

Epidemiologic and Genomic Characterizations of Porcine Kobuviruses in Diarrheic and Healthy Pigs
 , ,
, ,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Clinical Sample Collection
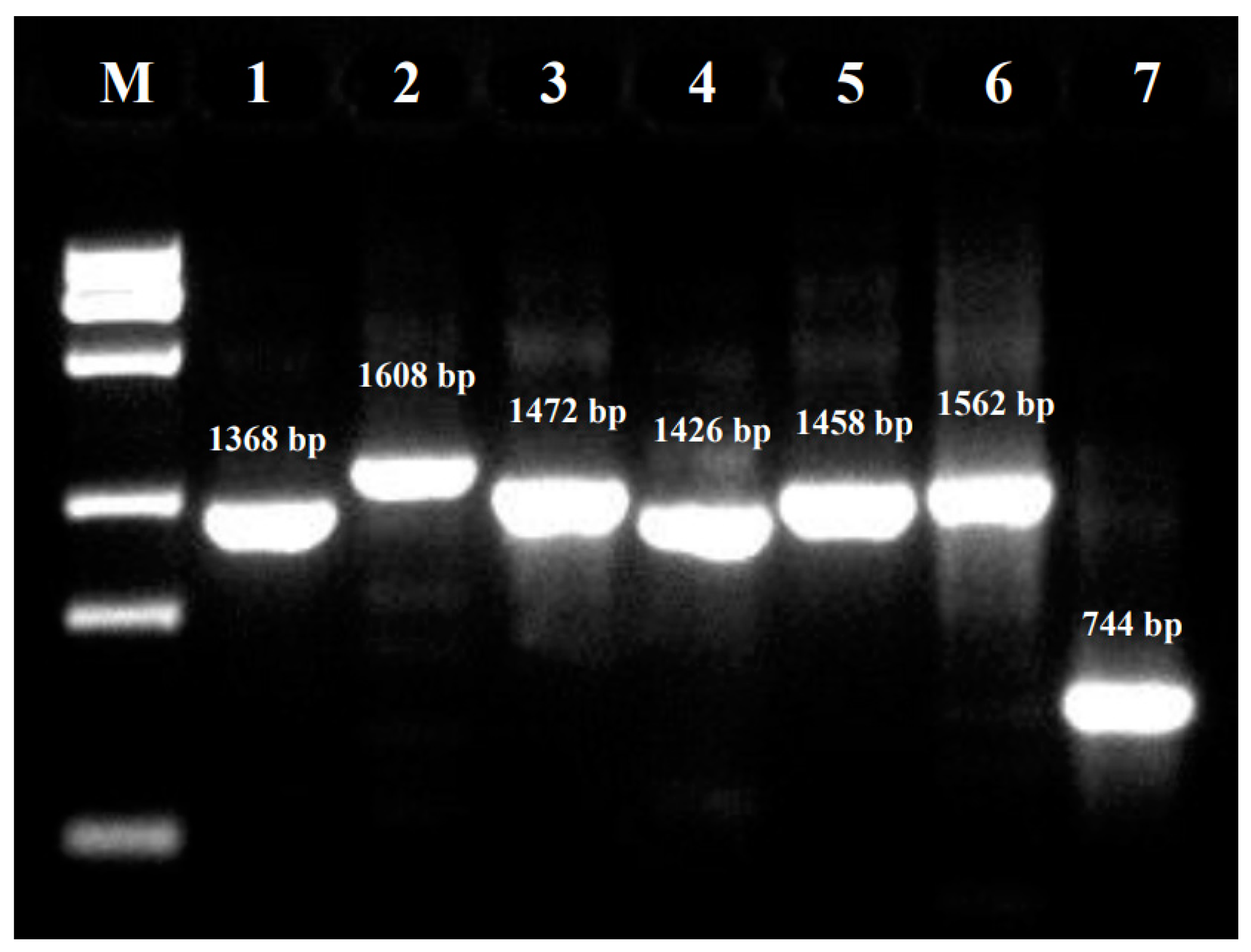
2.2. Real-Time PCR Detection and Genomic Sequencing
2.3. Sequence Alignment and Phylogenetic Analysis
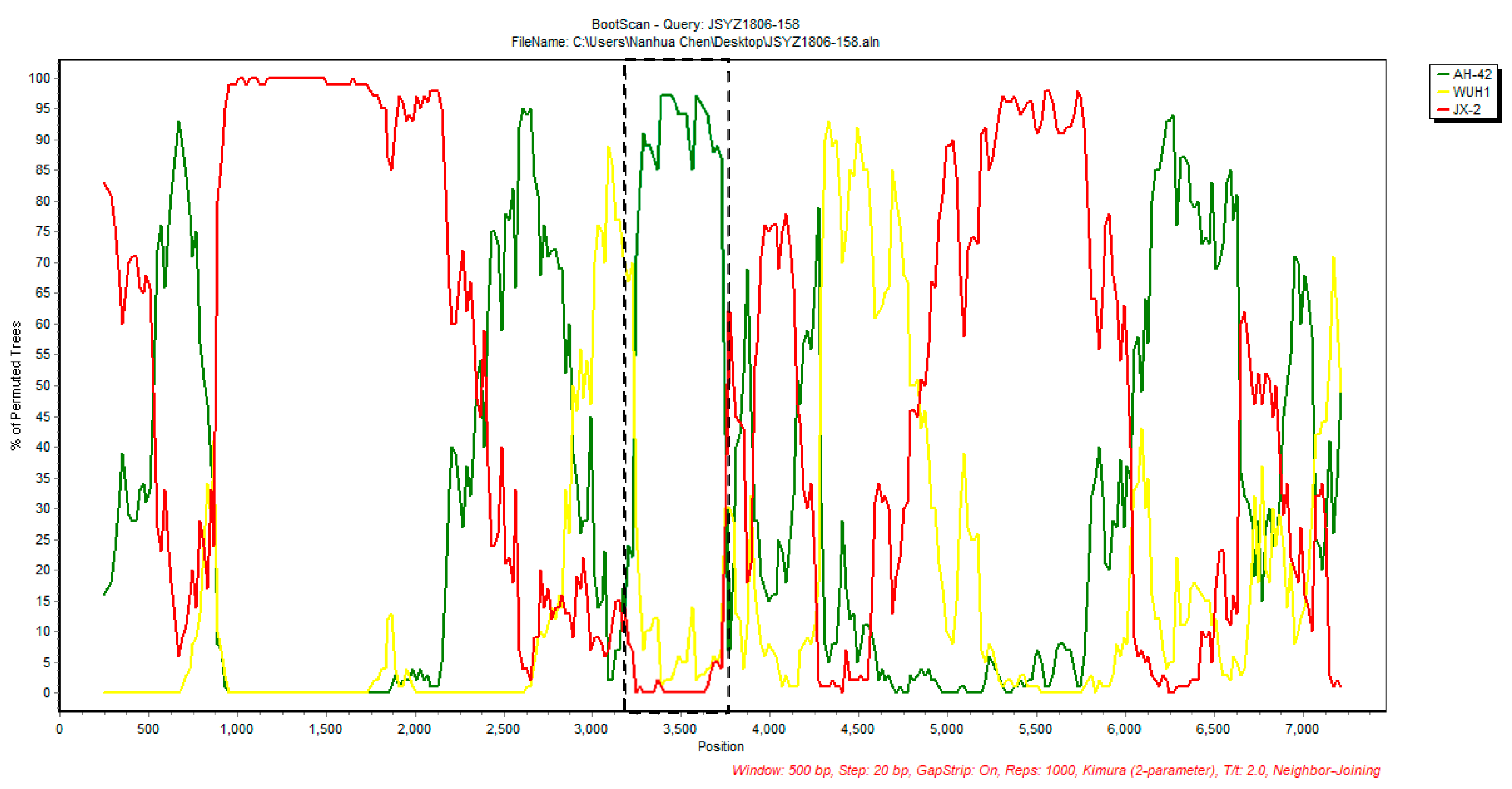
2.4. Recombination Detection and Selective Pressure Analysis
3. Results
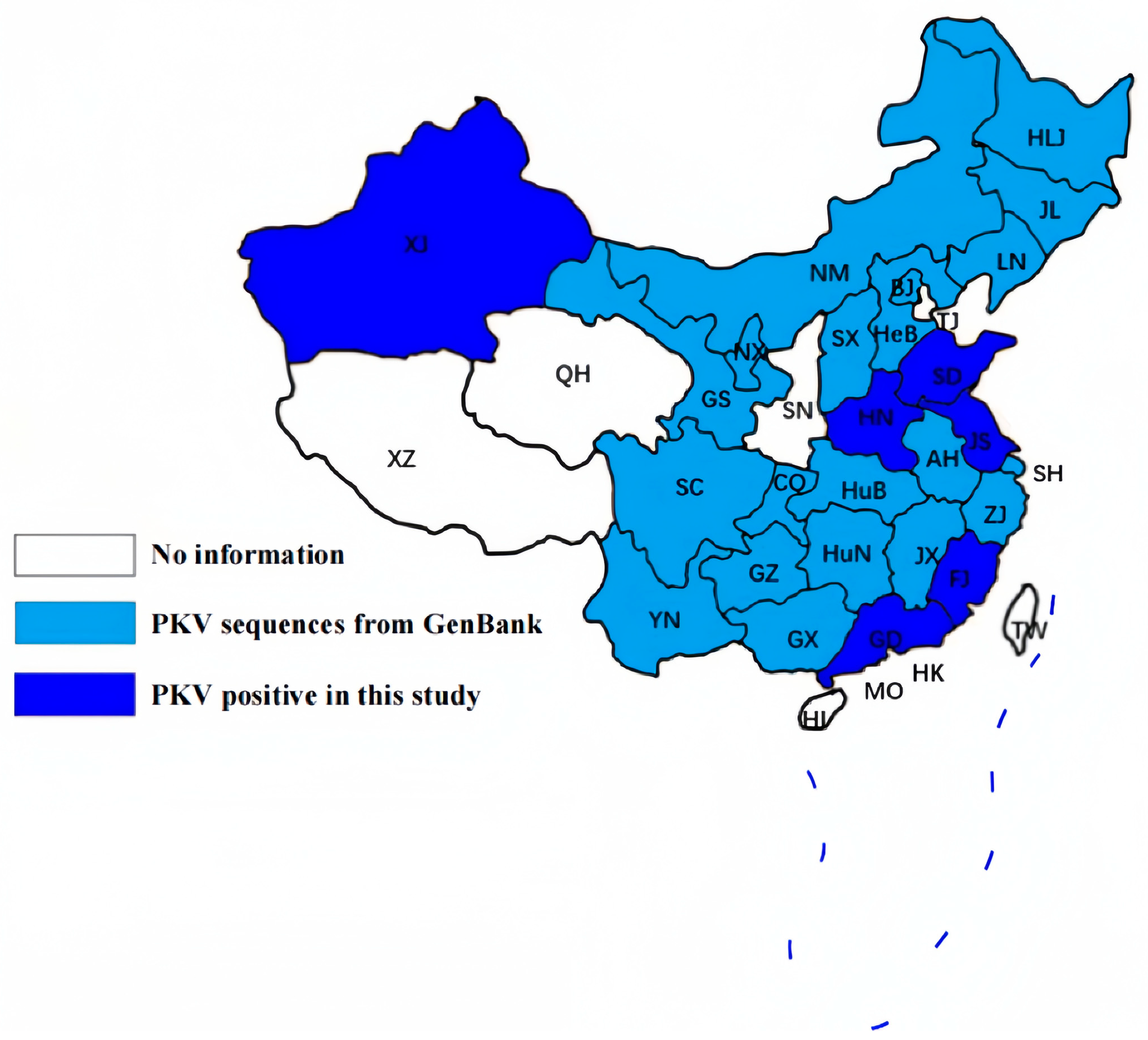
3.1. PKV Is Highly Prevalent in Chinese Swine Herds
3.2. PKV Is Unlikely to Be a Direct Causative Pathogen for Enteric Diseases
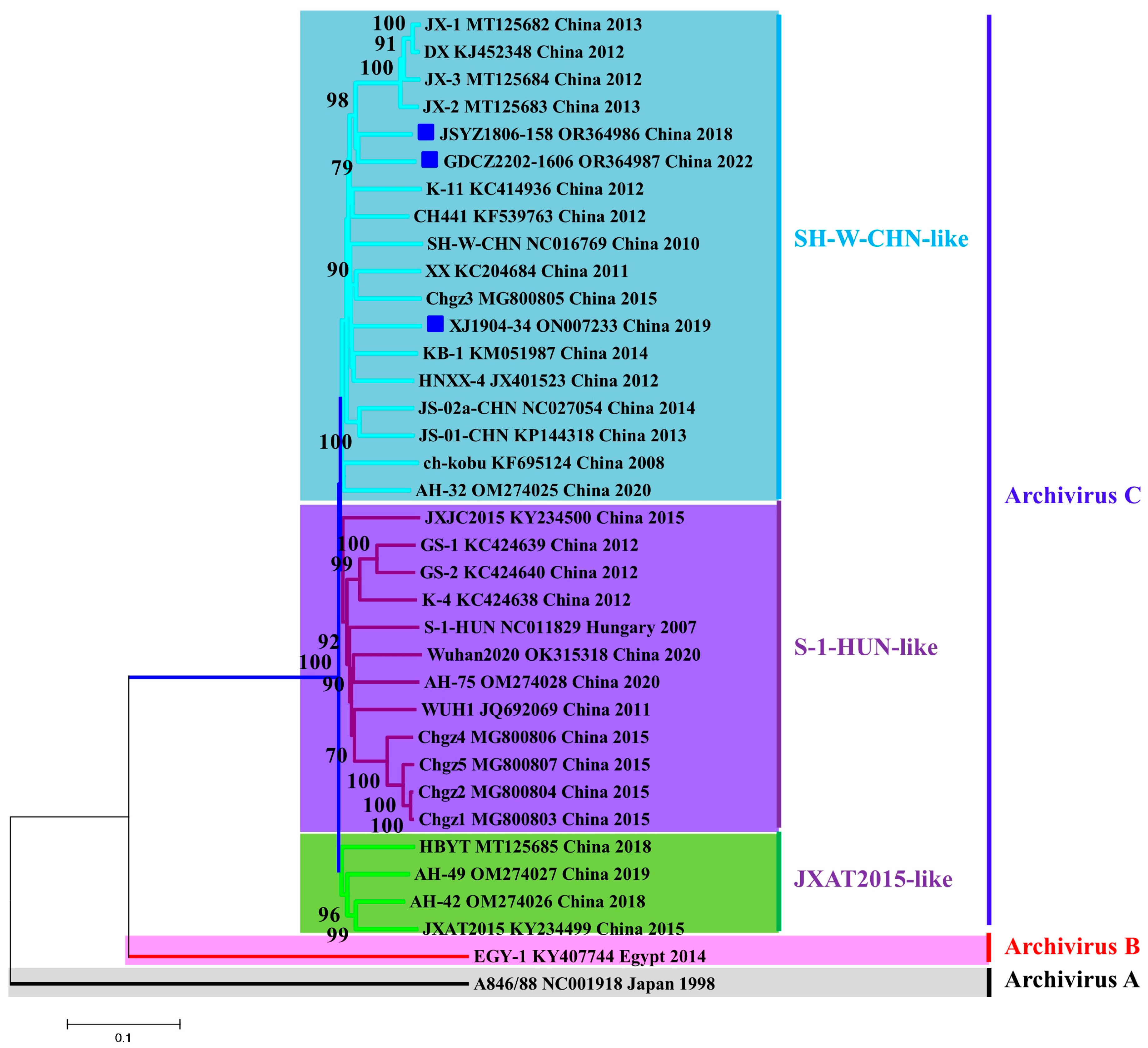
3.3. PKV Strains Are Rapidly Evolving in Chinese Swine Herds
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reuter, G.; Boldizsar, A.; Kiss, I.; Pankovics, P. Candidate new species of Kobuvirus in porcine hosts. Emerg. Infect. Dis. 2008, 14, 1968–1970. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Liang, J.; Wu, S.; Yan, Z.; Zhang, W. Complete genomic sequence analysis and intestinal tissue localization of a porcine Kobuvirus variant in China. Infect. Genet. Evol. 2022, 104, 105362. [Google Scholar] [CrossRef] [PubMed]
- Reuter, G.; Boldizsar, A.; Pankovics, P. Complete nucleotide and amino acid sequences and genetic organization of porcine kobuvirus, a member of a new species in the genus Kobuvirus, family Picornaviridae. Arch. Virol. 2009, 154, 101–108. [Google Scholar] [CrossRef]
