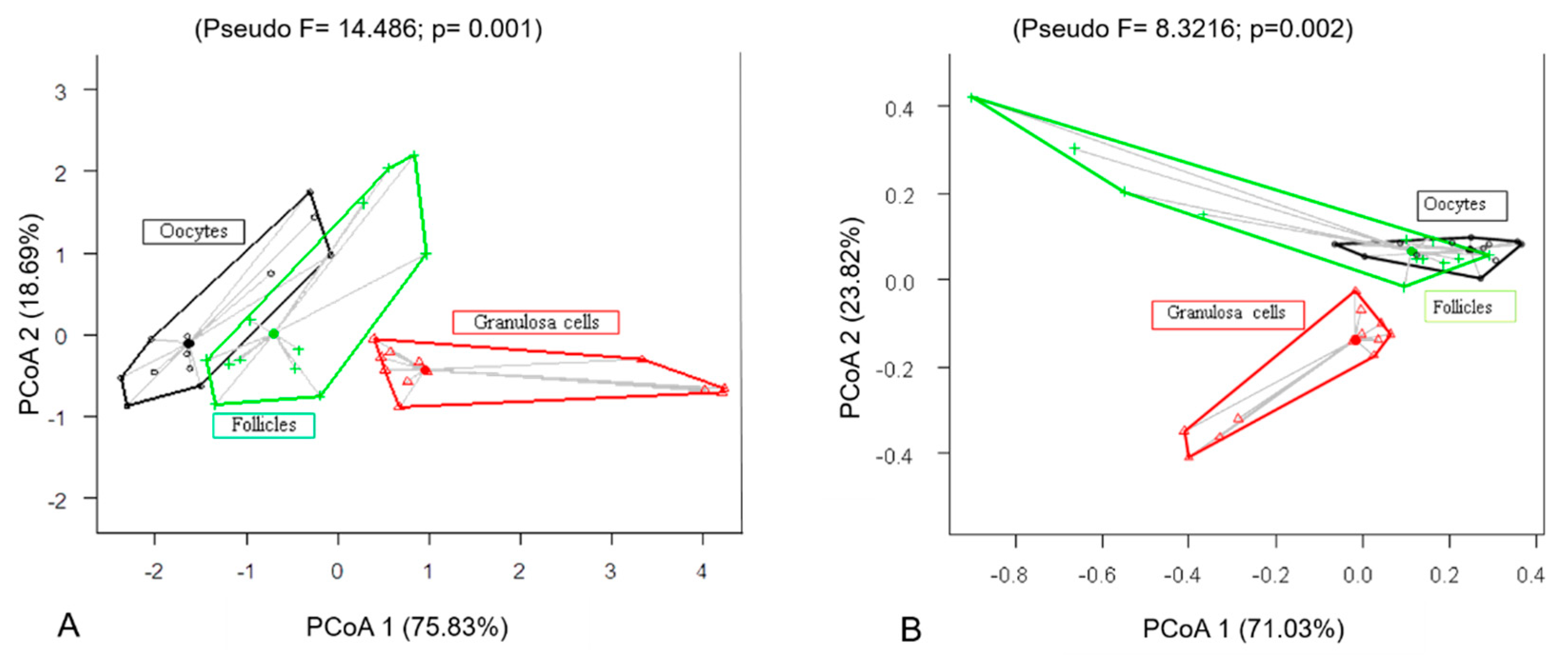
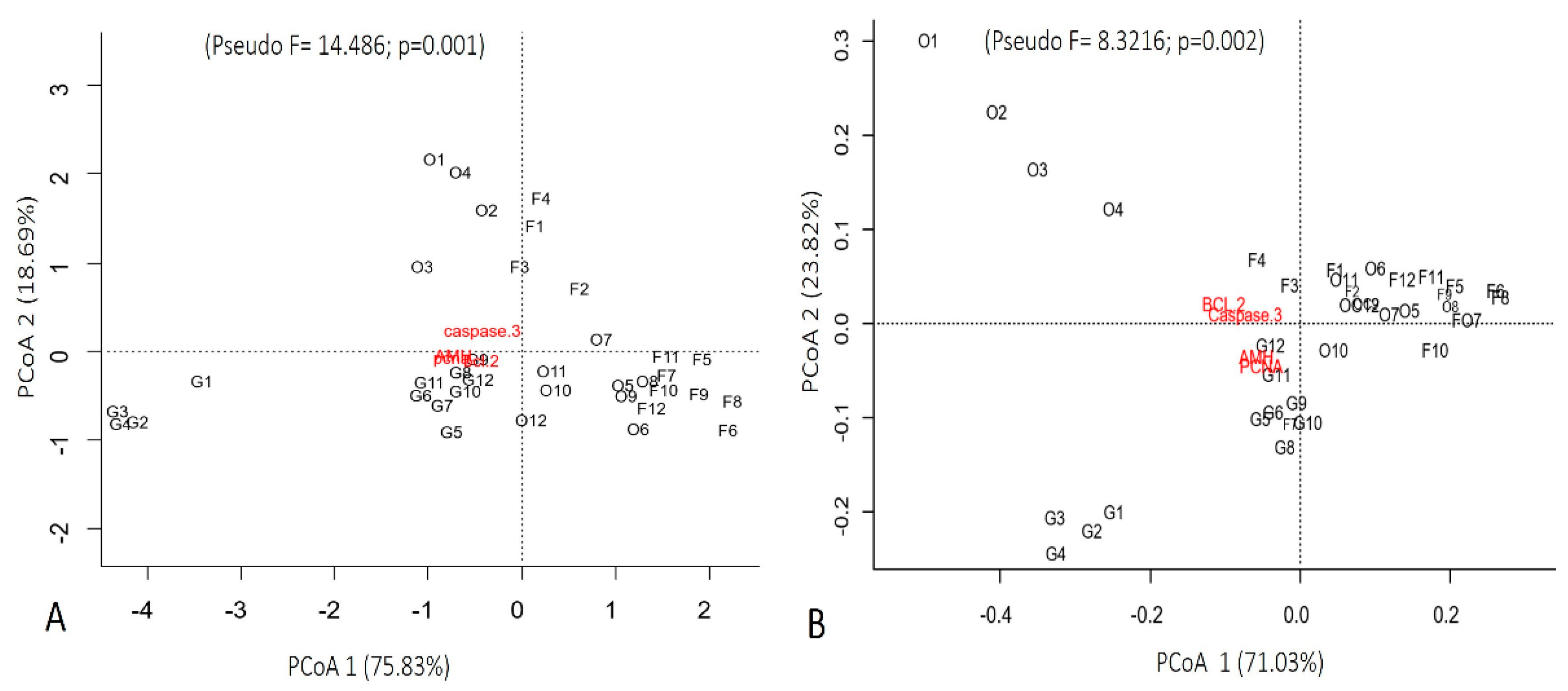
3.3. Quantification of Immunolabeled Cells for Caspase-3-Cleaved, BCL-2, PCNA, and AMH
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Coutinho, L.N.; Brito, M.B.; Monteiro, F.O.; Andrade, R.S.; Conceição, M.E.; Feliciano, M.R.; Castro, P.H.; Teixeira, P.P.; Vicente, W.R. Analysis of follicular events in owl monkeys (Aotus azarai infulatus) using B-mode and Doppler ultrasound. Theriogenology 2013, 80, 99–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayor, P.; Takeshita, R.S.C.; Coutinho, L.N.; Sanchéz, N.S.; Gálvez, H.G.; Ique, C.; Ruiz, J.C.; Monteiro, F.O.B. Functional morphology of the tubular genital organs in the female owl monkey (Aotus spp.). J. Med. Primatol. 2015, 44, 158–167. [Google Scholar] [CrossRef] [PubMed]
- Mayor, P.A.; Takeshita, R.S.C.; Coutinho, L.N.; Sánchez, N.; Gálvez, H.; Ique, C.; Monteiro, F.O.B. Ovarian function in captive owl monkeys (Aotus nancymaae and A. vociferans). J. Med. Primatol. 2015, 44, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Mayor, P.; Pereira, W.; Nacher, V.; Navarro, M.; Monteiro, F.O.B.; El Bizri, H.R.; Carretero, A. Menstrual cycle in four New World primates: Poeppig’s woolly monkey (Lagothrix poeppigii), red uakari (Cacajao calvus), large-headed capuchin (Sapajus macrocephalus) and nocturnal monkey (Aotus nancymaae). Theriogenology 2019, 123, 11–21. [Google Scholar] [CrossRef]
- Monteiro, F.O.B.; Coutinho, L.N.; Pompeu, E.S.S.; Castro, P.H.G.C.; Maia, C.E.; Pereira, W.L.A.; Vicente, W.R.R. Ovarian and uterine ultrasonography in Aotus azarai infulatus. Int. J. Primatol. 2009, 30, 327–336. [Google Scholar] [CrossRef]
- Monteiro, F.O.B.; Coutinho, L.N.; Takeshita, R.S.C.; Silva, G.A.; Silva, K.S.M.; Whiteman, C.W.; Castro, P.H.G.; Muniz, J.A.P.C.; Vicente, W.R.R. A protocol for gynecological and obstetric examination of owl monkeys using ultrasound. Rev. Ci Agra. 2011, 54, 5–11. [Google Scholar] [CrossRef]
- Lima, M.C.M.; Scalercio, S.R.R.A.; Lopes, C.T.A.; Martins, N.D.; Oliveira, K.G.; Caldas-Bussiere, M.C.; Santos, R.R.; Domingues, S.F.S. Monitoring sexual steroids and cortisol at different stages of the ovarian cycle from two capuchin monkey species: Use of non− or less invasive methods than blood sampling. Heliyon 2019, 5, e02166. [Google Scholar] [CrossRef]
- Miranda, S.A.; Leão, D.L.; Oliveira, K.G.; Sodré, I.S.; Domingues, S.F.S. Gestational ultrasonography and Dopplerfluxometry in capuchin monkeys (Sapajus apella) zoometric. Theriogenology 2018, 108, 63–73. [Google Scholar] [CrossRef]
- Fernandez-Duque, E. Owl monkeys Aotus spp. in the wild and the captivity. Int. Zoo Yearb. 2012, 46, 80–94. [Google Scholar] [CrossRef]
- Huck, M.G.; Rotundo, M.; Fernandez-Duque, E. Growth and development in wild owl monkeys (Aotus azarai) of Argentina. Int. J. Primatol. 2011, 32, 1133–1152. [Google Scholar] [CrossRef]
