

Effect of Inoculating Two Bacteriocin-Producing Lactiplantibacillus plantarum Strains at Ensiling on In Vitro Rumen Fermentation and Methane Emissions of Alfalfa Silage with Two Dry Matter Contents

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Strain Preparation
2.2. Alfalfa Silage Preparation
2.3. Lactic Acid Fermentation Characteristics and Fiber Analysis
2.4. In Vitro Rumen Fermentation
2.5. Statistical Analysis
3. Results
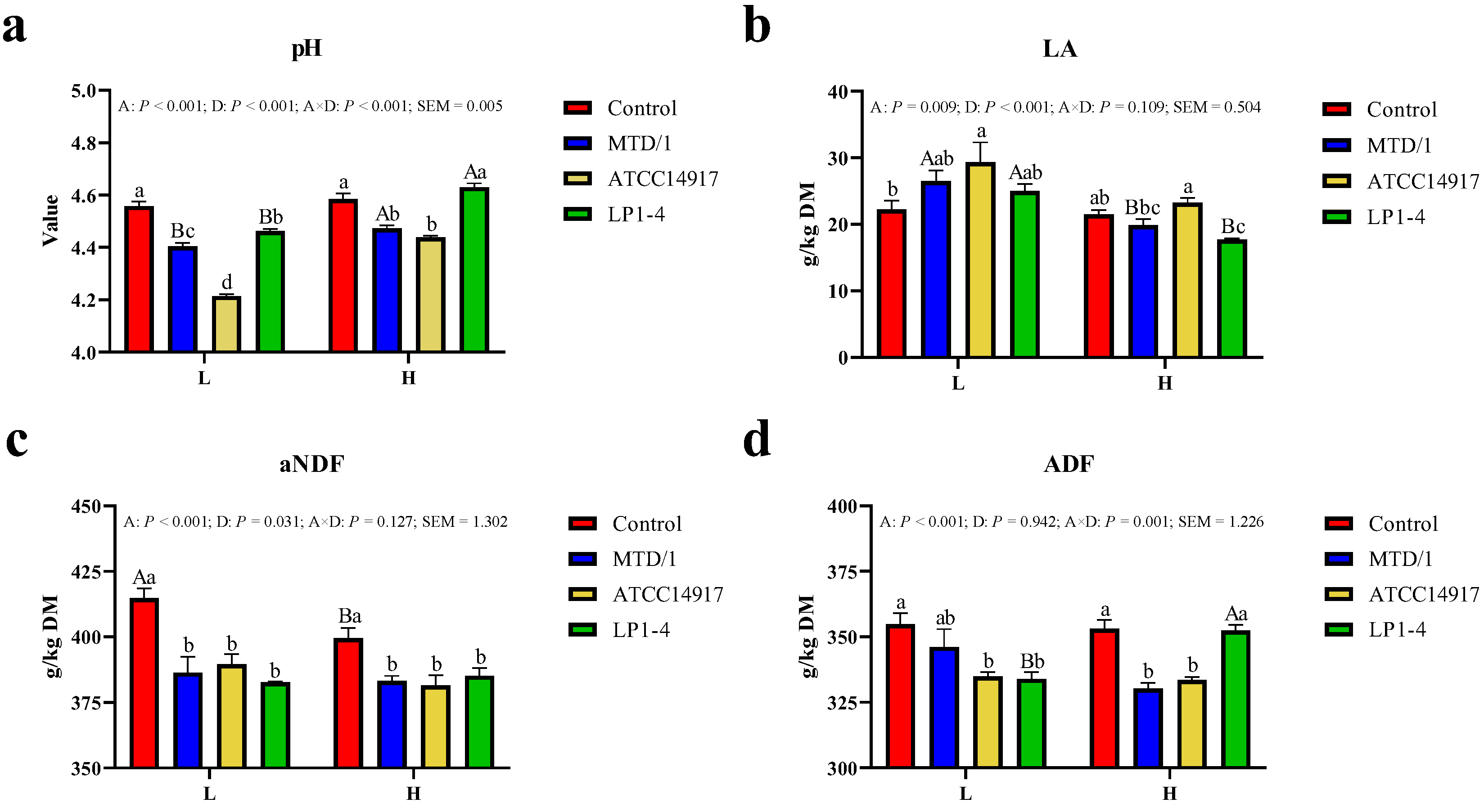
3.1. Lactic Acid Fermentation Characteristics and Fiber Contents of Silages at Moderate and High DM Contents after 90 Days
3.2. CH4 Production and Fermentation Characteristics of In Vitro Ruminal Fermentation of Alfalfa Silage
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhou, Y.; Mao, H.; Jiang, F.; Wang, J.; Liu, J.; Mcsweeney, C.S. Inhibition of rumen methanogenesis by tea saponins with reference to fermentation pattern and microbial communities in Hu sheep. Anim. Feed Sci. Technol. 2011, 166–167, 93–100. [Google Scholar] [CrossRef]
- Scheehle, E.A.; Kruger, D. Global anthropogenic methane and nitrous oxide emissions. Energy J. 2006, 27, 33–44. [Google Scholar] [CrossRef]
- McMichael, A.J.; Powles, J.W.; Butler, C.D.; Uauy, R. Energy and health 5-Food, livestock production, energy, climate change, and health. Lancet 2007, 370, 1253–1263. [Google Scholar] [CrossRef]
- Murray, R.M.; Bryant, A.M.; Leng, R.A. Rates of production of methane in the rumen and large intestine of sheep. Br. J. Nutr. 1976, 36, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lan, W.; Yang, C. Ruminal methane production: Associated microorganisms and the potential of applying hydrogen-utilizing bacteria for mitigation. Sci. Total Environ. 2019, 654, 1270–1283. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Liu, Z.; Yu, Z.; Zhu, W. Monensin and nisin affect rumen fermentation and microbiota differently in vitro. Front. Microbiol. 2017, 8, 1111. [Google Scholar] [CrossRef] [Green Version]
- Michael, C.A.; Dominey-Howes, D.; Labbate, M. The Antimicrobial resistance crisis: Causes, consequences, and management. Front. Public Health 2014, 2, 145. [Google Scholar] [CrossRef] [PubMed]
- Gillor, O.; Nigro, L.M.; Riley, M.A. Genetically engineered bacteriocins and their potential as the next generation of antimicrobials. Curr. Pharm. Design 2005, 11, 1067–1075. [Google Scholar] [CrossRef] [Green Version]