- Yamashita, T.; Sakae, K.; Tsuzuki, H.; Suzuki, Y.; Ishikawa, N.; Takeda, N.; Miyamura, T.; Yamazaki, S. Complete nucleotide sequence and genetic organization of Aichi virus, a distinct member of the Picornaviridae associated with acute gastroenteritis in humans. J. Virol. 1998, 72, 8408–8412. [Google Scholar] [CrossRef]
- Reuter, G.; Boros, A.; Pankovics, P. Kobuviruses—A comprehensive review. Rev. Med. Virol. 2011, 21, 32–41. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, T.; Kobayashi, S.; Sakae, K.; Nakata, S.; Chiba, S.; Ishihara, Y.; Isomura, S. Isolation of cytopathic small round viruses with BS-C-1 cells from patients with gastroenteritis. J. Infect. Dis. 1991, 164, 954–957. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, T.; Ito, M.; Kabashima, Y.; Tsuzuki, H.; Fujiura, A.; Sakae, K. Isolation and characterization of a new species of kobuvirus associated with cattle. J. Gen. Virol. 2003, 84 Pt 11, 3069–3077. [Google Scholar] [CrossRef]
- Milicevic, V.; Kureljusic, B.; Maksimovic-Zoric, J.; Savic, B.; Spalevic, L.; Zutic, J. Molecular detection and characterization of Porcine Kobuvirus in domestic pigs and wild boars in Serbia. Res. Vet. Sci. 2020, 132, 404–406. [Google Scholar] [CrossRef]