- Cardoso, D.L.; Guimarães, D.A.A.; Mayor, P.; Ferreira, M.A.P.; Dias, H.L.T.; Espinheiro, R.F.; Monteiro, F.O.B. Reproductive biology of owl (Aotus spp.) and capuchin (Sapajus spp.) monkeys. Anim. Reprod. Sci. 2021, 227, 106732. [Google Scholar] [CrossRef]
- Takeshita, R.S.C.; Monteiro, F.O.B.; Lins, F.L.M.L.; Andrade, R.S.; Silva, G.A.; Cardoso, A.M.C.; Pereira, W.L.A.; Rahal, S.C. Aspectos histológicos de ovários e testículos de macacos-da-noite (Aotus azarai infulatus) (Kuhl 1820). In Em: A Primatologia no Brasil, 13th ed.; Passos, S.C., Miranda, J.M.D., Eds.; Sociedade Brasileira de Primatologia: Curitiba, Brazil, 2014; Volume 13, pp. 303–314. [Google Scholar]
- Brito, A.B.; Brito, D.C.C.; Silva, W.B.; Rodrigues, A.P.R.; Figueiredo, J.R.; Domingues, S.F.S.; Santos, R.R. Morphology and morphometry of preantral follicles, and immunolocalization of angiogenic factors in ovarian tissue from the Neotropical primate Sapajus apella. Zygote 2018, 26, 424–429. [Google Scholar] [CrossRef]
- Brentnall, M.; Rodriguez-Menocal, L.; De Guevara, R.L.; Cepero, E.; Boise, L.H. Caspase-9, caspase-3 and caspase-7 have distinct roles during intrinsicapoptosis. BMC Cell Biol. 2013, 14, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, G.M. Caspases: The executioners of apoptosis. Biochem. J. 1997, 326, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jenkins, V.K.; Timmons, A.K.; Mccall, K. Diversity of cell death pathways: Insight from the fly ovary. Trends Cell Biol. 2013, 23, 567–574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Llambi, F.M.; Moldoveanu, T.; Tait, S.W.; Bouchier-Hayes, L.; Temirov, J.; Mccormick, L.L.; Dillon, C.P.; Green, D.R. A unified model of mammalian BCL-2 protein family interactions at the mitochondria. Mol. Cell. 2011, 44, 517–531. [Google Scholar] [CrossRef] [Green Version]
- Ola, M.S.; Nawaz, M.; Ahsan, H.G. Role of Bcl-2 family proteins and caspases in the regulation of apoptosis. Mol. Cell. Biochem. 2011, 351, 41–58. [Google Scholar] [CrossRef] [PubMed]
- Stoimenov, I.; Helleday, T. PCNA on the crossroad of cancer. Biochem. Soc. Trans. 2009, 37, 605–613. [Google Scholar] [CrossRef] [Green Version]
- Xu, B.; Hua, J.; Zhang, Y.; Jiang, X.; Zhang, H.; Ma, T.; Zheng, W.; Sun, R.; Shen, W.; Sha, J.; et al. Proliferating cell nuclear antigen (PCNA) regulates primordial follicle assembly by promoting apoptosis of oocytes in fetal and neonatal mouse ovaries. PLoS ONE 2011, 6, e16046. [Google Scholar] [CrossRef] [Green Version]
- Wandji, S.A.; Srsen, V.; Nathanielsz, P.W.; Eppig, J.J.; Fortune, J.E. Initiation of growth of baboon primordial follicles in vitro. Hum. Reprod. 1997, 12, 1993–2001. [Google Scholar] [CrossRef] [Green Version]
- Durlinger, A.L.; Gruijters, M.J.; Kramer, P.; Karels, B.; Kumar, T.R.; Matzuk, M.M.; Rose, U.M.; De Jong, F.H.; Uilenbroek, J.T.; Grootegoed, J.A.; et al. Anti-Müllerian hormone attenuates the effects of FSH on follicle development in the mouse ovary. Endocrinology 2001, 142, 4891–4899. [Google Scholar] [CrossRef] [PubMed]
