
Surgical Anatomy for Sterilization Procedures in Female Capybaras

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
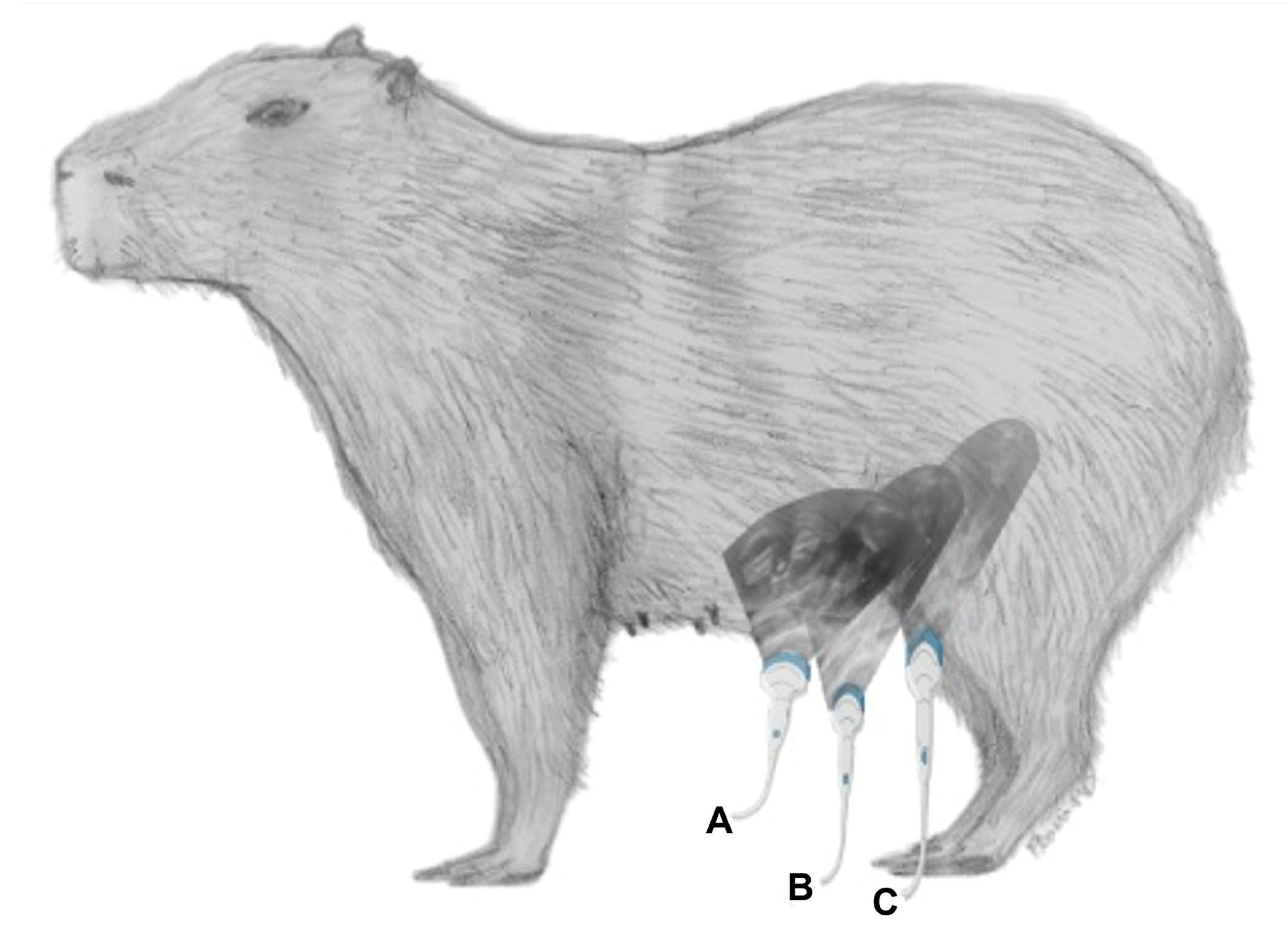
2. Materials and Methods
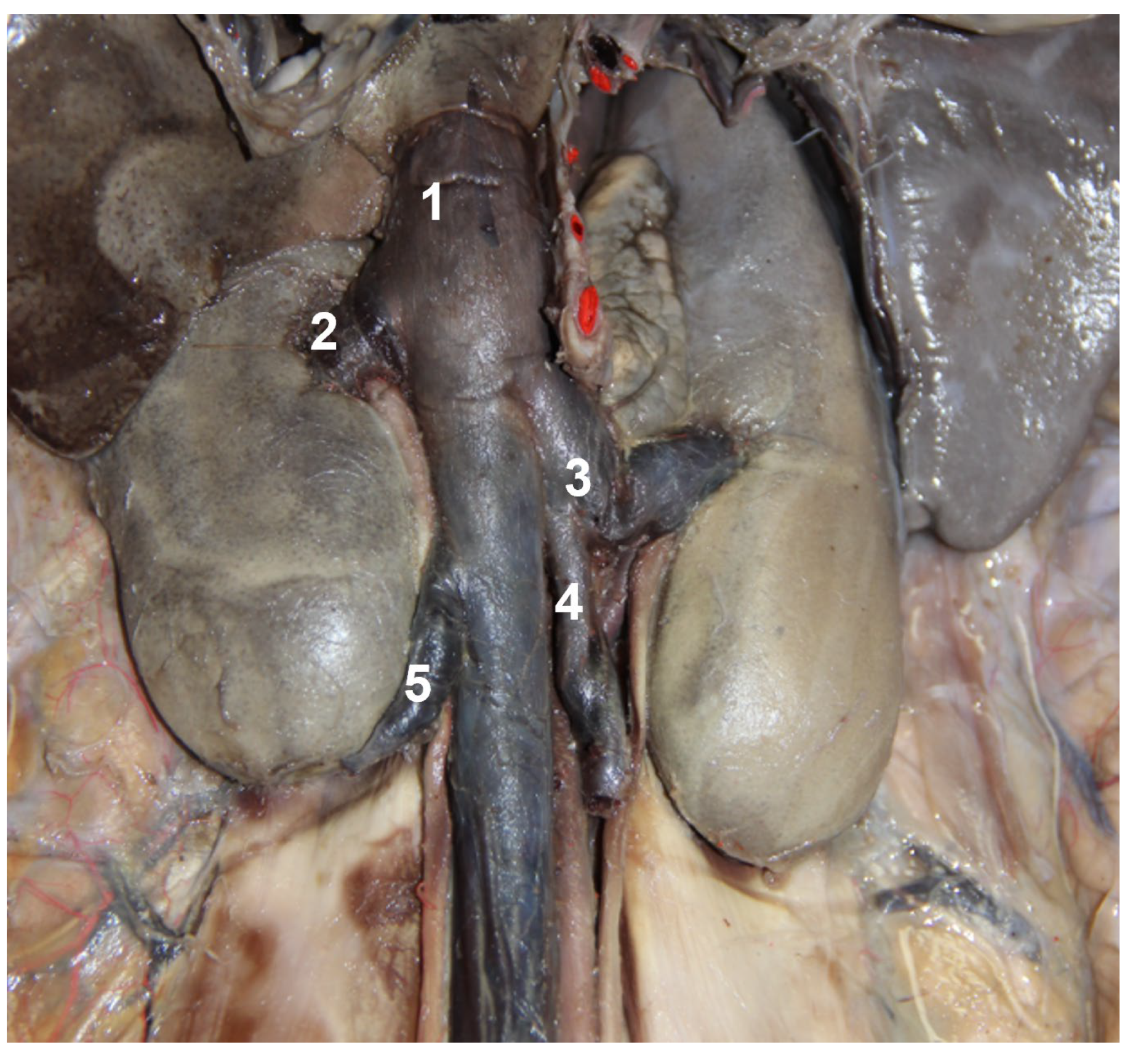
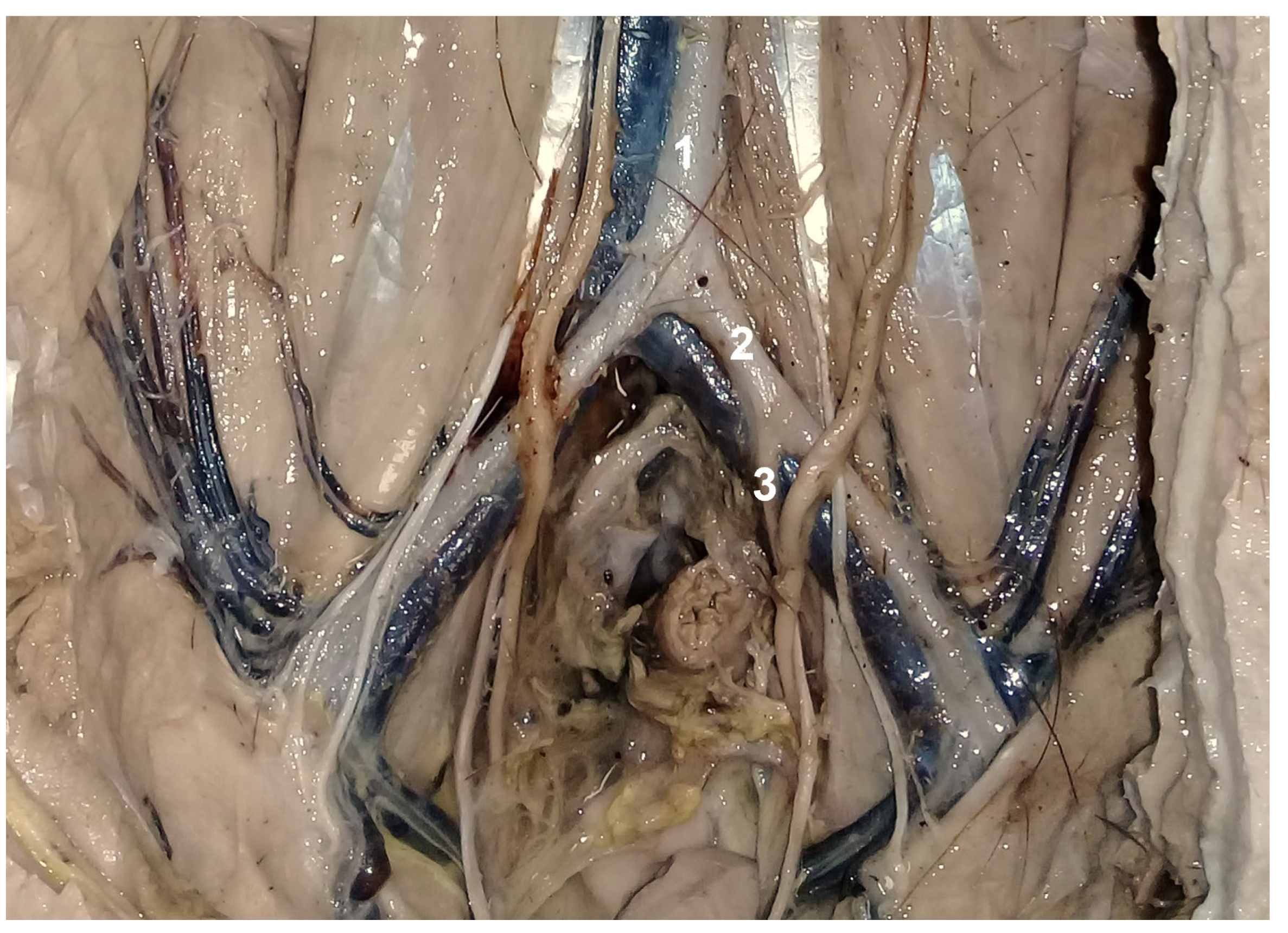
3. Results
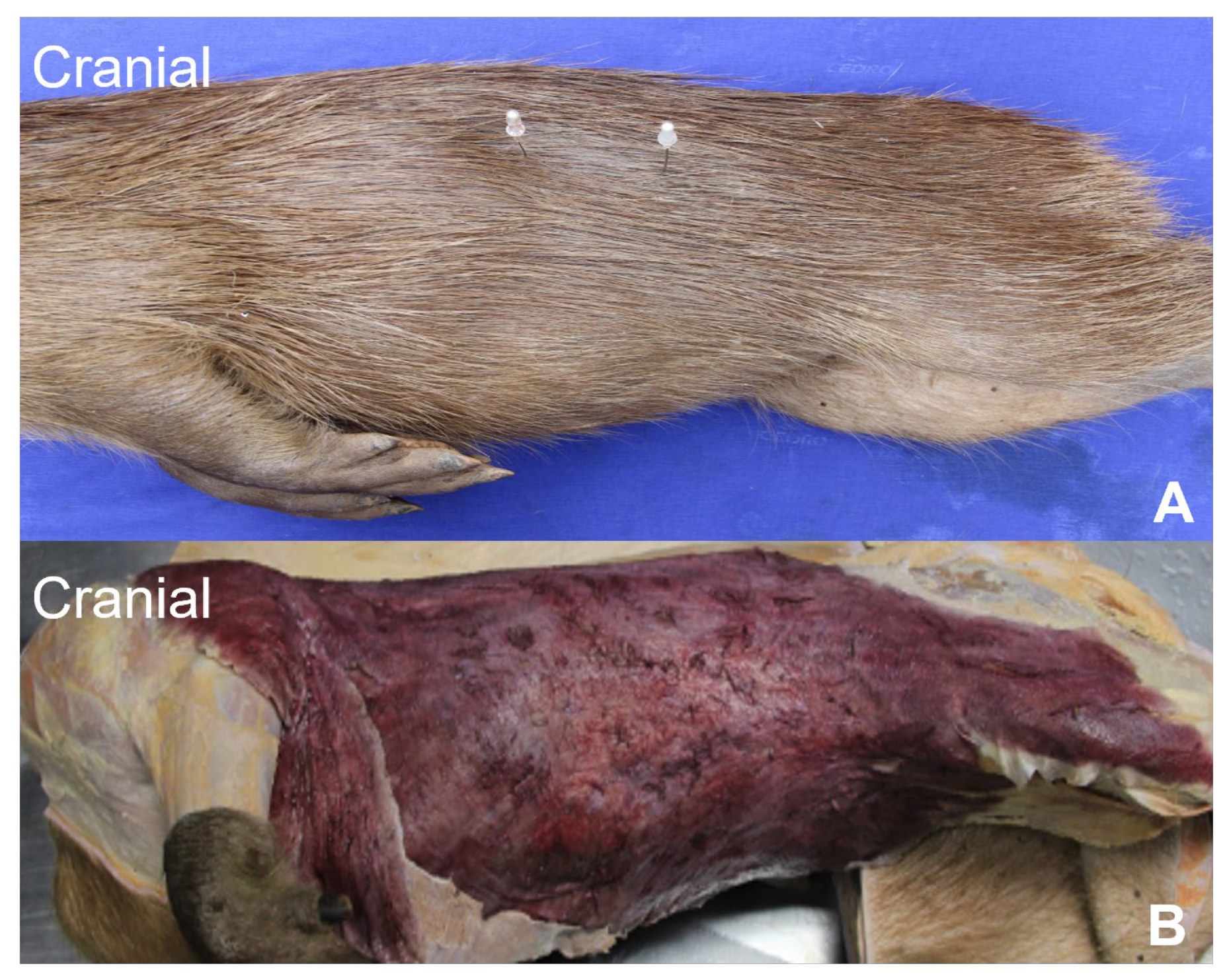
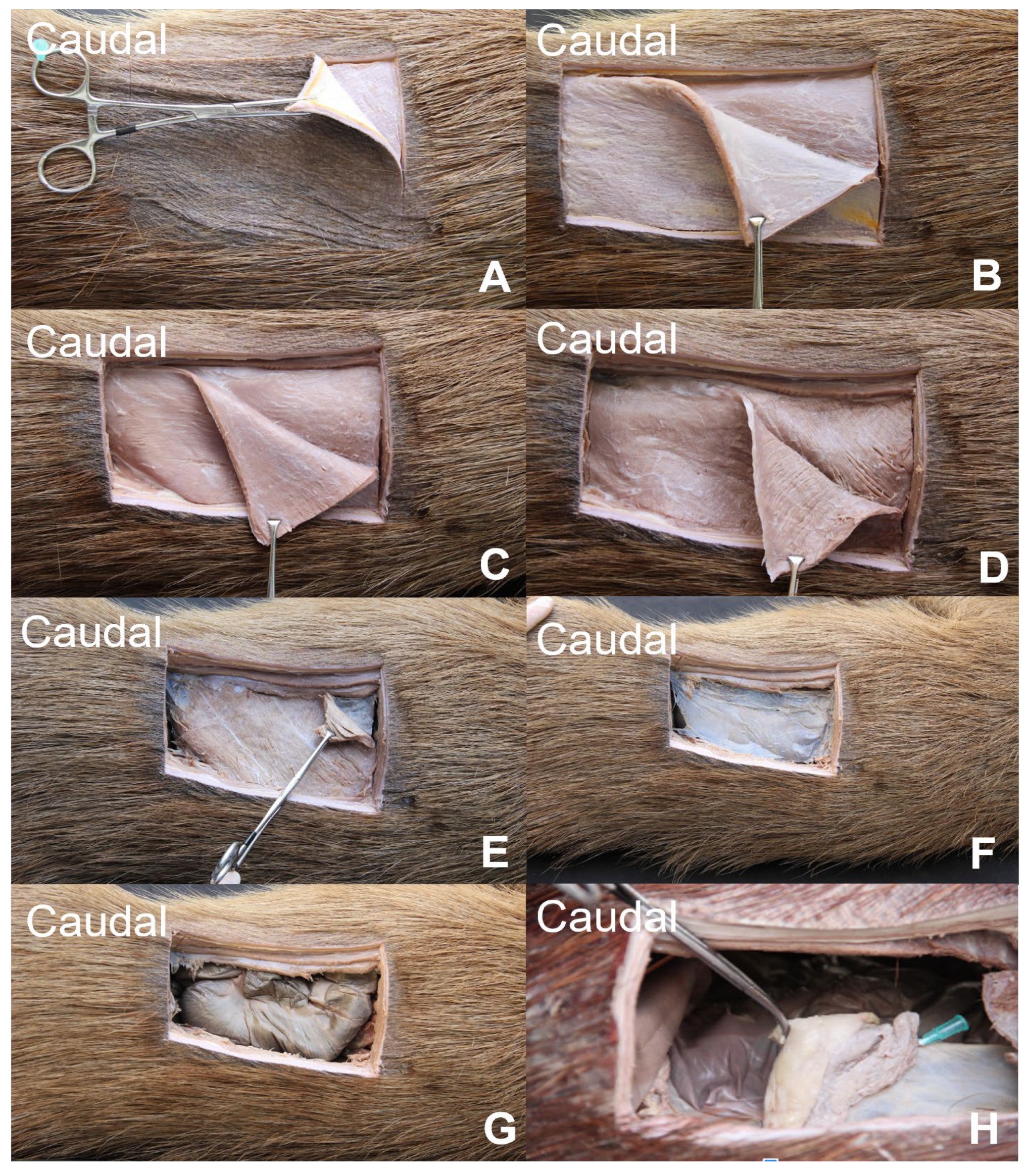
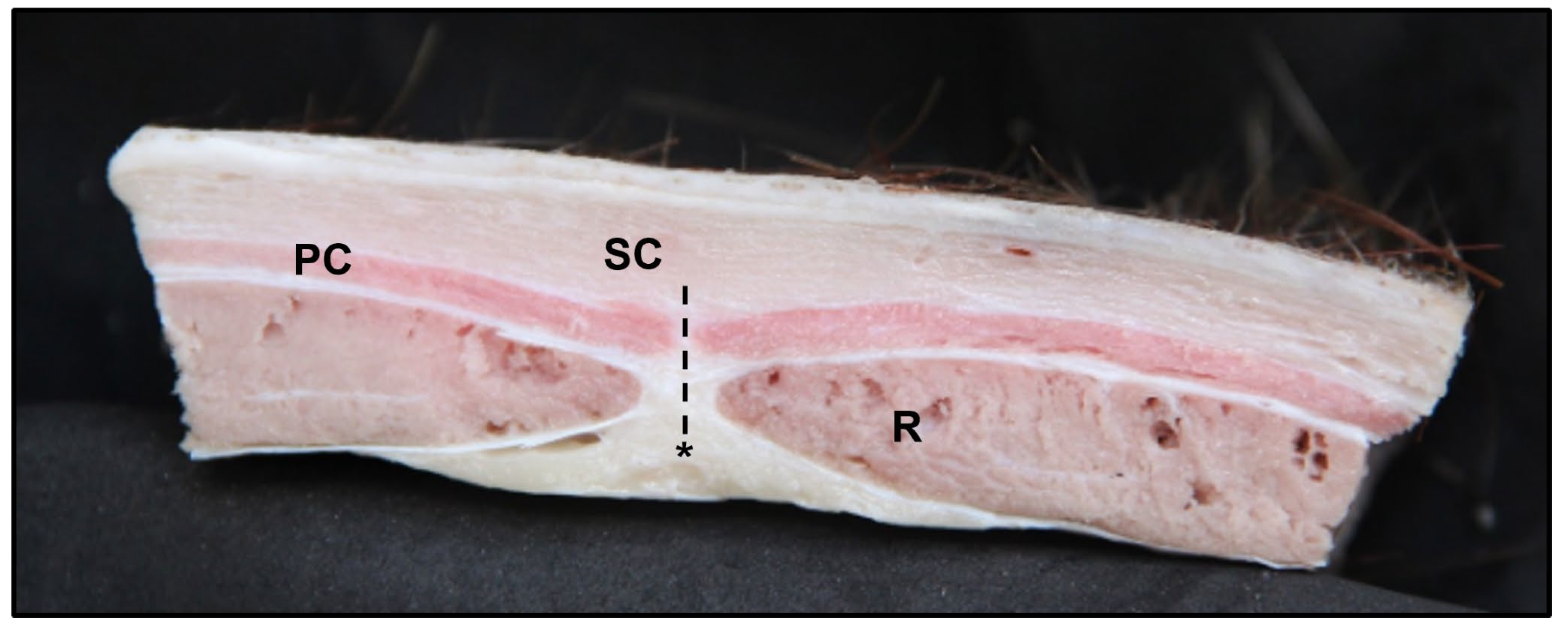
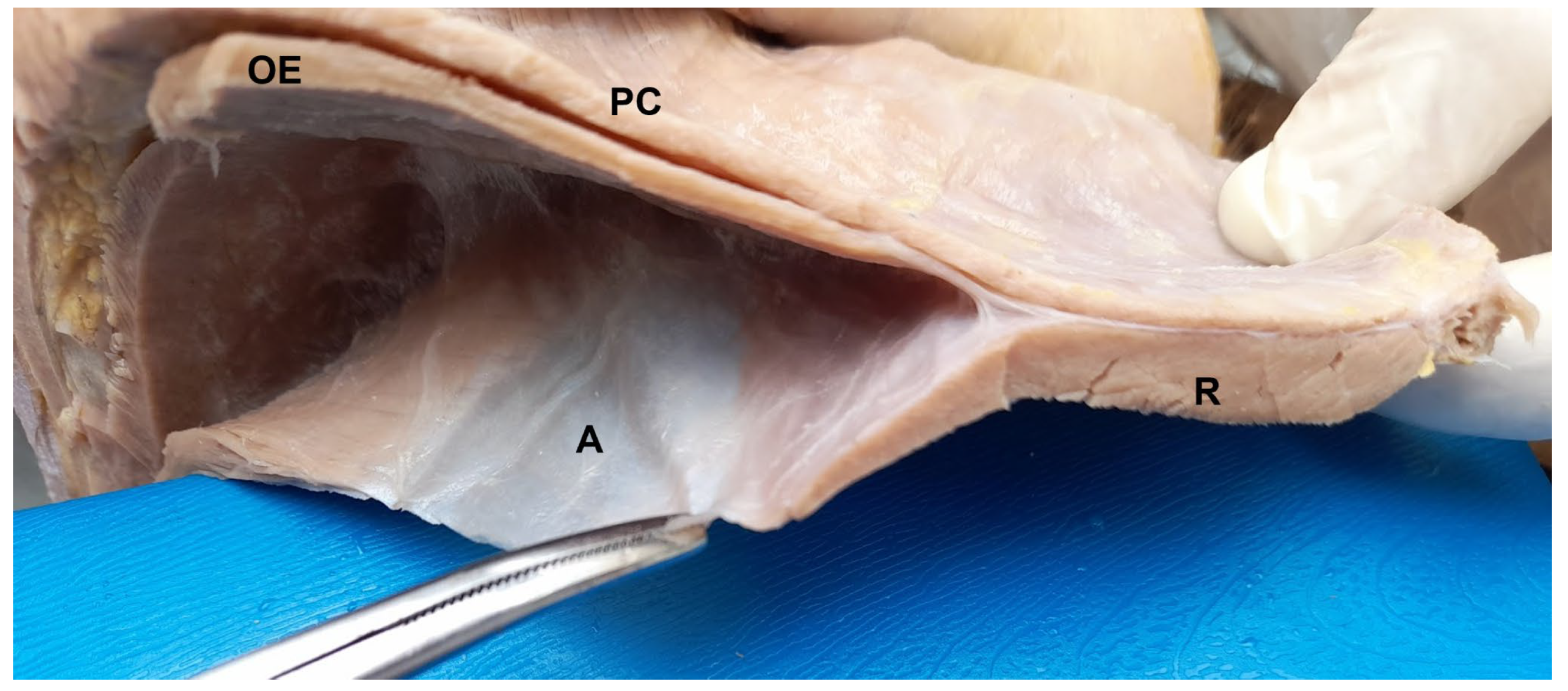
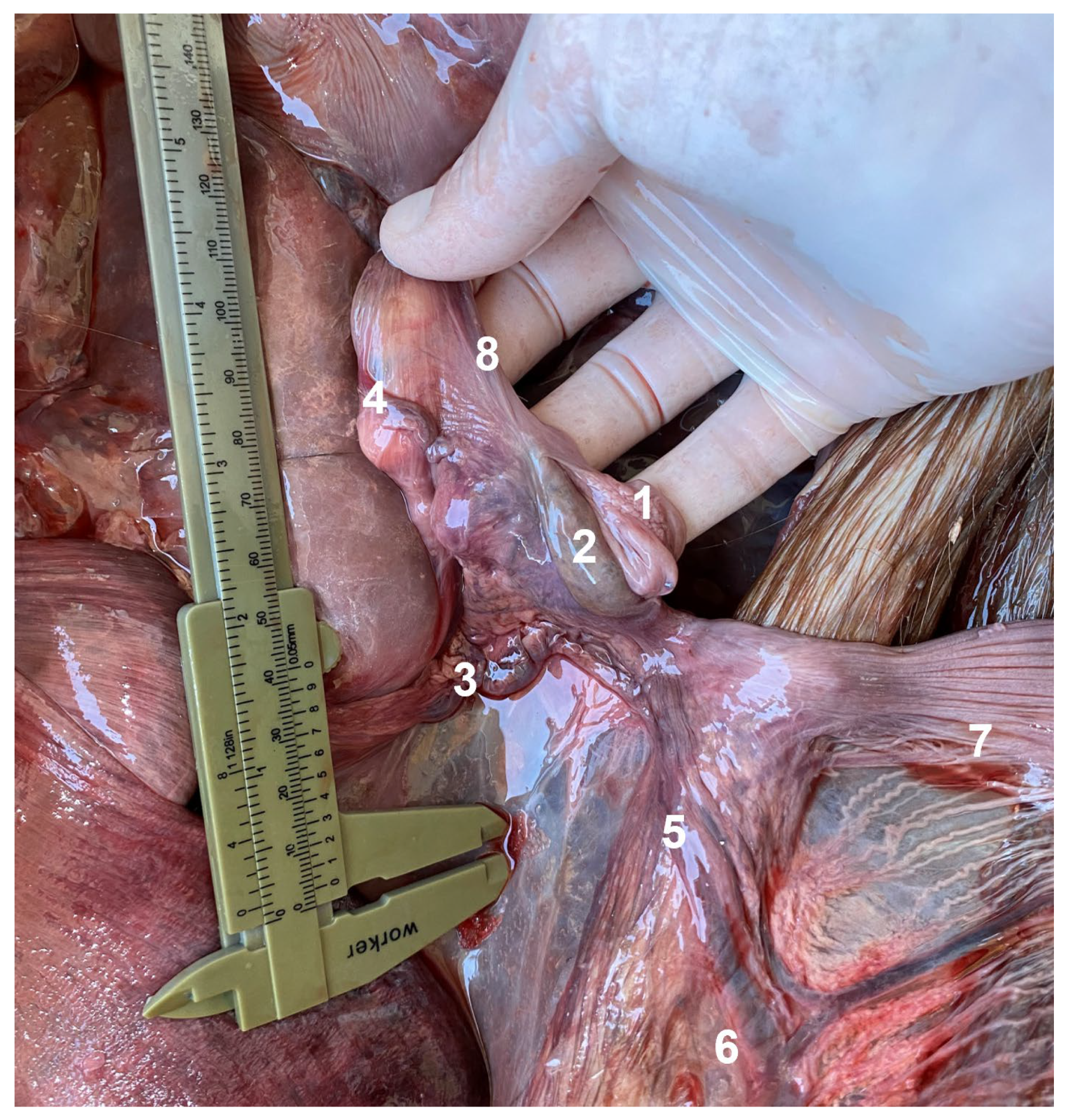
3.1. Lateral Abdominal (Flank) Dissection
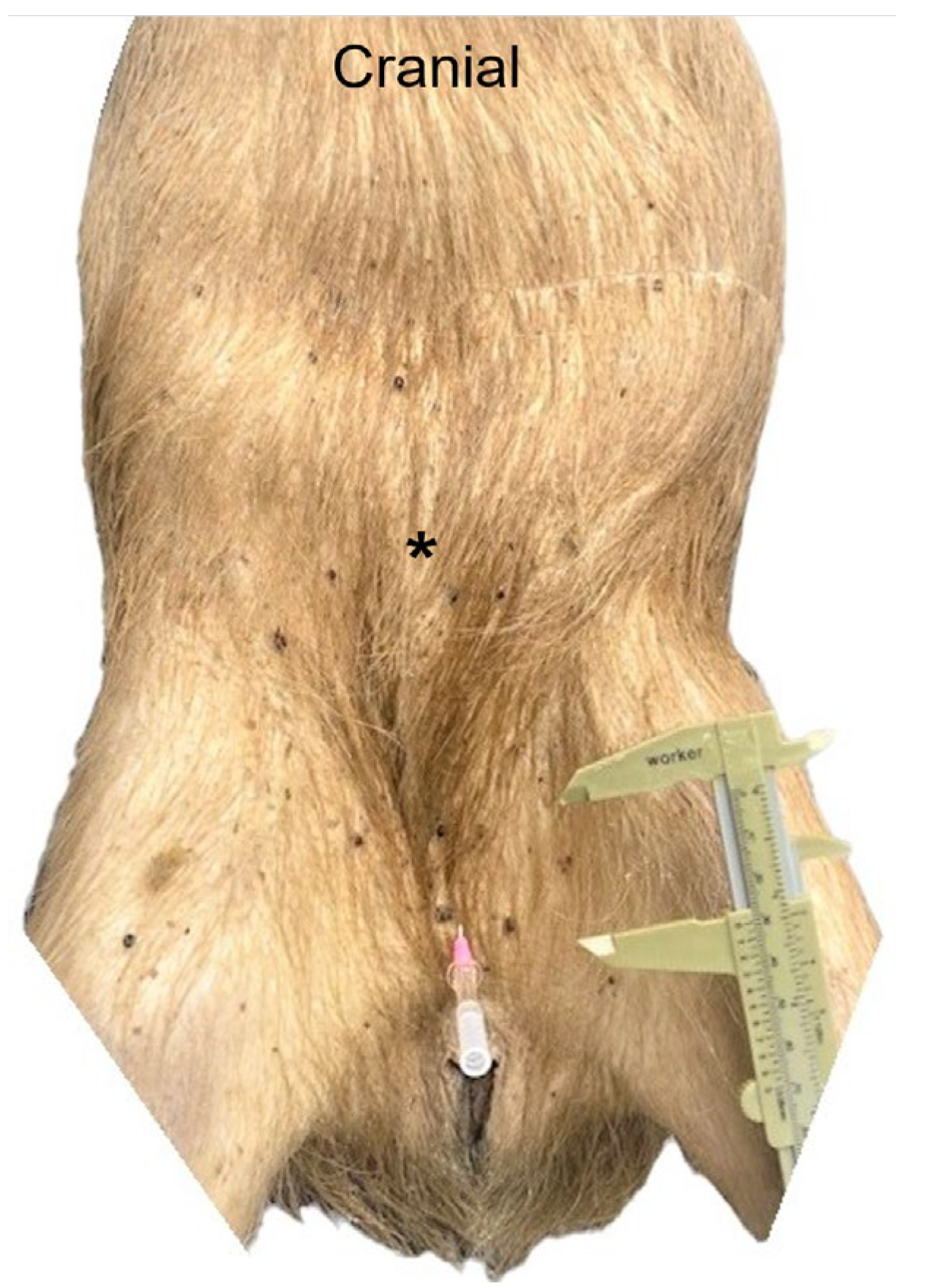