- Santos, J.C.; Sousa, R.C.; Otoni, C.G.; Moraes, A.R.; Souza, V.G.; Medeiros, E.A.; Espitia, P.J.; Pires, A.C.; Coimbra, J.S.; Soares, N.F. Nisin and other antimicrobial peptides: Production, mechanisms of action, and application in active food packaging. Innov. Food Sci. Emerg. 2018, 48, 179–194. [Google Scholar] [CrossRef]
- Cleveland, J.; Montville, T.J.; Nes, I.F.; Chikindas, M.L. Bacteriocins: Safe, natural antimicrobials for food preservation. Int. J. Food Microbiol. 2001, 71, 1–20. [Google Scholar] [CrossRef]
- Direkvandi, E.; Mohammadabadi, T.; Dashtizadeh, M.; Alqaisi, O.; Salem, A.Z.M. Lactobacillus plantarum as feed additive to improvement in vitro ruminal biofermentation and digestibility of some tropical tree leaves. J. Appl. Microbiol. 2021, 131, 2739–2747. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Zhang, L.; Yi, H.; Shi, J.; Xue, C.; Li, H.; Jiao, Y.; Shigwedha, N.; Du, M.; Han, X. Qualitative detection of class IIa bacteriocinogenic lactic acid bacteria from traditional chinese fermented food using a ygngv-motif-based assay. J. Microbiol. Meth. 2014, 100, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Ma, G.; Ma, H.; Lu, X.; Liu, J.; Sun, Y.; Bai, F.; Li, J. Screening for broad-spectrum antagonistic lactic acid bacteria from intestine of turbot and identification of bacteriocin produced by it. Food Sci. 2019, 6, 159–165. [Google Scholar] [CrossRef]
- Yang, F.; Wang, Y.; Zhao, S.; Wang, Y. Lactobacillus plantarum inoculants delay spoilage of high moisture alfalfa silages by regulating bacterial community composition. Front. Microbiol. 2020, 11, 1989. [Google Scholar] [CrossRef]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccha-rides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Menke, K.H.; Steingass, H. Estimation of the energetic feed value obtained from chemical analysis and in vitro gas production using rumen fluid. Anim. Res. Dev. 1988, 28, 7–55. [Google Scholar]
- Chen, L.; Dong, Z.; Li, J.; Shao, T. Ensiling characteristics, in vitro rumen fermentation, microbial communities and aerobic stability of low-dry matter silages produced with sweet sorghum and alfalfa mixtures. J. Sci. Food Agr. 2019, 99, 2140–2151. [Google Scholar] [CrossRef]
- Broderick, G.A.; Kang, J.H. Automated simultaneous determination of ammonia and total amino acids in ruminal fluid and in vitro media. J. Dairy Sci. 1980, 63, 64–75. [Google Scholar] [CrossRef]