- Modi, D.; Bhartiya, D.; Puri, C. Developmental expression and cellular distribution of Mullerian inhibiting substance in the primate ovary. Reproduction 2006, 132, 443–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersen, C.Y.; Byskov, A.G. Estradiol and regulation of anti-Mullerian hormone, inhibin-A, and inhibin-B secretion: Analysis of small antral and preovulatory human follicles’ fluid. J. Clin. Endocrinol. Metab. 2006, 91, 4064–4069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gougeon, A.; Chainy, G.B.N. Morphometric studies of small follicles in ovaries of women at different ages. J. Reprod. Fertil. 1987, 81, 433–442. [Google Scholar] [CrossRef]
- Garcia-Gasca, A.; Leal-Tarin, B.; Rios-Sicairos, J.; Hernandez-Cornejo, R.; Aguilar-Zarate, G.; Betancourt-Lozano, M. Follicular apoptosis in the mussel (Mytella strigata) as potential indicator of environmental stress in coastal ecosystems. J. Environ. Sci. Health A Tox. Hazard Subst. Environ. Eng. 2010, 45, 56–61. [Google Scholar] [CrossRef]
- Pantoja, J.C.D.; Oliveira, L.F.S.; Ferreira, M.A.P.; Silva, B.R.M.; Nunes, Z.M.P.; Mendes, Y.A.; Oliveira, R.S.; Rocha, R.M. Salinity and rainfall as inducers of cell proliferation and apoptosis in mangrove oyster Crassostrea gasar spermatogenesis. Reg. Stud. Mar. Sci. 2020, 39, 101411. [Google Scholar] [CrossRef]
- Legendre, P.; Legendre, L. Transformations for Community Composition Data, 3rd ed.; Numer Ecol; MCM Publishing: Kindlington, UK, 2012. [Google Scholar]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral Ecol. 2001, 26, 32–46. [Google Scholar]
- Anderson, M.J. PERMANOVA: A FORTRAN Computer Program for Permutational Multivariate Analysis of Variance; Department of Statistics University of Auckland: Auckland, New Zealand, 2005. [Google Scholar]
- Gougeon, A.; Ecochard, R.; Thalabard, J.C. Age-related changes of the population of human ovarian follicles: Increase in the disappearance rate of non-growing and early-growing follicles in aging women. Biol. Reprod. 1994, 50, 653–663. [Google Scholar] [CrossRef]
- Domingues, S.F.S.; Diniz, L.V.; Furtado, S.H.C.; Ohashi, O.M.; Rondina, D.; Silva, L.D.M. Histological study of capuchin monkey (Cebus apella) ovarian follicles. Acta Amaz. 2004, 34, 495–501. [Google Scholar] [CrossRef]
- Forabosco, A.; Sforza, C. Establishment of ovarian reserve: A quantitation morphometric study of the developing human ovary. Fertil. Steril. 2007, 88, 675–683. [Google Scholar] [CrossRef]
- Baker, T.G. A quantitative and cytological study of germ cells in human ovaries. Proc. R. Soc. Lond. B Biol. Sci. 1963, 158, 417–433. [Google Scholar] [CrossRef]
- Faddy, M.J.; Gosden, R.G. A mathematical model of follicle dynamics in the human ovary. Hum. Reprod. 1995, 10, 770–775. [Google Scholar] [CrossRef]