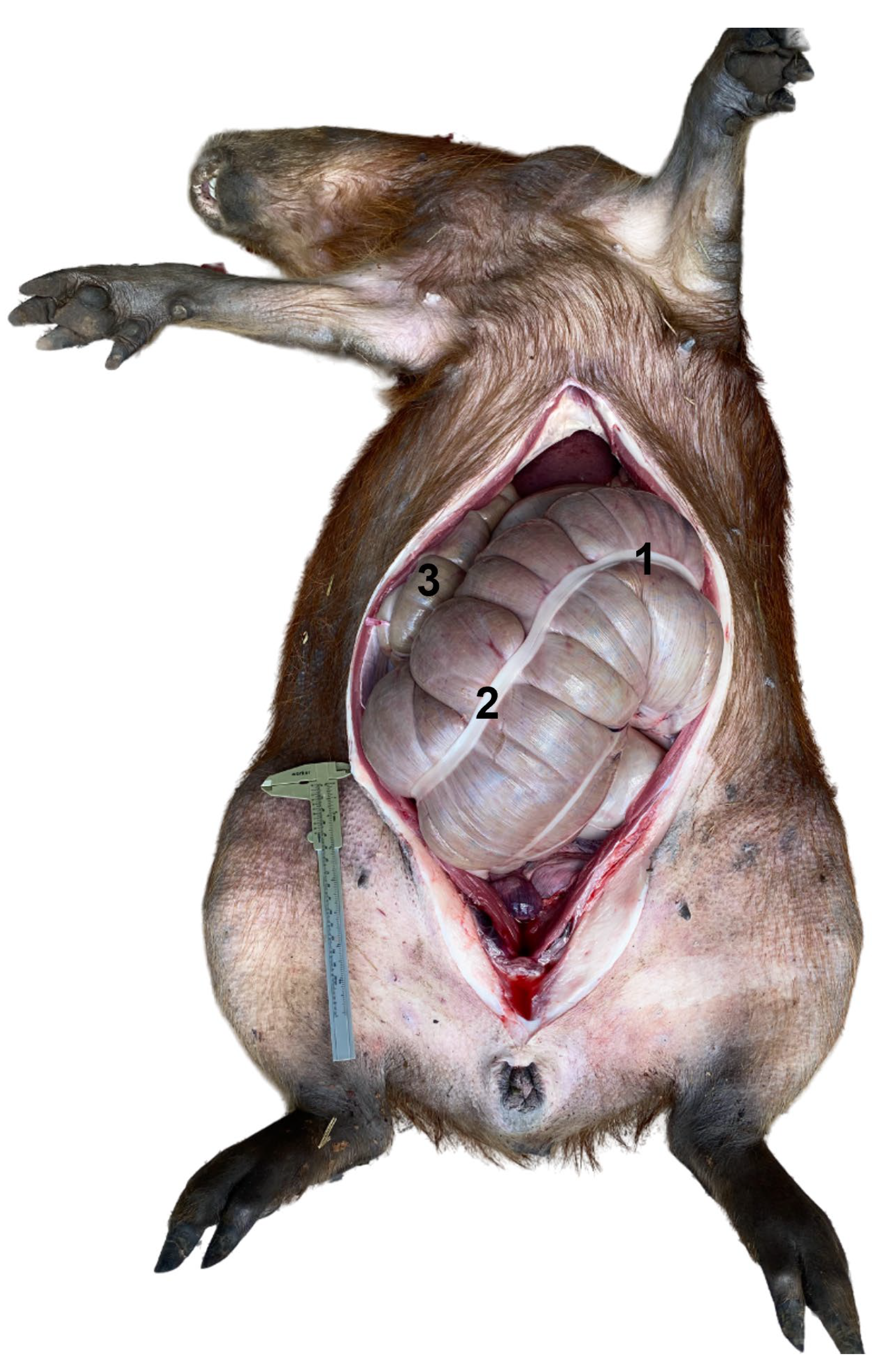
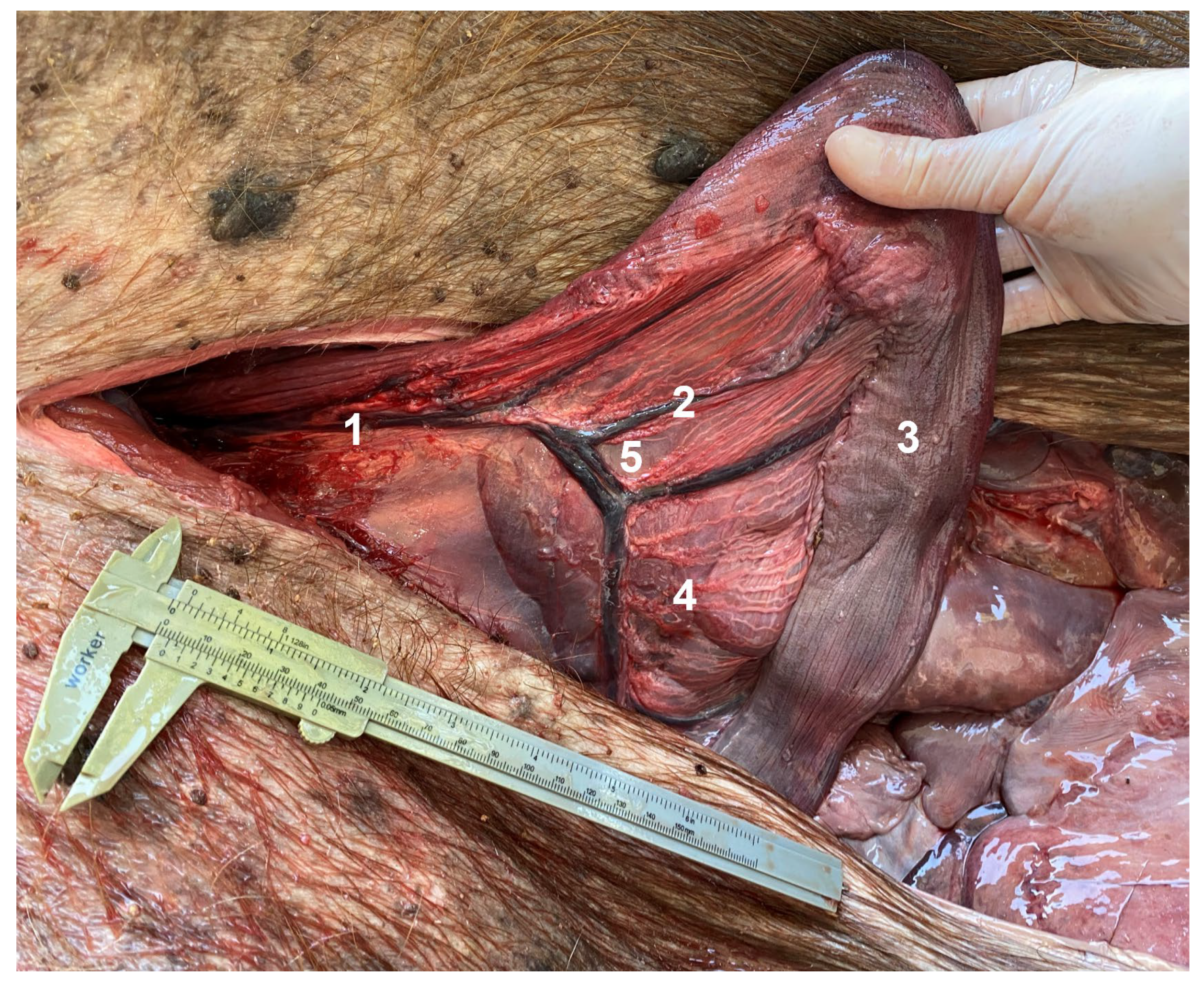
3.2. Ventral Abdominal Post-Umbilical Dissection (LA)
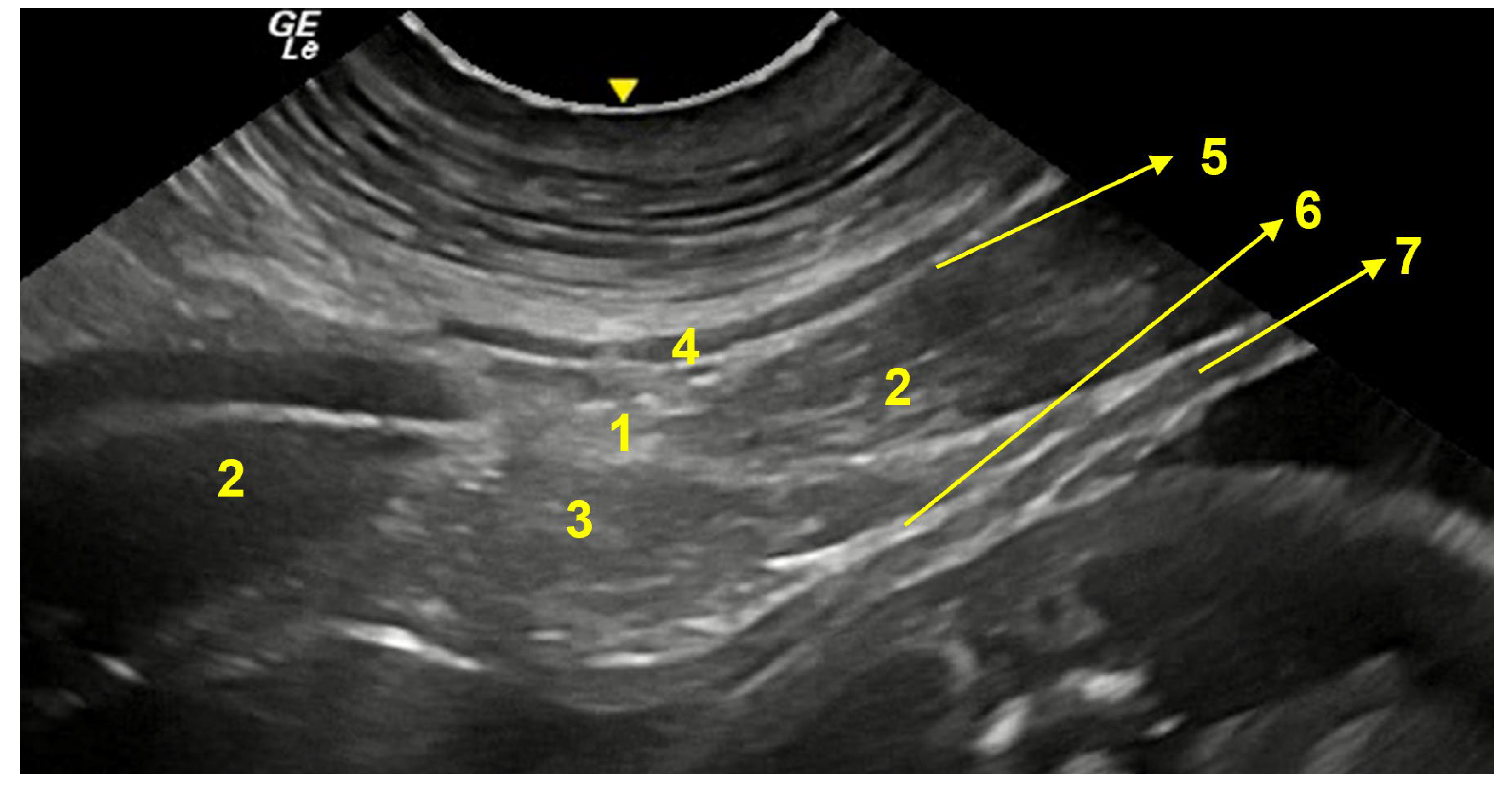
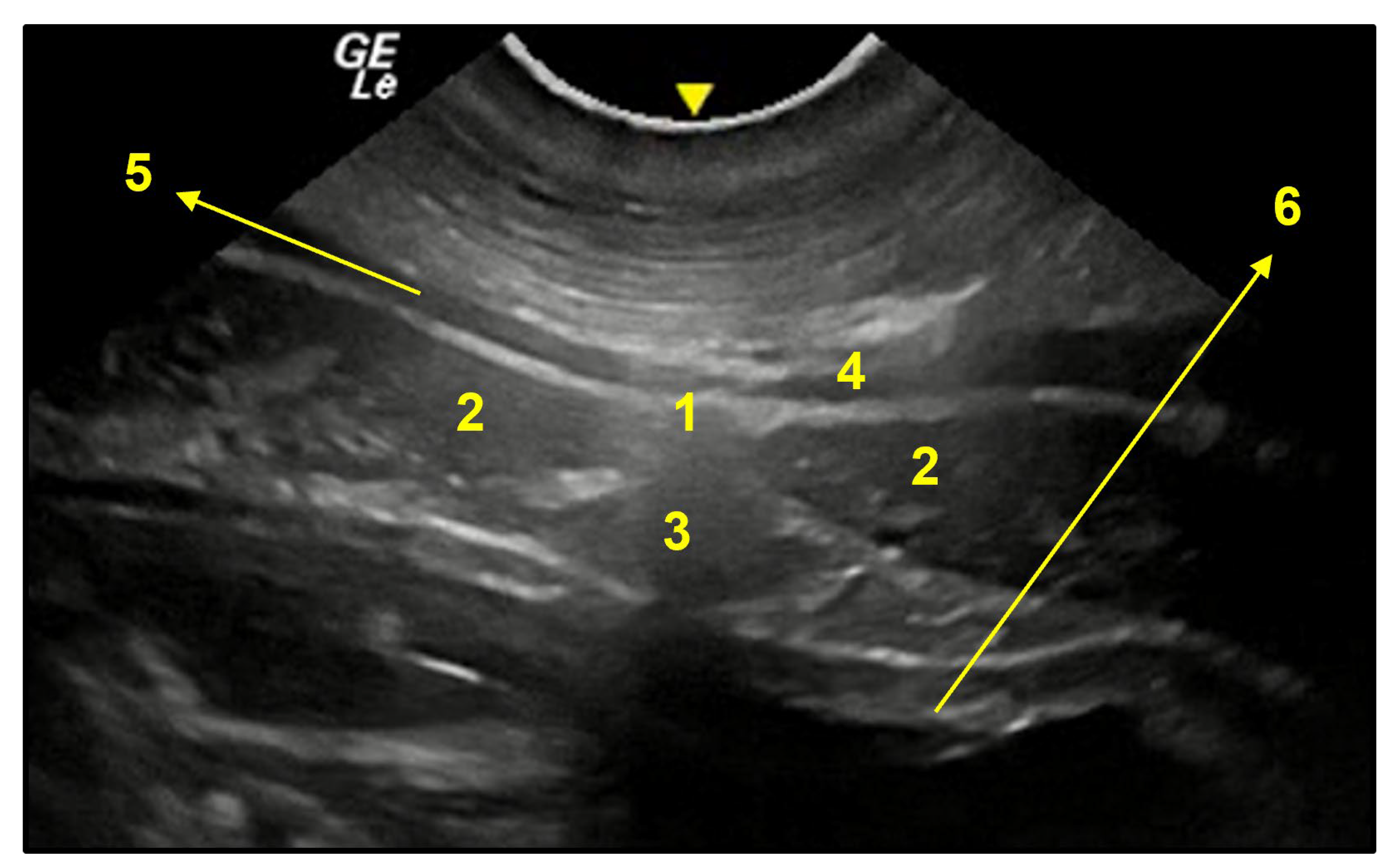
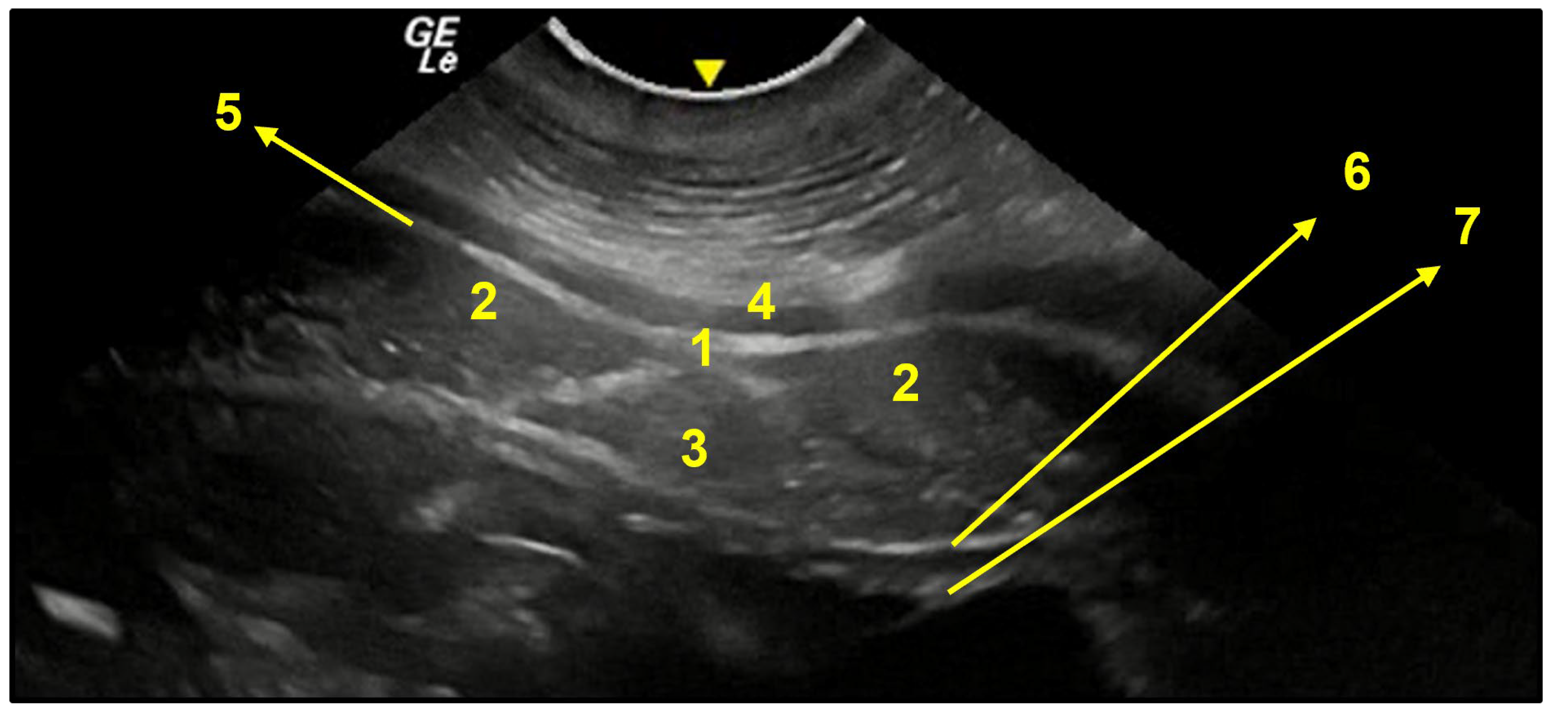
3.3. Ventral Abdominal Ultrasound
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ojasti, J.; Herrera, E. Estudio Biológico del Chiguire o Capibara; Fondo Nacional de Investigaciones Agropecuarias: Caracas, Venezuela, 1973; p. 273. [Google Scholar]