- Hu, W.; Schmidt, R.J.; McDonell, E.E.; Klingerman, C.M.; Kung, L.M. The effect of Lactobacillus buchneri 40788 or Lactobacillus plantarum MTD-1 on the fermentation and aerobic stability of corn silages ensiled at two dry matter contents. J. Dairy Sci. 2009, 92, 3907–3914. [Google Scholar] [CrossRef] [Green Version]
- Ke, W.; Ding, Z.; Li, F.; Xu, D.; Bai, J.; Muhammad, I.; Zhang, Y.; Zhao, L.; Guo, X. The effects of malic or citric acid on the fermentation quality, proteolysis and lipolysis of alfalfa silage ensiled at two dry matter contents. J. Anim. Physiol. Anim. Nutr. 2021, 106, 988–994. [Google Scholar] [CrossRef]
- Zhang, Y.; Huang, Z.; Vyas, D.; Adesogan, A.T.; Francoc, M.; Ke, W.; Li, F.; Bai, J.; Ding, Z.; Guo, X. Antioxidant status, chemical composition and fermentation profile of alfalfa silage ensiled at two dry matter contents with a novel Lactobacillus plantarum strain with high-antioxidant activity. Anim. Feed Sci. Technol. 2021, 272, 114751. [Google Scholar] [CrossRef]
- Heron, S.J.E.; Edwards, R.A.; McDonald, P. Changes in the nitrogenous components of gamma-irradiated and inoculated ensiled ryegrass. J. Sci. Food Agric. 1986, 37, 979–985. [Google Scholar] [CrossRef]
- Heron, S.J.E.; Owen, T.R. Review of the effects of ‘Ecosyl’ silage inoculant on in vivo digestibilities and metabolizable energy of grass silages. In Management Issues for the Grassland Farmer in the 1990’s; Mayne, C.S., Ed.; Occasional Symposium No 25; British Grassland Society: Birmingham, UK, 1991; pp. 230–233. [Google Scholar]
- Palarea-Albaladejo, J.; Rooke, J.A.; Nevison, I.M.; Dewhurst, R.J. Compositional mixed modeling of methane emissions and ruminal volatile fatty acids from individual cattle and multiple experiments. J. Anim. Sci. 2017, 95, 2467–2480. [Google Scholar] [CrossRef] [PubMed]
- Oskoueian, E.; Jahromi, M.F.; Jafari, S.; Shakeri, M.; Le, H.H.; Ebrahimi, M. Manipulation of rice straw silage fermentation with different types of lactic acid bacteria inoculant affects rumen microbial fermentation characteristics and methane production. Vet. Sci. 2021, 8, 100. [Google Scholar] [CrossRef]
- Wolin, M.J. Interactions between the bacterial species of the rumen. In Digestion and Metabolism in the Ruminant; McDonald, I.M., Warner, A.C.I., Eds.; University of New England Publishing Unit: Sydney, Australia, 1975; pp. 135–148. [Google Scholar]