- Koering, M.J. Preantral follicle development during the menstrual cycle in the Macaca mulatta ovary. Am. J. Anat. 1983, 166, 429–443. [Google Scholar] [CrossRef]
- Miller, P.B.; Charleston, J.S.; Battaglia, D.E.; Klein, N.A.; Soules, M.R. Morphometric analysis of primordial follicle number in pigtailed monkey ovaries: Symmetry and relationship with age. Biol. Reprod. 1999, 61, 553–556. [Google Scholar] [CrossRef] [Green Version]
- Lanna, M.F. Estudo Morfométrico do Desenvolvimento Follicular Ovariano de Camundongas Albino Suíço. Master’s Thesis, Universidade Federal de Viçosa, Viçosa, Brazil, 2009. [Google Scholar]
- Boone, D.L.; Tsang, B.K. Caspase-3 in the rat ovary: Localization and possible role in follicular atresia and luteal regression. Biol. Reprod. 1998, 58, 1533–1539. [Google Scholar] [CrossRef]
- Van Nassauw, L.; Tao, L.; Harrisson, F. Distribution of apoptosis-related proteins in the quail ovary during folliculogenesis: BCL-2, BAX and CPP32. Acta Histochem. 1999, 101, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Glamočlija, V.; Vilović, K.; Saraga-Babić, M.; Baranović, A.; Sapunar, D. Apoptosis and active caspase-3 expression in human granulosa cells. Fertil. Steril. 2005, 83, 426–431. [Google Scholar] [CrossRef] [PubMed]
- Escobar, M.L.; Echeverría, O.M.; Casasa, A.S.; Garcia, G.; Aguilar, S.J.; Vázquez-Nino, G.H. Involvement of pro-apoptotic and pro-autophagic proteins in granulosa cell death. Cell Biol. 2013, 1, 9–17. [Google Scholar]
- Lombardi, L.A.; Simões, R.S.; Maganhin, C.C.; Baracat, M.C.P.; Silva-Sasso, G.R.; Florencio-Silva, R.; Soares, J.M., Jr.; Baracat, E.C. Immunohistochemical evaluation of proliferation, apoptosis and steroidogenic enzymes in the ovary of rats with polycystic ovary. Rev. Assoc. Med. Bras. 2014, 60, 349–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salvetti, N.R.; Panzani, C.G.; Gimeno, E.J.; Neme, L.G.; Alfaro, N.S.; Ortega, H.H. An imbalance between apoptosis and proliferation contributes to follicular persistence in polycystic ovaries in rats. Reprod. Biol. Endocrinol. 2009, 7, 68. [Google Scholar] [CrossRef] [Green Version]
- Palumbo, A.; Yeh, J. In situ localization of apoptosis in the rat ovary during follicular atresia. Biol. Reprod. 1994, 51, 888–895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kugu, K.; Ratts, V.S.; Piquette, G.N.; Tilly, K.I.; Tao, X.J.; Martimbeau, S.; Aberdeen, G.W.; Krajewski, S.; Reed, J.C.; Pepe, G.J.; et al. Analysis of apoptosis and expression of bcl-2 gene family members in the human and baboon ovary. Cell Death Differ. 1998, 5, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Tilly, J.L.; Tilly, K.I.; Perez, G.I. The genes of cell death and cellular susceptibility to apoptosis in the ovary: A hypothesis. Cell Death Differ. 1997, 4, 180–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morita, Y.; Perez, G.I.; Maravei, D.V.; Tilly, K.I.; Tilly, J.L. Targeted expression of bcl-2 in mouse oocytes inhibits ovarian follicle atresia and prevents spontaneous and chemotherapy-induced oocyte apoptosis in vitro. Mol. Endocrinol. 1999, 13, 841–850. [Google Scholar] [CrossRef]
