1. Introduction
Over the last three decades, landmark-based geometric morphometric (GM) methods have been increasingly applied to quantify and compare size and shape variation and covariation [
1,
2,
3,
4,
5]. Before performing GM analyses, the definition of a suitable configuration of landmarks in relation to the research aim is required [
6,
7]. A simple landmark configuration might be perfectly adequate to quantify shape differences appropriate to the question at hand. In studies of biological transformations such as growth or evolution, the landmarks define equivalent points that are ‘the same’ in terms of development or evolution (‘this point turns into that point’ is homologous), but the locations of homologous landmarks and their density are limited by the extent to which they can be identified and usually the presence of identifiable anatomical features, as well as the preservation of material and available time for digitization.
In many biological applications, landmarks cannot readily be identified, e.g., over smooth regions such as the human cranial vault or tooth crowns. In an attempt to provide detailed information on such regions, different approaches have been proposed to mark up semilandmarks (or dense point correspondences) among curves or surfaces between landmarks [
8,
9]. The method of sliding semilandmarks, which locates semilandmarks by minimising the bending energy of thin-plate splines (TPS) or Procrustes distance [
10,
11,
12], is most commonly used in biology. Alternative semilandmarking methods include rigid registration approaches, e.g., the auto3dgm package [
13] based on the iterative closest points (ICP) algorithm [
14], and non-rigid registration approaches, e.g., non-rigid ICP (NICP) [
15,
16] and the optical flow algorithm [
17], among others. The fundamental task of these semilandmarking approaches is to transfer the semilandmarks from a template surface (e.g., a mean surface) to the target specimen. It is worth noting that semilandmarks rely primarily on mathematical mappings and/or the similarity of topographic features, rather than developmental or evolutionary equivalences based on prior knowledge.
Recent studies have assessed the performance of different semilandmarking approaches based on principal components (PCs) [
10,
13,
18,
19,
20], distance matrices [
13,
21,
22], and the geometric deviation between template and transformed meshes [
18,
23]. These have found that different approaches yield different semilandmark locations and, thus, result in analytical results that differ to some degree. This was further investigated in a prior study [
24] that provided the starting point for the present one. The performance of three of the semilandmarking approaches described above was systematically examined. These included the sliding TPS approach outlined above. The second approach employed hybrid rigid registration combining least-squares (LS) [
25] and ICP algorithms (LS&ICP). After using the LS algorithm to fit the template landmarks to those of each specimen, the ICP algorithm rigidly refitted the template to the target, minimising the sum of squared distances between landmarks and estimated semilandmarks, found by searching for the nearest points on the target from the registered template semilandmarks. The third approach (TPS&NICP) [
26] used TPS to perform an initial non-rigid registration of the template landmarks and surface to specimens, and then the NICP algorithm [
15] was applied to further warp the deformed template surface to each specimen as rigidly as possible, optimizing the cost function by assigning an affine transformation to each vertex, rather than an interpolation function as used in TPS, before transferring the semilandmarks from the template to the nearest point of the specimen surfaces. We compared semilandmarking approaches, differences in the locations of semilandmarks, Procrustes distances between landmark and semilandmark configurations, estimates of mean landmark and semilandmark configurations, PCs of configuration shape, and estimates of allometry.
Because homology is unknown for regions that were semilandmarked, it is not possible to assess how well semilandmarks represent homology; rather, the focus was on comparing the results of analyses based on semilandmarks between and within methods, with increasing semilandmark density. The analyses showed that each semilandmarking approach yields different semilandmarks locations, which result in differences in each of the comparisons [
24]. The sliding TPS algorithm and TPS&NICP approach yielded results that are more similar to each other than those based on LS&ICP. Further, we assessed consistency within methods among results obtained using different densities of semilandmarks, finding that sliding TPS and TPS&NICP approaches are most consistent, especially where true landmarks are dense. The extent to which these differences are important depends on the context, the question being addressed, and the purpose of the study, but all semilandmarking approaches estimate homology with error, the extent of which is unknowable. Therefore, all subsequent statistical analyses that aim to describe developmental or evolutionary transformation are subject to that error and should be treated with an appropriate degree of caution [
7,
24].
Geometric morphometric analyses enable the visualisation of statistical findings, generating landmark and semilandmark configurations that represent shapes or forms (sizes and shapes) of interest such as the mean or allometrically scaled configurations. Surfaces or regular grids are often warped to these configurations to aid the visualisation of shape differences and, where applicable, changes. This is most commonly performed using TPS [
27,
28]. However, the authors of [
6] noted that ‘With sliding semilandmarks, their relative positions on equivalent curves, surfaces, etc. are not singly interpretable, but rather should be read as a whole, respecting the fact that the underlying assumption in their construction is one of equivalence of the curve or surface patch as a whole’. This was recently reiterated [
3,
29]: ‘the coordinates of semilandmarks along the surface are meaningless, and one cannot interpret the position of single semilandmarks, only the surface geometry that all semilandmarks describe together’. Thus, although semilandmarks are treated as landmarks in statistical analyses, ‘errors’ in their locations (or differences using different methods to locate them) influence statistical outcomes, as was demonstrated in the previous study [
24]; visualisations and interpretations of differences should ignore their locations and focus on the shape of the curve or surface they describe.