- Nantel-Fortier, N.; Lachapelle, V.; Letellier, A.; L’Homme, Y.; Brassard, J. Kobuvirus shedding dynamics in a swine production system and their association with diarrhea. Vet. Microbiol. 2019, 235, 319–326. [Google Scholar] [CrossRef]
- Zhai, S.L.; Zhang, H.; Lin, T.; Chen, S.N.; Zhou, X.; Chen, Q.L.; Lv, D.H.; Wen, X.H.; Zhou, X.R.; Jia, C.L.; et al. A novel porcine kobuvirus emerged in piglets with severe diarrhoea in China. Transbound. Emerg. Dis. 2017, 64, 1030–1036. [Google Scholar] [CrossRef]
- Lu, L.; Van Dung, N.; Ivens, A.; Bogaardt, C.; O’Toole, A.; Bryant, J.E.; Carrique-Mas, J.; Van Cuong, N.; Anh, P.H.; Rabaa, M.A.; et al. Genetic diversity and cross-species transmission of kobuviruses in Vietnam. Virus Evol. 2018, 4, vey002. [Google Scholar] [CrossRef] [PubMed]
- Khamrin, P.; Maneekarn, N.; Okitsu, S.; Ushijima, H. Epidemiology of human and animal kobuviruses. Virusdisease 2014, 25, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.M.; Jin, M.; Zhang, Q.; Li, H.Y.; Li, D.D.; Xu, Z.Q.; Li, J.S.; Cui, S.X.; Yang, S.H.; Liu, N.; et al. Candidate porcine Kobuvirus, China. Emerg. Infect. Dis. 2009, 15, 823–825. [Google Scholar] [CrossRef]
- Khamrin, P.; Maneekarn, N.; Kongkaew, A.; Kongkaew, S.; Okitsu, S.; Ushijima, H. Porcine kobuvirus in piglets, Thailand. Emerg. Infect. Dis. 2009, 15, 2075–2076. [Google Scholar] [CrossRef]