- Gompel, A.; Sabourin, J.C.; Martin, A.; Yaneva, H.; Audoin, J.; Decroix, Y.; Poitout, P. Bcl-2 experience in normal endometrium during the menstrual cycle. Am. J. Pathol. 1994, 144, 1135–1202. [Google Scholar]
- Sabourin, J.C.; Martin, A.; Baruch, J.; Truc, J.B.; Gompel, A.; Poitout, P. Bcl-2 expression in normal breast tissue during the menstrual cycle. Int. J. Cancer 1994, 59, 1–6. [Google Scholar] [CrossRef]
- Hirshfield, A.N. Development of follicles in the mammalian ovary. Int. Rev. Cytol. 1991, 124, 43–101. [Google Scholar]
- Korsmeyer, S.J. Bcl-2 initiates a new category of oncogenes: Regulators of cell death. Blood 1992, 80, 879–886. [Google Scholar] [CrossRef] [Green Version]
- Ratts, V.S.; Flaws, J.A.; Kolp, R.; Sorenson, C.M.; Tilly, J.L. Ablation of bcl2 gene expression decreases the numbers of oocytes and primordial follicles established in the post-natal female mouse gonad. Endocrinology 1995, 36, 3665–3668. [Google Scholar] [CrossRef]
- Oktay, K.; Schenken, R.S.; Nelson, J.F. Proliferating cell nuclear antigen marks the initiation of follicular growth in the rat. Biol. Reprod. 1995, 53, 295–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tománek, M.; Chronowska, E. Immunohistochemical localization of proliferating cell nuclear antigen (PCNA) in the pig ovary. Folia Histochem. Cytobiol. 2006, 44, 269–274. [Google Scholar] [PubMed]
- Downey, K.M.; Tan, C.K.; So, A.G. DNA polymerase delta: A second eukaryotic DNA replicase. BioEssays 1990, 12, 231–236. [Google Scholar] [CrossRef] [PubMed]
- Carlsson, I.B.; Laitinen, M.P.; Scott, J.E.; Louhio, H.; Velentzis, L.; Tuuri, T.; Aaltonen, J.; Ritvos, O.; Winston, R.M.; Hovatta, O. Kit ligand and c-Kit are expressed during early human ovarian follicular development and their interaction is required for the survival of follicles in long-term culture. Reproduction 2006, 131, 641–649. [Google Scholar] [CrossRef]
- Weenen, C.; Laven, J.S.E.; Von Bergh, A.R.M.; Cranfield, M.; Groome, N.P.; Visser, J.A.; Kramer, P.; Fauser, B.C.; Themmen, A.P.N. Anti-Müllerian hormone expression pattern in the human ovary: Potential implications for initial and cyclic follicle recruitment. Mol. Hum. Reprod. 2004, 10, 77–83. [Google Scholar] [CrossRef]
- Durlinger, A.L.; Kramer, P.; Karels, B.; De Jong, F.H.; Uilenbroek, J.T.; Grootegoed, J.A.; Themmen, A.P. Control of primordial follicle recruitment by anti-Mullerian hormone in the mouse ovary. Endocrinology 1999, 140, 5789–5796. [Google Scholar] [CrossRef]
- Durlinger, A.L.; Gruijters, M.J.; Kramer, P.; Karels, B.; Ingraham, H.A.; Nachtigal, M.W.; Uilenbroek, J.T.; Grootegoed, J.A.; Themmen, A.P. Anti-Mullerian hormone inhibits initiation of primordial follicle growth in the mouse ovary. Endocrinology 2002, 143, 1076–1084. [Google Scholar] [CrossRef]