It is, therefore, of interest to know the extent to which the shapes of surfaces warped to fit semilandmark and landmark configurations varying in semilandmark density and locations (e.g., arising from different approaches to placing them) are consistent. If different densities and approaches yield identical or very similar visualisations, this may be reassuring in certain practical applications. For instance, a mean surface might be used in clinical work to compare measurements taken on a patient with an estimate of the population mean [
30] and facial approximation from the skull alone in the realm of forensic science [
31]. Additionally, surfaces from GM analyses are used to virtually repair and reconstruct fossil material [
32] and build 3D models for functional analyses such as finite element analysis (FEA) [
33]. The extent to which such estimated surfaces differ when derived using different semilandmark densities and semilandmarking approaches is unknown, yet it is important in that it may affect subsequent morphometric or functional analyses. This question is addressed in the present study.
The main purpose of this study is to empirically test two hypotheses using surface scans of human heads and ape crania: that there are no differences in surface mesh shape (the shape of the configuration of surface vertices and the nodes of the surface mesh, rather than the landmark and semilandmark configuration) between estimates derived using different semilandmarking densities and approaches applied to surfaces representing (a) the mean of a sample and (b) allometrically scaled shapes.
To these ends, a template surface mesh is warped to fit the estimated mean and allometrically scaled mean landmark and semilandmark configurations derived from different semilandmarking densities and approaches, and the resulting surfaces are compared. Additionally, these surfaces are compared with surfaces warped using landmarks alone. Of interest is the extent to which these surfaces differ and how they differ. The focus is on the comparison of the shape of the surface rather than the geometry of the underlying mesh.
4. Discussion
The use of digital surface meshes of biological and anthropological specimens in 3D GM studies has become increasingly common, as has the use of landmarks and semilandmarks generated by different semilandmarking approaches in order to compare the details of morphology [
10,
13,
18,
21,
34,
40]. While dense coverage by semilandmarks allows for more detailed descriptions of form and, potentially, biological signals [
41], it introduces several difficulties in comparing forms. Further, given that semilandmarks are treated as equivalent between specimens in GM analyses and are given the same weight as landmarks, the basis of equivalence is an important consideration. In studies of biological transformations such as those that occur during development and evolution, the equivalences required to model and compare them are developmental or evolutionary. Landmarks and semilandmarks at each stage need mark-up points that are equivalent between specimens in terms of development or evolution at another stage (homologous points). For landmarks, this matching is based on prior knowledge, but for semilandmarks, it is algorithmic and relies on mathematical mappings and topographical features. As such, the extent to which semilandmarks can be considered homologous has contributed to the debate about their validity and usefulness in relation to the study of developmental or evolutionary transformations [
6,
7,
41].
It has been noted by previous researchers that because the locations of semilandmarks on surfaces and curves are uncertain they should not be interpreted singly, but rather as a whole [
3,
6,
29]. While this avoids overinterpreting differences in individual semilandmark locations, it does not avoid statistical issues. Thus, differences in semilandmark locations will lead to different distance matrices among specimens and, thus, to different analytical results. The extent of this issue has been explored in several previous studies [
11,
13,
18,
19,
20,
21,
22]. Additionally, the use of high-density semilandmarks raises statistical issues related to the ratio of variables to specimens (i.e., high
p and low
n) and in assessing covariances within landmark and semilandmark configurations [
7,
42].
Statistical considerations aside, high-density semilandmarks are routinely used to assess shape variations and covariations and to perform classification [
41,
43,
44,
45], with results presented as visualisations of a warped surface mesh. It is, therefore, of interest to know how different semilandmarking approaches and semilandmark densities affect visualisations. This has been addressed by the analyses presented here.