- Mones, A.; Ojasti, J. Mammalian Species: Hydrochoerus hydrochaeris. ASM 1986, 264, 1–7. [Google Scholar]
- Ferraz, K.M.P.M.D.B.; Bonach, K.; Verdade, L.M. Relationship between body mass and body length in capybaras (Hydrochoerus hydrochaeris). Biota Neotropica 2005, 5, 197–200. [Google Scholar] [CrossRef]
- Bovo, A.A.D.A.; Ferraz, K.M.P.M.B.; Verdade, L.M.; Moreira, J.R. 11. Capybaras (Hydrochoerus hydrochaeris) in Anthropogenic Environments: Challenges and Conflicts. In Biodiversity in Agricultural Landscapes of Southeastern Brazil; Gheler-Costa, C., Lyra-Jorge, M.C., Verdade, L.M., Eds.; De Gruyter Open Ltd: Warsaw, Poland; Berlin, Germany, 2016; pp. 178–189. [Google Scholar]
- São Paulo (Estado). Resolução Conjunta SMA/SUCEN n 01, de 24 de Março de 2016; Secretaria de Infraestrutura e Meio Ambiente: Sao Paulo, Brazil, 2016. [Google Scholar]
- Passos Nunes, F.B.; Jorge, F.M.G.; Nunes, M.P.; Nunes, A.Z.; Jorge-Neto, P.N.; Neto, A.C.A.; Labruna, M.B.; Pizzutto, C.S. Surgical sterilization of free-ranging capybaras (Hydrochoerus hydrochaeris): “Passos Nunes” uterine horn ligature. Anim. Reprod. 2022, 19, e20220029. [Google Scholar]