- Khamrin, P.; Maneekarn, N.; Hidaka, S.; Kishikawa, S.; Ushijima, K.; Okitsu, S.; Ushijima, H. Molecular detection of kobuvirus sequences in stool samples collected from healthy pigs in Japan. Infect. Genet. Evol. 2010, 10, 950–954. [Google Scholar] [CrossRef] [PubMed]
- An, D.J.; Jeoung, H.Y.; Jeong, W.; Lee, H.S.; Park, J.Y.; Kim, B. Porcine kobuvirus from pig stool in Korea. Virus Genes 2011, 42, 208–211. [Google Scholar] [CrossRef] [PubMed]
- Barry, A.F.; Ribeiro, J.; Alfieri, A.F.; van der Poel, W.H.; Alfieri, A.A. First detection of kobuvirus in farm animals in Brazil and the Netherlands. Infect. Genet. Evol. 2011, 11, 1811–1814. [Google Scholar] [CrossRef] [PubMed]
- Di Profio, F.; Ceci, C.; Di Felice, E.; Marsilio, F.; Di Martino, B. Molecular detection of porcine kobuviruses in Italian swine. Res. Vet. Sci. 2013, 95, 782–785. [Google Scholar] [CrossRef]
- Verma, H.; Mor, S.K.; Abdel-Glil, M.Y.; Goyal, S.M. Identification and molecular characterization of Porcine kobuvirus in U. S. swine. Virus Genes 2013, 46, 551–553. [Google Scholar] [CrossRef]
- Dufkova, L.; Scigalkova, I.; Moutelikova, R.; Malenovska, H.; Prodelalova, J. Genetic diversity of porcine sapoviruses, kobuviruses, and astroviruses in asymptomatic pigs: An emerging new sapovirus GIII genotype. Arch. Virol. 2013, 158, 549–558. [Google Scholar] [CrossRef]
- Amimo, J.O.; Okoth, E.; Junga, J.O.; Ogara, W.O.; Njahira, M.N.; Wang, Q.; Vlasova, A.N.; Saif, L.J.; Djikeng, A. Molecular detection and genetic characterization of kobuviruses and astroviruses in asymptomatic local pigs in East Africa. Arch. Virol. 2014, 159, 1313–1319. [Google Scholar] [CrossRef]