In this study, we compare surface meshes warped to configurations of landmarks and semilandmarks arising from GM analyses that represent the overall mean and allometrically scaled surfaces. The aim is to compare the surface meshes used for visualisation rather than the statistical outcomes of analyses of the landmark and semilandmark configurations. These were compared in [
24]. Three different semilandmarking approaches were used with varying semilandmark densities. These are the method of sliding semilandmarks, minimising the bending energy of a set of thin-plate splines or Procrustes distances [
37], the non-rigid combined approach of TPS&NICP [
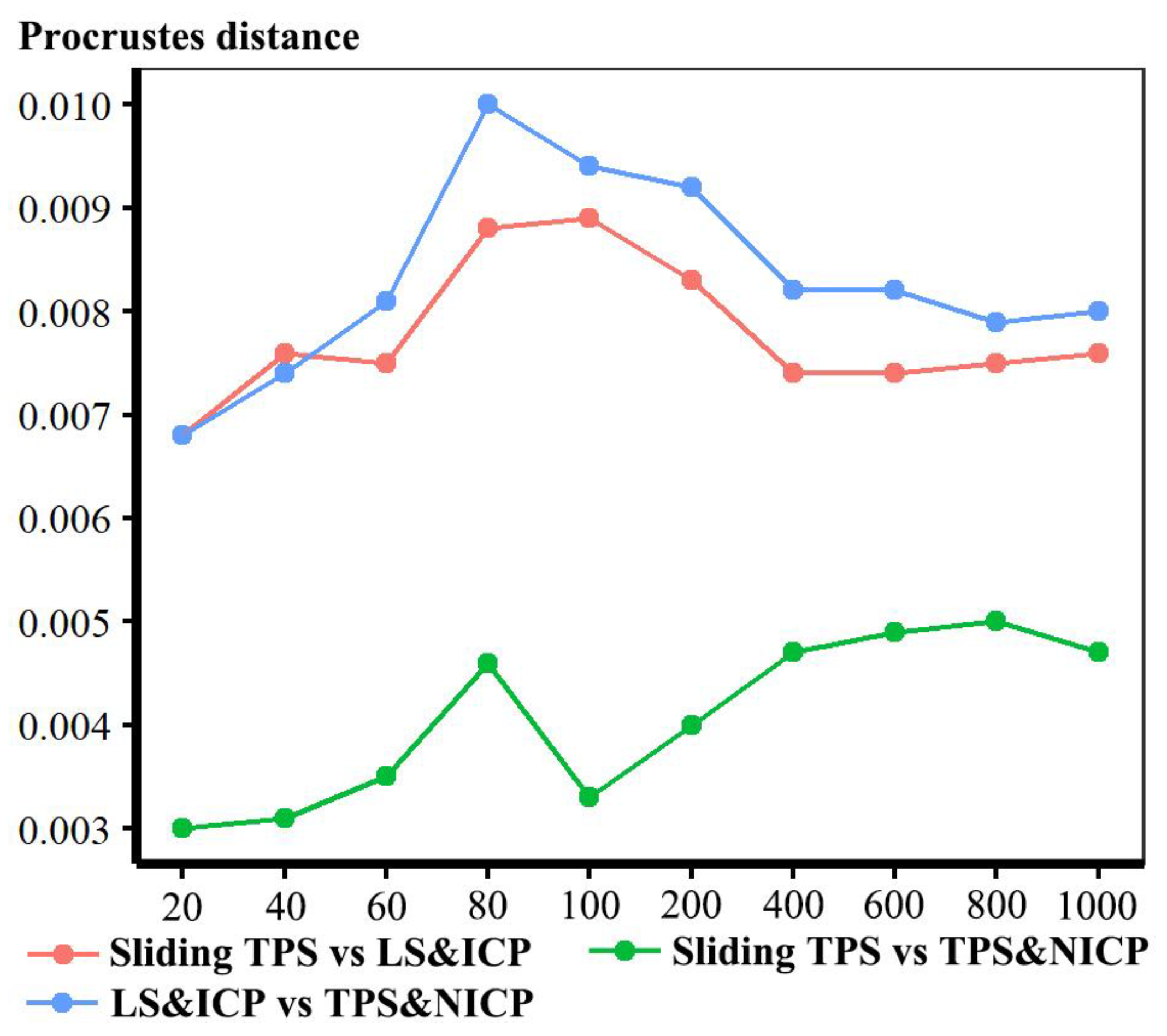
26], and the rigid LS&ICP approach. These lead to semilandmark configurations that differ in the locations of semilandmarks. These differences are smallest between sliding the TPS and TPS&NICP approaches and larger when comparing these with the LS&ICP approach (
Figure 2). However, the locations of individual semilandmarks are not interpretable and, as noted above, they lie on the surface and so should be interpreted as a whole in terms of the differences between surfaces that fit them.
This study aimed to achieve this by empirically testing two hypotheses using surface scans of human heads and ape crania: that there are no differences in surface mesh shape between estimates derived using different semilandmarking densities and approaches applied to surfaces representing (a) the mean of a sample and (b) allometrically scaled shapes. The surfaces were quantitatively compared using the coordinates of their vertices after re-semilandmarking and re-warping (see Methods) to calculate Procrustes distances between them and, where relevant, by extracting and comparing principal components. They were visually compared using colour maps of differences in local surface areas. Both hypotheses are falsified; differences clearly exist between estimated mean and allometrically scaled surfaces, but the degree of difference between semilandmarking approaches is small to moderate, with the non-rigid semilandmarking approaches (sliding TPS and TPS&NICP) showing a high degree of consistency.
Because landmarks have more secure homology than semilandmarks and should be chosen with respect to the question at hand [
6,
7], they are likely few in number and less likely to result in statistical issues arising from large numbers of variables relative to the number of specimens. Additionally, surfaces can be warped to landmarks to visualise analytical results, albeit with less detail than warping based on dense correspondences. Thus, the present study also assessed differences between warped surfaces based on landmarks and semilandmarks and those based on the landmark configuration alone, using different reference surfaces.