- Passos Nunes, F.B. Elimination of Brazilian Spotted Fever risk of transmission through capybaras (Hydrochoerus hydrochaeris) management in an area of transmission at Itu municipality, São Paulo. Master’s Thesis, Universidade de São Paulo, São Paulo, Brazil, 2019. [Google Scholar]
- Rosenfield, D.A.; Pizzutto, C.S. Wildlife population control—Reproductive physiology under the influence of contraceptive methods in mammalian wildlife, with emphasis on immunocontraception: The best choice? A literature review. Braz. J. Veter- Res. Anim. Sci. 2018, 55, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Hartman, M.J.; Monnet, E.; Kirberger, R.M.; Schoeman, J. Laparoscopic salpingectomy in two captive leopards (Panthera pardus) using a single portal access system. J. Zoo Wildl. Med. 2015, 46, 945–948. [Google Scholar] [CrossRef] [PubMed]
- Yanai, P.R.; Ferraro, M.A.; Lima, A.F.K.T.; Cortopassi, S.R.G.; Silva, L.C.L.C. Surgical contraception of free-ranging female capybaras: Description and comparison of open and minimally invasive techniques. Veter. Surg. 2022, 51, 11. [Google Scholar] [CrossRef] [PubMed]
- Ferraz, K.M.P.M.B. Desenvolvimento de um modelo de manejo de controle reprodutivo de capivaras no Campus “Luiz de Queiroz”. Projeto de Pesquisa- edital superintendência de gestão ambiental da USP. Universidade de São Paulo, Escola Superior de Agricultura “Luiz Queiroz”, Departamento de Ciências Florestais. Piracicaba, p. 10,2013. Available online: http://www.sga.usp.br/wp-content/uploads/sites/103/2017/06/43-Modelo-de-Manejo-de-controle-reprodutivo-de-capivaras-.pdf (accessed on 16 November 2020).