- Durlinger, A.L.; Visser, J.A.; Themmen, A.P. Regulation of ovarian function: The role of anti-Mullerian hormone. Reproduction 2002, 124, 601–609. [Google Scholar] [CrossRef]
- Thomas, F.H.; Telfer, E.E.; Fraser, H.M. Expression of anti-Mullerian hormone protein during early follicular development in the primate ovary in vivo is influenced by suppression of gonadotropin secretion and inhibition of vascular endothelial growth factor. Endocrinology 2007, 148, 2273–2281. [Google Scholar] [CrossRef] [Green Version]
- Scalercio, S.R.R.A.; Brito, A.B.; Domingues, S.F.S.; Santos, R.R.; Amorim, C.A.I. Immunolocalization of growth, inhibitory, and proliferative factors involved in initial ovarian folliculogenesis from adult common squirrel monkey (Saimiri collinsi). Reprod. Sci. 2015, 22, 68–74. [Google Scholar] [CrossRef] [Green Version]
- Walker, M.L.; Anderson, D.C.; Herndon, J.G.; Walker, L.C. Ovarian aging in squirrel monkeys (Saimiri sciureus). Reproduction 2009, 138, 793–799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amorim, C.A.; Jacobs, S.; Devireddy, R.V.; Van Langendonckt, A.V.; Vanacker, J.; Jaeger, J.; Luyckx, V.; Donnez, J.; Dolmans, M.M. Successful vitrification and autografting of baboon (Papio anubis) ovarian tissue. Hum. Reprod. 2013, 28, 2146–2156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bancsi, L.F.; Broekmans, F.J.M.; Eijkemans, M.J.C.; De Jong, F.H.; JDF Habbema, J.D.F.; Te Velde, E.R. Predictors of poor ovarian response in vitro fertilization: A prospective study comparing basal markers of ovarian reserve. Fertil. Steril. 2002, 77, 328–336. [Google Scholar] [CrossRef] [PubMed]
- Broer, S.L.; Broekmans, F.J.; Laven, J.S.; Fauser, B.C. Anti-Müllerian hormone: Ovarian reserve testing and its potential clinical implications. Hum. Reprod. Update 2014, 20, 688–701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, T.H.; Liu, C.H.; Huang, C.C.; Wu, Y.L.; Shih, Y.T.; Ho, H.N.; Yang, Y.S.; Lee, M.S. Serum anti-Müllerian hormone and estradiol levels as predictors of ovarian hyperstimulation syndrome in assisted reproduction technology cycles. Hum. Reprod. 2008, 23, 160–167. [Google Scholar] [CrossRef] [Green Version]
- Scheffer, J.B.; Lozano, D.M.; Frydman, R.; Fanchin, R. Relação entre os níveis séricos do hormônio anti-Mulleriano, inibina B, estradiol e hormônio folículo estimulante no terceiro dia e o status folicular ovariano. Rev. Bras. Ginecol. Obstet. 2007, 29, 186–191. [Google Scholar] [CrossRef]
- Takahashi, C.; Fujito, A.; Kazuka, M.; Sugiyama, R.; Ito, H.; Isaka, K. Anti-Müllerian hormone substance from follicular fluid is positively associated with success in oocyte fertilization during in vitro fertilization. Fertil. Steril. 2008, 89, 586–591. [Google Scholar] [CrossRef]
- Taketo, T.; Saeed, J.; Manganaro, T.; Takahashi, M.; Donahoe, P.K. Müllerian inhibiting substance production associated with loss of oocytes and testicular differentiation in the transplanted mouse XX gonadal primordium. Biol. Reprod. 1993, 49, 13–23. [Google Scholar] [CrossRef] [Green Version]