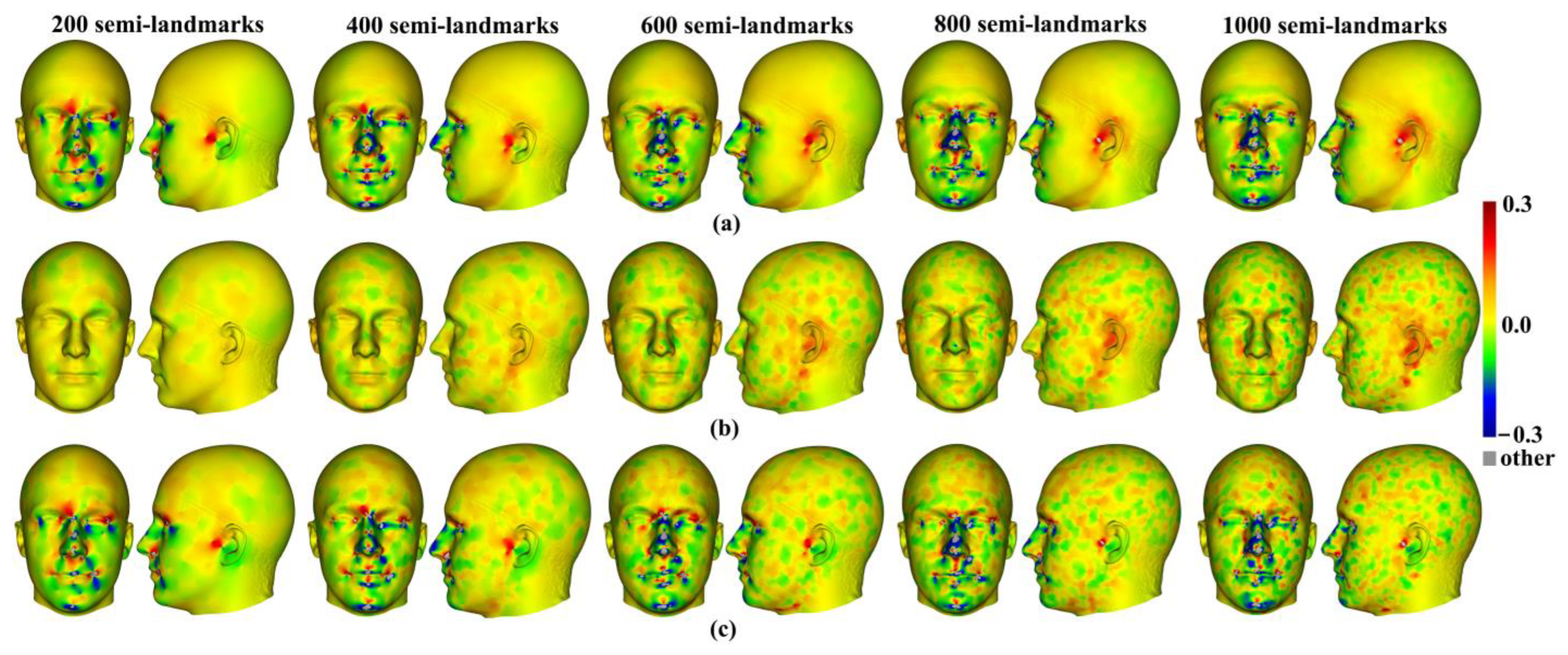
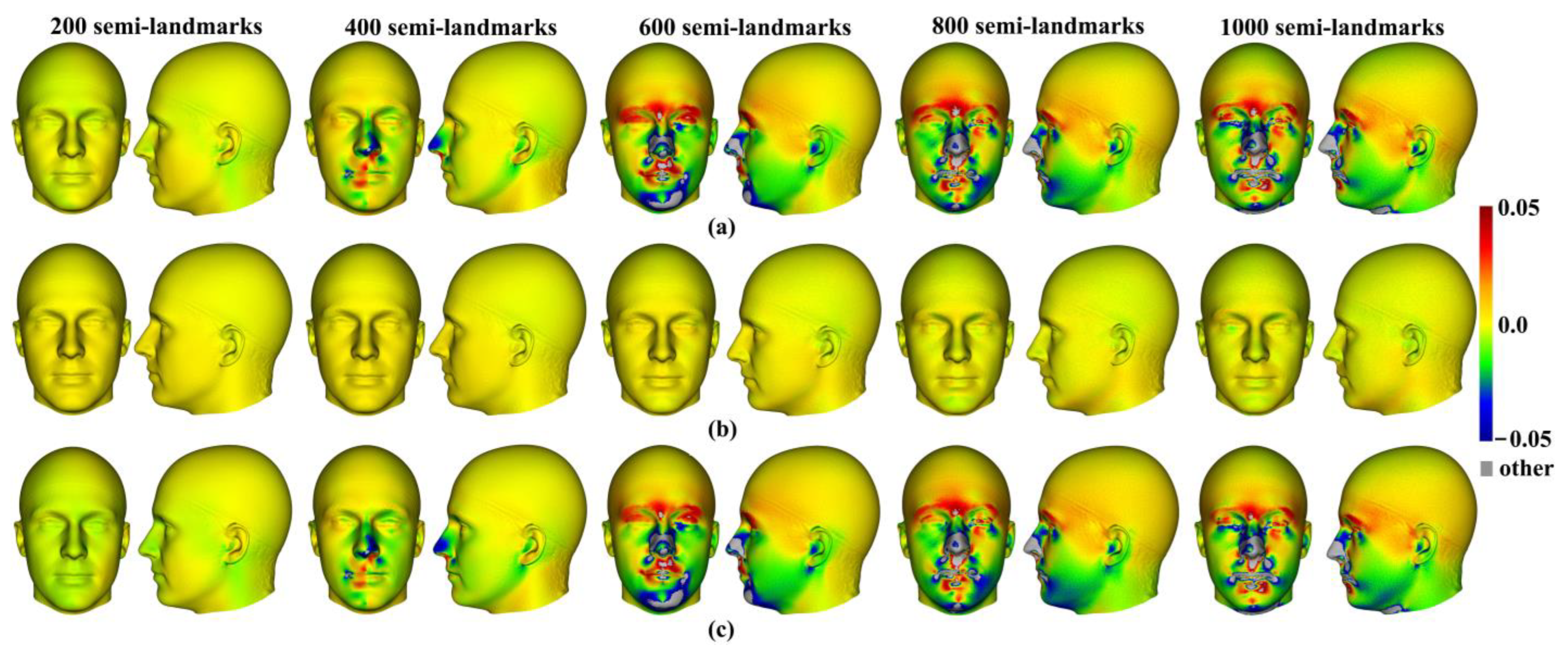
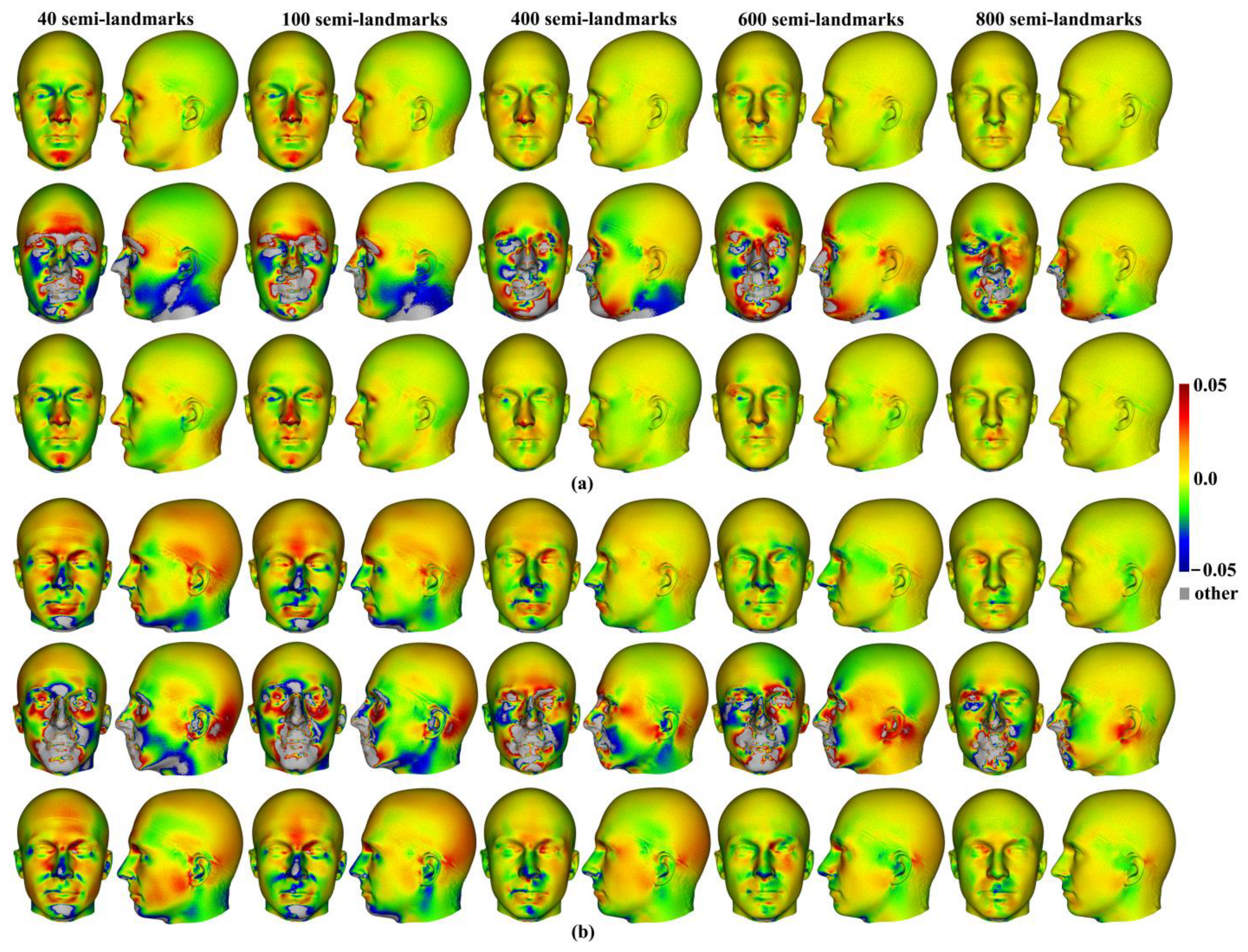
Three semilandmarking approaches were used to estimate the sample mean surface meshes by warping the template (an initial estimate of the average surface in each dataset) to the mean landmark and semilandmark coordinates arising from each method using varying semilandmark densities. For the head surfaces, the means are visually quite similar (
Figure 4) but differ in detail (
Figure 6 and
Figure 7). The resulting mean surfaces from sliding TPS and TPS&NICP are the most similar, and those from LS&ICP are the most different. Similar results are obtained in estimating the mean surface of the ape crania (
Table 1 and
Figure 8 and
Figure 9), but the LS&ICP approach performed poorly in locating semilandmarks in reasonably corresponding locations with the more complex ape cranial