- Rodrigues, M.V.; Paula, T.A.R.; Ferreira, L.B.C.; Csermak Junior, A.C.; Araujo, G.R.; Deco-Souza, T. Manejo de população problema através de método contraceptivo cirúrgico em grupos de capivaras (Hydrochoerus hydrochaeris). Rev. Bras. Reprod. Anim. 2017, 41, 710–715. [Google Scholar]
- Boulanger, J.R.; Curtis, P.D.; Cooch, E.G.; Denicola, A.J. Sterilization as an alternative deer control technique: A review. Wildl. Interact. 2012, 6, 273–282. [Google Scholar]
- Pérez, W.; Senos, R.; Vazquez, N. Anatomy of the Gross Intestine of the Capybara (Hydrochoerus Hydrochaeris). Am. J. Anim. Veter- Sci. 2012, 7, 92–95. [Google Scholar] [CrossRef] [Green Version]
- Rosenfield, D.A.; Nichi, M.; Losano, J.D.; Kawai, G.; Leite, R.F.; Acosta, A.J.; Baquero, O.; Pizzutto, C.S. Field-testing a single-dose immunocontraceptive in free-ranging male capybara (Hydrochoerus hydrochaeris): Evaluation of effects on reproductive physiology, secondary sexual characteristics, and agonistic behavior. Anim. Reprod. Sci. 2019, 209, 106148. [Google Scholar] [CrossRef] [PubMed]
- Kiani, A.; Clauss, M.; Ortmann, S.; Vendl, C.; Congdon, E.R.; Herrera, E.A.; Kreuzer, M.; Schwarm, A. Digestive physiology of captive capybara (Hydrochoerus hydrochaeris). Zoo Biol. 2019, 38, 167–179. [Google Scholar] [CrossRef] [PubMed]
- Walker, F.O.; Cartwright, M.S.; Wiesler, E.R.; Caress, J. Ultrasound of nerve and muscle. Clin. Neurophysiol. 2004, 115, 495–507. [Google Scholar] [CrossRef] [PubMed]
- Tamayo-Arango, L.; Garzón-Alzate, A. Preservation of Animal Cadavers with a Formaldehyde-free Solution for Gross Anatomy. J. Morphol. Sci. 2018, 35, 136–141. [Google Scholar] [CrossRef] [Green Version]
- Pradere, J.D.; González, F.M.; Ruiz, A.Z.E.; Correa, A. Anatomía del Útero y Ovarios del Capibara (Hydrochoerus hydrochaeris): Irrigación Arterial. Rev. Fac. Cienc. Vet. 2006, 47, 25–32. [Google Scholar]
- Nomina Anatomica Veterinaria. International Committee on Veterinary Gross Anatomical Nomenclature, 6th ed.; Editorial Committee: Hannover, Germany; Ghent, Belgium; Columbia, MO, USA; Rio de Janeiro, Brazil, 2017; 178p. [Google Scholar]
- Hall, M.I.; Rodriguez-Sosa, J.R.; Plochocki, J.H. Reorganization of mammalian body wall patterning with cloacal septation. Sci. Rep. 2017, 7, 9182. [Google Scholar] [CrossRef] [PubMed]
- Dyce, K.M.; Sack, W.O.; Wensing, C.J.G. Textbook of Veterinary Anatomy, 4th ed; Elsevier: Amsterdam, The Netherlands, 2010; 849p. [Google Scholar]
- Konig, H.E.; Liebich, H.-G. Veterinary Anatomy of Domestic Animals—Textbook and Colour Atlas, 6th ed.; Thieme: Stuttgart, Germany, 2020; 859p. [Google Scholar]
- Potter, G.E.; Rabb, E.L.; Jones; Wm, D.C.; Hermann, C.L.; Gibbs, L.W. The Muscular System of Guinea Pig (Cavia porcellus). Bios 1957, 28, 104–115. [Google Scholar]
- Rizk, N.N. A new description of the anterior abdominal wall in man and mammals. J. Anat. 1980, 131, 373–385. [Google Scholar] [PubMed]
- Langworthy, O.R. A morphological study of the panniculus carnosus and its genetical relation to the pectoral musculature in rodents. Am. J. Anat. 1925, 35, 49–63. [Google Scholar] [CrossRef]
- Bahri, O.A.; Naldaiz-Gastesi, N.; Kennedy, D.C.; Wheatley, A.M.; Mccullagh, A.J.A.; Mccullagh, K.J.A. The panniculus carnosus muscle: A novel model of striated muscle regeneration that exhibits sex differences in the mdx mouse. Sci. Rep. 2019, 9, 15964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrera, H.A. Capybara Digestive Adaptations. In Capybara: Biology, Use and Conservation of an Exceptional Neotropical Species; Moreira, J.R., Ferraz, K.M.P.M.B., Herrera, E.A., Macdonald, D.W., Eds.; Springer: New York, NY, USA, 2013; pp. 97–106. [Google Scholar]
- Carvalho, R.G.d. Morfologia e biometria do aparelho reprodutor feminino da capivara (Hydrochoerus hydrochaeris). Ph.D. Thesis, Universidade Estadual Paulista (UNESP), Jaboticabal, São Paulo, Brazil, 2011. [Google Scholar]
- Miglino, M.A.; Santos, T.C.; Kanashiro, C.; Ferraz, R.H.d.S. Morphology and Reproductive Physiology of Female Capybaras. In Capybara: Biology, Use and Conservation of an Exceptional Neotropical Species; Moreira, J.R., Ferraz, K.M.P.M.B., Herrera, E.A., Macdonald, D.W., Eds.; Springer: New York, NY, USA, 2013; pp. 135–146. [Google Scholar]
- Orihuela, P.I.; Andrada, H.S.; Mac Entyre, C.Z. Irrigación Sanguínea de las Vísceras Abdominales del Carpincho. Ph.D. Thesis, Universidad de la República, Montevideo, Uruguay, 2019. [Google Scholar]
- Bester, E.G.; Schulman, M.; Kirberger, R.M.; Hartman, M. Unilateral salpingectomy in Sprague Dawley rats and its effect on litter size. J. South Afr. Veter- Assoc. 2021, 92, 2101. [Google Scholar] [CrossRef] [PubMed]
- Mcgrath, H.; Hardie, R.J.; Davis, E. Lateral flank approach for ovariohysterectomy in small animals. Compend. Contin. Educ. Small Anim. Pract. 2004, 26, 922–931. [Google Scholar]














Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jorge, F.M.G.; Donoso, F.M.P.M.; Alcobaça, M.M.d.O.; Cristofoli, M.; Passos Nunes, F.B.; Pizzutto, C.S.; Assis Neto, A.C.d. Surgical Anatomy for Sterilization Procedures in Female Capybaras. Animals 2023, 13, 438. https://doi.org/10.3390/ani13030438
Jorge FMG, Donoso FMPM, Alcobaça MMdO, Cristofoli M, Passos Nunes FB, Pizzutto CS, Assis Neto ACd. Surgical Anatomy for Sterilization Procedures in Female Capybaras. Animals. 2023; 13(3):438. https://doi.org/10.3390/ani13030438
Chicago/Turabian StyleJorge, Fabiana M. G., Flavia Maria Pia Montenegro Donoso, Mayla Magalhães de Oliveira Alcobaça, Marilu Cristofoli, Fernanda B. Passos Nunes, Cristiane S. Pizzutto, and Antonio Chaves de Assis Neto. 2023. "Surgical Anatomy for Sterilization Procedures in Female Capybaras" Animals 13, no. 3: 438. https://doi.org/10.3390/ani13030438