- Hirobe, S.; He, W.W.; Lee, M.M.; Donahoe, P.K. Mullerian inhibiting substance messenger ribonucleic acid expression in granulosa and Sertoli cells coincides with their mitotic activity. Endocrinology 1992, 131, 854–862. [Google Scholar]
- Hirobe, S.; He, W.W.; Gustafson, M.L.; Maclaughlin, D.T.; Donahoe, P.K. Müllerian inhibiting substance gene expression in the cycling rat ovary correlates with recruited or Graafian follicle selection. Biol. Reprod. 1994, 50, 1238–1243. [Google Scholar] [CrossRef] [Green Version]
- Ueno, S.; Kuroda, T.; Maclaughlin, D.T.; Ragin, R.C.; Manganaro, T.F.; Donahoe, P.K. Müllerian inhibiting substance in the adult rat ovary during various stages of the estrous cycle. Endocrinology 1989, 125, 1060–1066. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Seibel, M.M.; Maclaughlin, D.T.; Donahoe, P.K.; Ransil, B.J.; Hametz, P.A.; Richards, C.J. The inhibitory effects of Mullerian-inhibiting substance on epidermal growth factor induced proliferation and progesterone production of human granulosa-luteal cells. J. Clin. Endocrinol. Metab. 1992, 75, 911–917. [Google Scholar] [PubMed]
- Seifer, D.B.; Maclaughlin, D.T.; Penzias, A.S.; Behrman, H.R.; Asmundson, L.; Donahoe, P.K.; Haning, R.V.; Flynn, S.D. Gonadotropin-releasing hormone agonist-induced differences in granulosa cell cycle kinetics are associated with alterations in follicular fluid Müllerian-inhibiting substance and androgen content. J. Clin. Endocrinol. Metab. 1993, 76, 711–714. [Google Scholar] [PubMed] [Green Version]
- Van Houten, E.; Themmen, A.P.; Visser, J.A. Anti-Müllerian hormone (AMH): Regulator and marker of ovarian function. Ann. Endocrinol. 2010, 71, 191–197. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Follicle Structures | Ovarian Follicles | |||
|---|---|---|---|---|---|
| Preantral (Absence of Antrum) | Antral (Presence of Antrum) | ||||
| Primordial | Primary | Small | Large | ||
| Aotus nancymae | Diameter (µm) | 35.26 ± 7.13 | 109.35 ± 25.03 | 275.36 ± 67.57 | 728.51 ± 126.75 |
| Layers | 1 | 2–6 | 7–10 | >10 | |
| Sapajus macrocephalus | Diameter (µm) | 33.76 ± 2.27 | 112.30 ± 31.43 | 238.84 ± 67.48 | 858.54 ± 149.05 |
| Layers | 1 | 2–6 | 7–10 | >10 | |
| Species | Sexual Phases | Ovarian Follicles | |||
|---|---|---|---|---|---|
| Pre-Antral | Antral | ||||
| Primordial | Primary | Small | Large | ||
| Aotus nancymae | Follicular | 147.4 ± 6.6 a | 83.9 ± 6.8 ab | 17.6 ± 1.0 bc | 5.0 ± 0.1 c |
| Luteal | 107.5 ±15.7 a | 51.1 ± 1.6 ab | 14.0 ± 0.5 bc | 3.8 ± 0.1 c | |
| Gestational | 123.2 ± 6.6 a | 49.7 ± 6.2 ab | 13.7 ± 0.1 bc | 3.7 ± 0.1 c | |
| Sapajus macrocephalus | Follicular | 141.0 ± 11.2 a | 70.8 ± 1.6 ab | 16.6 ± 0.7 bc | 5.4 ± 0.1 c |
| Luteal | 104.5 ±6.5 a | 45.8 ± 2.3 ab | 13.0 ± 0.5 bc | 3.6 ± 0.07 c | |