surfaces. In both datasets, estimated mean surfaces converge with increasing semilandmarking density on the surface from the highest density (
Table 4 and
Table 5 and
Figure 10,
Figure 11 and
Figure 12). For the head surface data, warping the template surface to the mean landmark configuration (
Figure 23b) resulted in a surface that was quite similar in general to that warped to landmarks and high-density semilandmarks, but differed in detail from the semilandmark-based mean (
Figure 23d). This similarity is in large part due to the fact that the template surface is already an initial estimate of the mean. Repeating the analysis using the surface of the individual nearest to the mean landmarks and semilandmarks resulted in an estimate of the mean surface (
Figure 23c) that presented greater differences from the semilandmark-based mean surface (
Figure 23e). Visually, this approach worked reasonably despite the lack of identifiable landmarks to guide the warping of the scalp; however, this is likely because the template scalp was not an initial estimate of, and very similar to, the mean.
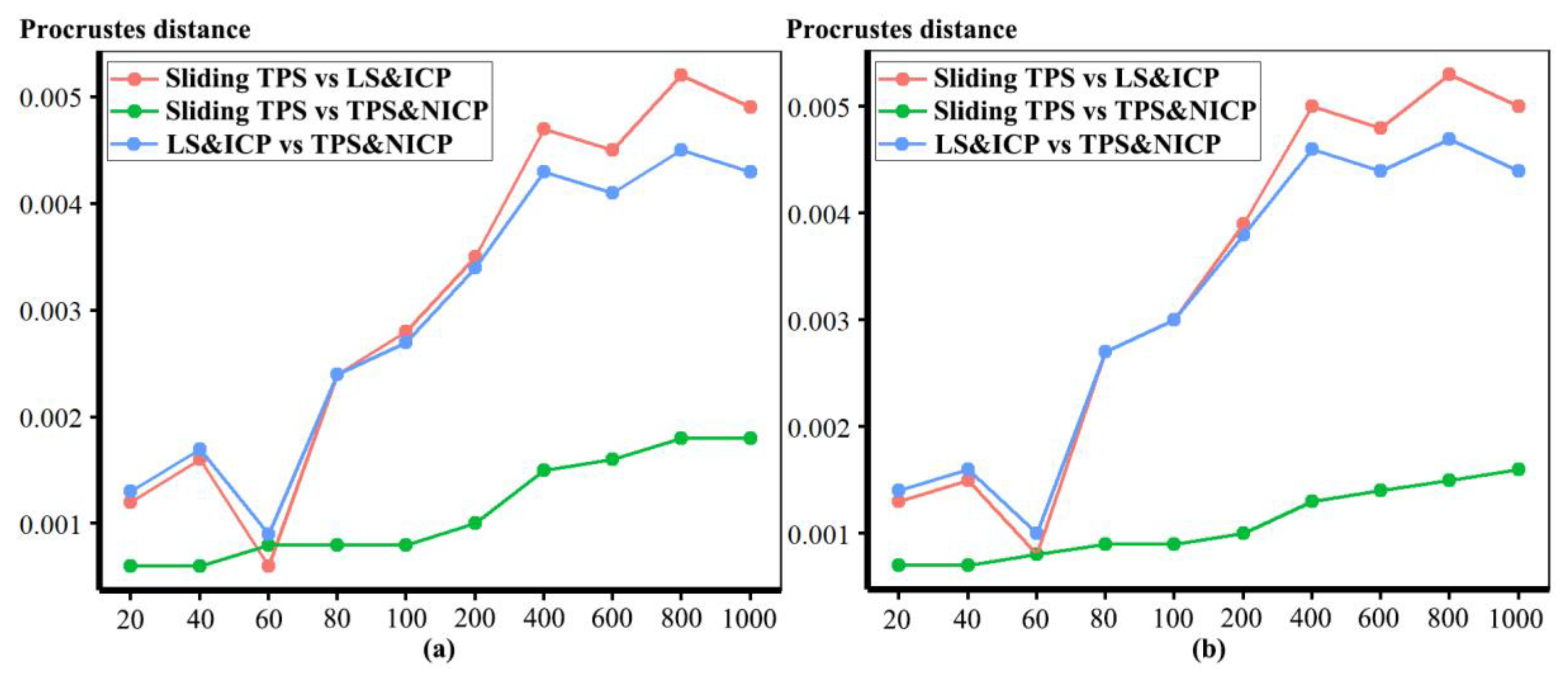
The mean ape surfaces estimated using sliding TPS and TPS&NICP with varying densities of semilandmarks are also visually quite similar (
Figure 8), although the surface from LS&ICP shows some obvious differences. Focusing on sliding TPS and TPS&NICP, the mean surfaces resulting from these methods using varying numbers of semilandmarks are very similar, with differences increasing with semilandmarking density, especially where surface topography is complex (
Figure 9 and
Table 1). Surfaces estimated using increasing numbers of semilandmarks converge on the surface estimated using the maximum number of semilandmarks (
Figure 10,
Figure 11 and
Figure 13).