- Van Dung, N.; Anh, P.H.; Van Cuong, N.; Hoa, N.T.; Carrique-Mas, J.; Hien, V.B.; Sharp, C.; Rabaa, M.; Berto, A.; Campbell, J.; et al. Large-scale screening and characterization of enteroviruses and kobuviruses infecting pigs in Vietnam. J. Gen. Virol. 2016, 97, 378–388. [Google Scholar] [CrossRef] [PubMed]
- Jackova, A.; Sliz, I.; Mandelik, R.; Salamunova, S.; Novotny, J.; Kolesarova, M.; Vlasakova, M.; Vilcek, S. Porcine kobuvirus 1 in healthy and diarrheic pigs: Genetic detection and characterization of virus and co-infection with rotavirus A. Infect. Genet. Evol. 2017, 49, 73–77. [Google Scholar] [CrossRef] [PubMed]
- Collins, P.J.; McMenamy, M.J.; McClintock, J.; Lagan-Tregaskis, P.; McCabe, L.; Doherty, S.; O’Shea, H.; Welsh, M.; McKillen, J. Molecular detection of kobuviruses in livestock in Northern Ireland and the Republic of Ireland. Arch. Virol. 2017, 162, 1275–1279. [Google Scholar] [CrossRef] [PubMed]
- Goecke, N.B.; Hjulsager, C.K.; Kongsted, H.; Boye, M.; Rasmussen, S.; Granberg, F.; Fischer, T.K.; Midgley, S.E.; Rasmussen, L.D.; Angen, O.; et al. No evidence of enteric viral involvement in the new neonatal porcine diarrhoea syndrome in Danish pigs. BMC Vet. Res. 2017, 13, 315. [Google Scholar] [CrossRef] [PubMed]