- Belaich, J.P.; Bruschi, M.; Garcia, J.L. Microbiology and Biochemistry of Strict Anaerobes Involved in Interspecies Hydrogen Transfer; Belaich, J.P., Bruschi, M., Garcia, J., Eds.; Plenum Public Corporation: New York, NY, USA, 1990. [Google Scholar]
- Stewart, C.S.; Bryant, M.P. The Rumen Bacteria in the Rumen Microbial Ecosystem; Hobson, P.N., Ed.; Elsevier Applied Science: New York, NY, USA, 1988; pp. 21–75. [Google Scholar]
- Baker, S.K. Rumen methanogens, and inhibition of methanogenesis. Aust. J. Agric. Res. 1999, 50, 1293–1298. [Google Scholar] [CrossRef]
- Wolin, M.J.; Miller, T.L. Microbe-microbe interactions. In The Rumen Microbial Ecosystem; Hobson, P.N., Stewart, C.S., Eds.; Springer Science & Business Media: London, UK, 1997; pp. 467–488. [Google Scholar]
- Gamo, Y.; Mii, M.; Zhou, X.G.; Sar, C.; Santoso, B.; Arai, I.; Kimura, K.; Takahashi, J. Effects of lactic acid bacteria, yeasts and galactooligosaccha-ride supplementation on in vitro rumen methane production. In Proceedings of the 1st International Conference on Greenhouse Gases and Animal Agriculture (GGAA), Obihiro, Japan, 7–11 November 2001; Takahashi, J., Young, B.A., Eds.; 2001; pp. 7–11. [Google Scholar]
- Feng, Y. Nutrition of Ruminants; Feng, Y., Ed.; Science Press: Beijing, China, 2004. [Google Scholar]
- Charmley, E. Towards improved silage quality—A review. The Canadian veterinary journal. Can. J. Anim. Sci. 2001, 81, 157–168. [Google Scholar] [CrossRef]


{kind=link}
| Iterms 1 | M | H | Effects 3 | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Control | MTD/1 | ATCC-14917 | LP1-4 | Mean | Control | MTD/1 | ATCC-14917 | LP1-4 | Mean | SEM 2 | A | D | A × D | |
| pH | 6.67 | 6.67 | 6.74 | 6.75 | 6.71 | 6.69 | 6.68 | 6.72 | 6.70 | 6.70 | 0.014 | 0.359 | 0.682 | 0.801 |
| Total gas (mL/g DM) | 103 Ba | 86.6 Bb | 83.5 b | 99.0 a | 93.0 | 124 Aa | 106 Ab | 90.5 c | 92.5 c | 103 | 18.473 | <0.001 | <0.001 | <0.001 |
| CH4 (mL/g DM) | 11.3 a | 7.34 b | 3.53 c | 6.34 Abc | 7.14 | 9.88 a | 7.23 b | 3.60 c | 2.11 Bc | 5.71 | 0.977 | <0.001 | 0.003 | 0.006 |
| CH4-to-total gas (% mL) | 11.0 a | 8.49 Aab | 4.27 c | 7.29 Abc | 7.77 | 8.00 a | 6.85 Bb | 3.92 c | 2.29 Bc | 5.27 | 1.151 | <0.001 | <0.001 | 0.011 |