It should be noted that in the implementation of NICP used here, the initial registration of surfaces between the template and target uses a triplet of TPS. This is also the case for the sliding TPS approach. This shared initial, non-rigid registration doubtless contributes to the similarities in results obtained using these approaches when compared to the rigid, least-squares registration employed in the LS&ICP approach. However, even the LS&ICP approach used the same landmark set for registration. It would be of interest in future work to assess the impact of using different landmark configurations to estimate semilandmarks.
Using the mean landmark configuration alone to warp the template surface mesh results in a visually similar surface to the mean based on landmarks and high-density semilandmarks, but it differs in detail, especially around crests and ridges (
Figure 24a,b,d). Visualising the mean by warping the ape surface used to generate the template results in a more different surface (
Figure 24c,e,f), which, in some ways resembles the mean based on landmarks and high-density semilandmarks (
Figure 24a), but it differs particularly in regions with complex topography (
Figure 24c,f). These landmark-based warpings differ in detail from the landmark and semilandmark-based ones, but they also bear a resemblance. Whether or not they are adequate depends on the purpose to which they are put. They may be sufficient to describe general aspects of shape variation but would likely yield different results if used to build finite element models (FEM). The warping of a surface that is an initial estimate of the mean to the landmarks alone inevitably yields a surface more like that based on landmarks and semilandmarks than warping a surface from an individual, even if close to the mean. This also applies to comparisons of mean surfaces resulting from semilandmarking approaches and densities.
The predicted allometrically scaled mean surfaces were also compared among semilandmarking approaches and densities. With the head surface dataset, sliding TPS and TPS&NICP produced very similar surfaces, particularly at the highest semilandmarking densities (
Figure 14 and
Figure 15). The surfaces from LS&ICP were dissimilar. Likewise, for ape cranial surfaces, the allometrically scaled mean surfaces from sliding TPS and TPS&NICP are similar but differ in detail, especially around ridges and crests (
Table 6 and
Figure 16). They become more dissimilar in the regions of crests and ridges as semilandmarking density increases, reflecting the more detailed control of warping from greater semilandmark densities. Both semilandmarking approaches show a similar pattern of convergence on the surface derived from the highest density, of surfaces with increasing densities of semilandmarking (
Figure 21 and
Figure 22, and
Table 8).
These differences among allometrically scaled means from both datasets and the different approaches and densities of semilandmarking are summarized by the PC plots in
Figure 17 and
Figure 18.
Figure 17 presents for the head surface data, the first two PCs from an analysis of the mean, and allometrically scaled mean surfaces derived from varying densities of semilandmarks and each approach. It shows that sliding TPS and TPS&NICP achieve very similar results, with many points overlapping, but LS&ICP results in quite different estimates of the same surfaces, which vary along a different vector from the other two approaches. The comparable analysis for the ape crania compared only sliding TPS and TPS&NICP, and the resulting PC plot shows that these achieve very similar results. These findings provide a perspective on the differences identified in the Procrustes distance matrices and visual comparisons in the analyses described above. Thus, the Procrustes distances between the mean surfaces from varying the semilandmarking approaches and densities are small compared with those between surfaces allometrically scaled to the maximum and minimum sample centroid sizes. The colour maps are very sensitive, identifying and emphasising what are, in reality, very small differences.
Allometrically scaled ape cranial surfaces from sliding TPS with 800 semilandmarks are compared with surfaces derived by warping the template surface and the surface used to generate the template to the allometrically scaled landmark configurations. The resulting predictions of surfaces at both the sample maximum and minimum centroid sizes share general similarities with, but differ in detail from, the surfaces based on semilandmarks (
Figure 25d,e,f and
Figure 26d,e,f). As with the head surfaces, these differences reflect similar aspects of scaling, which may be adequate in describing general scaling trends but would likely lead to differences in FEA results among models based upon them.