| Gestational | 135.1 ± 8.7 a | 53.4 ± 1.8 ab | 13.4 ± 0.4 bc | 3.4± 0.06 c | |
| Species | Immunoreaction | Quantification | Sexual Phases | ||
|---|---|---|---|---|---|
| Follicular | Luteal | Gestational | |||
| Aotus nancymae | Caspase-3 | Ov. follicles | 0.64 ± 0.07 a | 0.27 ± 0.07 b | 0.30 ± 0.04 ab |
| Granulosa cells | 0.61 ± 0.02 a | 0.44 ± 0.04 b | 0.46 ± 0.02 ab | ||
| Oocytes | 0.79 ± 0.09 a | 0.33 ± 0.08 b | 0.37 ± 0.03 ab | ||
| BCL-2 | Ov. follicles | 0.23 ± 0.04 | 0.19 ± 0.03 | 0.22 ± 0.04 | |
| Granulosa cells | 0.51 ± 0.04 a | 0.36 ± 0.03 ab | 0.35 ± 0.03 b | ||
| Oocytes | 0.28 ± 0.04 | 0.23 ± 0.02 | 0.30 ± 0.07 | ||
| PCNA | Ov. follicles | 0.24 ± 0.02 a | 0.18 ± 0.01 b | 0.19 ± 0.01 ab | |
| Granulosa cells | 0.65 ± 0.04 a | 0.42 ± 0.01 ab | 0.36 ± 0.03 b | ||
| Oocytes | 0.32 ± 0.03 a | 0.24 ± 0.03 b | 0.26 ± 0.01 ab | ||
| AMH | Ov. follicles | 0.28 ± 0.03 a | 0.16 ± 0.02 b | 0.19 ± 0.03 ab | |
| Granulosa cells | 0.60 ± 0.07 a | 0.32 ± 0.01 b | 0.33 ± 0.02 ab | ||
| Oocytes | 0.32 ± 0.04 a | 0.22 ± 0.03 b | 0.25 ± 0.05 ab | ||
| Sapajus macrocephalus | Caspase-3 | Ov. follicles | 0.52 ± 0.13 a | 0.24 ± 0.04 b | 0.26 ± 0.05 ab |
| Granulosa cells | 0.55 ± 0.03 a | 0.39 ± 0.02 b | 0.37 ± 0.04 ab | ||
| Oocytes | 0.77 ± 0.09 a | 0.30 ± 0.03 b | 0.30 ± 0.02 ab | ||
| BCL-2 | Ov. follicles | 0.30 ± 0.02 a | 0.17 ± 0.01 b | 0.2 ±0.02 ab | |
| Granulosa cells | 0.55 ± 0.04 a | 0.34 ± 0.02 b | 0.36 ± 0.03 ab | ||
| Oocytes | 0.97 ± 0.24 a | 0.26 ± 0.04 b | 0.36 ± 0.01 ab | ||
| PCNA | Ov. follicles | 0.24 ± 0.01 | 0.20 ± 0.01 | 0.23 ± 0.01 | |
| Granulosa cells | 0.69 ± 0.03 a | 0.48 ± 0.02 ab | 0.38 ± 0.02 b | ||
| Oocytes | 0.28 ± 0.01 ab | 0.24 ± 0.01 a | 0.30 ± 0.03 b | ||
| AMH | Ov. follicles | 0.27 ± 0.02 a | 0.16 ± 0.01 b | 0.18 ± 0.03 ab | |
| Granulosa cells | 0.59 ± 0.04 a | 0.31 ± 0.01 b | 0.34 ± 0.03 ab | ||
| Oocytes | 0.32 ± 0.03 a | 0.19 ± 0.02 b | 0.21 ± 0.03 ab | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cardoso, D.d.L.; Guimarães, D.A.d.A.; Mayor, P.; Ferreira, M.A.P.; Coutinho, L.N.; Monteiro, F.O.B. Follicular Atresia, Cell Proliferation, and Anti-Mullerian Hormone in Two Neotropical Primates (Aotus nancymae and Sapajus macrocephalus). Animals 2023, 13, 1051. https://doi.org/10.3390/ani13061051
Cardoso DdL, Guimarães DAdA, Mayor P, Ferreira MAP, Coutinho LN, Monteiro FOB. Follicular Atresia, Cell Proliferation, and Anti-Mullerian Hormone in Two Neotropical Primates (Aotus nancymae and Sapajus macrocephalus). Animals. 2023; 13(6):1051. https://doi.org/10.3390/ani13061051
Chicago/Turabian StyleCardoso, Deise de Lima, Diva Anélie de Araújo Guimarães, Pedro Mayor, Maria Auxiliadora Pantoja Ferreira, Leandro Nassar Coutinho, and Frederico Ozanan Barros Monteiro. 2023. "Follicular Atresia, Cell Proliferation, and Anti-Mullerian Hormone in Two Neotropical Primates (Aotus nancymae and Sapajus macrocephalus)" Animals 13, no. 6: 1051. https://doi.org/10.3390/ani13061051