| IDVMD (g/kg DM) | 65.5 Ab | 70.3 Aa | 71.7 Aa | 69.3 Aa | 69.2 | 52.1 Bb | 54.5 Bb | 63.4 Ba | 61.5 Ba | 57.9 | 2.609 | <0.001 | <0.001 | 0.001 |
| NH3 (mg/100 mL) | 22.9 | 24.3 | 24.8 | 25.1 | 24.2 | 23.2 | 23.1 | 27.5 | 26.2 | 25.0 | 2.344 | 0.006 | 0.230 | 0.277 |
| Total VFA (mM) | 72.0 A | 68.6 A | 67.8 A | 68.0 A | 69.1 | 54.2 Bab | 50.4 Bb | 57.0 Ba | 57.5 Ba | 54.8 | 4.409 | 0.033 | <0.001 | 0.006 |
| Acetate (mM) | 40.7 Aa | 37.5 Aab | 36.9 Ab | 36.4 Ab | 37.9 | 30.8 Ba | 29.2 Bb | 28.7 Bb | 29.2 Bb | 29.5 | 1.032 | <0.001 | <0.001 | 0.171 |
| Propionate (mM) | 10.3 b | 10.7 b | 12.1 ab | 13.2 a | 11.6 | 11.0 b | 11.0 b | 14.9 a | 14.6 a | 12.9 | 1.060 | <0.001 | 0.007 | 0.233 |
| Acetate to propionate | 3.98 Aa | 3.50 Aab | 3.05 Abc | 2.76 Ac | 3.32 | 2.78 Ba | 2.54 Bab | 2.13 Bbc | 1.80 Bc | 2.32 | 0.055 | <0.001 | <0.001 | 0.703 |
| Butyrate (mM) | 15.1 Aa | 12.8 Aab | 12.3 Aab | 11.3 Ac | 12.9 | 8.52 Ba | 5.76 Bc | 7.37 Bb | 7.38 Bb | 7.26 | 0.958 | 0.001 | <0.001 | 0.039 |
| Isobutyrate (mM) | 1.20 | 1.18 A | 1.29 A | 1.44 | 1.28 | 0.66 b | 0.61 Bb | 1.04 Ba | 1.14 a | 0.86 | 0.062 | 0.041 | 0.001 | 0.577 |
| Valerate (mM) | 2.30 a | 2.01 Bab | 1.77 Bb | 1.84 Bb | 1.98 | 2.00 b | 2.62 Aab | 3.16 Aa | 3.23 Aa | 2.75 | 0.045 | 0.027 | <0.001 | <0.001 |
| Isovalerate (mM) | 2.45 | 3.30 A | 3.37 A | 3.90 A | 3.26 | 1.15 b | 1.19 Bb | 1.86 Ba | 1.90 Ba | 1.52 | 0.266 | 0.012 | <0.001 | 0.486 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Z.; Li, F.; Kharazian, Z.A.; Guo, X. Effect of Inoculating Two Bacteriocin-Producing Lactiplantibacillus plantarum Strains at Ensiling on In Vitro Rumen Fermentation and Methane Emissions of Alfalfa Silage with Two Dry Matter Contents. Animals 2023, 13, 384. https://doi.org/10.3390/ani13030384
Li Z, Li F, Kharazian ZA, Guo X. Effect of Inoculating Two Bacteriocin-Producing Lactiplantibacillus plantarum Strains at Ensiling on In Vitro Rumen Fermentation and Methane Emissions of Alfalfa Silage with Two Dry Matter Contents. Animals. 2023; 13(3):384. https://doi.org/10.3390/ani13030384
Chicago/Turabian StyleLi, Ziqian, Fuhou Li, Zohreh Akhavan Kharazian, and Xusheng Guo. 2023. "Effect of Inoculating Two Bacteriocin-Producing Lactiplantibacillus plantarum Strains at Ensiling on In Vitro Rumen Fermentation and Methane Emissions of Alfalfa Silage with Two Dry Matter Contents" Animals 13, no. 3: 384. https://doi.org/10.3390/ani13030384