The differences in scaling are emphasised by the PCAs of
Figure 27 and
Figure 28, where, for both datasets, the surfaces derived by warping the surface of the individual nearest to the mean to the allometrically scaled mean landmark configurations are distant from the semilandmark-based surfaces and are arranged along a vector that is not parallel to the vector between surfaces scaled using semilandmarks. Warping the template surface to the mean and allometrically scaled means in both datasets results in a vector parallel to that derived by warping the head surface of the individual nearest to the mean or the ape cranium used to generate the template, but with the mean close to the means from the semilandmark-based approaches. This indicates that these different surfaces scale in very similar ways. Thus, the choice of template surface determines where in the shape space the allometric vector is located while the landmarks and semilandmarks used to deform the surface determine how it is deformed. Semilandmarks result in the surface regions between landmarks being deformed differently from what is achieved by warping to the landmark configurations alone. This is not surprising and underlines how semilandmarks contribute to controlling surface deformations.
The results of this study show that different semilandmarking approaches and densities achieve different visualisations of mean and allometrically scaled surfaces. The degree of difference depends on the approach, with non-rigid semilandmarking (sliding TPS and TPS&NICP) producing surfaces that are consistently more similar to each other than to those derived using the rigid LS&ICP approach. Additionally, the non-rigid approaches show consistency in the surfaces produced using semilandmarks of varying densities. While Procrustes distances and colour maps emphasise differences among the approaches, PCAs comparing the scaled mean surfaces show that the differences between surfaces from non-rigid semilandmarking approaches are very small when compared to the differences among allometrically scaled means. The differences between surfaces derived using LS&ICP are greater.
Semilandmarking involves a great deal of extra effort compared with landmarking alone, and, as has been noted earlier, brings with it some severe statistical issues. This has led to the questioning of their benefits and criticism stating that they may lead to erroneous conclusions [
7,
42]. Thus, this study compared surfaces warped using landmark configurations alone with those from landmark and semilandmarking configurations. These comparisons have shown that if a surface that is an initial estimate of the mean surface is used then the mean surfaces are well estimated. This is to be expected since the mean landmarks have little warping to do. This finding likely explains why LS&ICP results in more similar mean surfaces to those from sliding TPS and TPS&NICP at lower rather than higher semilandmarking densities (
Figure 6 and
Figure 7). When an alternative surface is used, the surface visualisation is different, having inherited features from this new surface. Surfaces warped to scaled landmark configurations show differences and some similarities to those warped to landmarks and semilandmarks in combination. Such analyses and visualisations based on landmarks alone may be perfectly adequate for many questions; they involve less work to produce and avoid the statistical issues that can arise with many semilandmarks and few specimens. However, compared with surfaces from semilandmarks, they would likely lead to different results if used to build finite element models.
Finally, we should emphasise that consistency is not the same as accuracy [
7]. It is tempting to conclude that the remarkable consistency of surface shapes derived using sliding TPS and TPS&NICP reflects accuracy in the estimation of means. Our results cannot, however, support or refute this possibility since no ‘true mean’ is known (or knowable). Estimates of means depend on what quantities are measured and compared because means are a statistical, rather than biological, entity, particular to the data used to calculate the mean. The results are ‘correct’ for the variables (semilandmark locations) resulting from each method. However, with semilandmarks, there is inevitable uncertainty about the extent to which they are equivalent between specimens in terms of homology. Our studies show that differences in semilandmark locations among specimens will lead to differences in statistical results [
24] and visualisations (present study). In these studies, these differences are quite small relative to the differences among specimens, but it is not clear to what extent these empirical results apply to diverse datasets and semilandmarking approaches (e.g., minimisation of Procrustes distances by sliding [
10]; morphometric ‘fishnets’ [
46]). This can only be addressed by further extensive studies of real data and through simulation experiments, in which an initial ‘mean’ is perturbed and then estimated from the perturbed data.
For now, we have shown that the two non-rigid semilandmarking approaches yield consistent estimates of mean and scaled surfaces. Semilandmarking involves a great deal of additional work and runs statistical risks in analyses. With these things in mind, the investigator should carefully consider if semilandmarking is necessary to answer the question at hand and balance this need against the statistical and biological (e.g., regarding homology) downsides and the time involved in gathering and using semilandmarks to assess shape variances and covariances. It may be a more secure strategy to base statistical tests on homologous landmarks and visualisations on landmarks and semilandmarks from parallel analyses.
It should be borne in mind that homology is often also uncertain for landmarks and that different sets of landmarks will lead to different results. However, the three approaches that we compared in this study led to visually similar estimates of surface meshes that may be adequate for visualisation and functional simulation, in the sense that they are likely to be fair representations of average and scaled surfaces, but there is no single ‘true’ representation against which to assess this (see above). Their applicability depends on how much error in the estimation of the surface shapes is judged acceptable given the context of the particular study.
Finally, it should be noted that this study is limited in its scope; being based on only human heads and ape crania, different datasets need to be examined to assess the reliability of the findings. Studies also need to be conducted using simulated data in which true mean and allometrically scaled surfaces are known in order to assess the accuracy of the estimates of these surfaces. Additionally, this study compared a limited range of possible approaches to semilandmarking, and future work needs to extend these comparisons to include other methods and ‘landmark-free’ approaches.
































{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}