- Theuns, S.; Vanmechelen, B.; Bernaert, Q.; Deboutte, W.; Vandenhole, M.; Beller, L.; Matthijnssens, J.; Maes, P.; Nauwynck, H.J. Nanopore sequencing as a revolutionary diagnostic tool for porcine viral enteric disease complexes identifies porcine kobuvirus as an important enteric virus. Sci. Rep. 2018, 8, 9830. [Google Scholar] [CrossRef]
- Valko, A.; Marosi, A.; Csagola, A.; Farkas, R.; Ronai, Z.; Dan, A. Frequency of diarrhoea-associated viruses in swine of various ages in Hungary. Acta Vet. Hung. 2019, 67, 140–150. [Google Scholar] [CrossRef]
- Garcia-Hernandez, M.E.; Trujillo-Ortega, M.E.; Alcaraz-Estrada, S.L.; Lozano-Aguirre-Beltran, L.; Sandoval-Jaime, C.; Taboada-Ramirez, B.I.; Sarmiento-Silva, R.E. Molecular Detection and Characterization of Porcine Epidemic Diarrhea Virus and Porcine Aichivirus C Coinfection in Mexico. Viruses 2021, 13, 738. [Google Scholar] [CrossRef]
- Malik, Y.S.; Bhat, S.; Sircar, S.; Verma, A.K.; Barman, N.N.; Deka, P.J.; Ghosh, S.; Reuter, G.; Dhama, K. Kobuvirus Detection in the Critically Endangered Pygmy Hog (Porcula salvania), India. J. Zoo Wildl. Med. 2021, 52, 343–347. [Google Scholar] [CrossRef]
- Park, S.J.; Kim, H.K.; Moon, H.J.; Song, D.S.; Rho, S.M.; Han, J.Y.; Nguyen, V.G.; Park, B.K. Molecular detection of porcine kobuviruses in pigs in Korea and their association with diarrhea. Arch. Virol. 2010, 155, 1803–1811. [Google Scholar] [CrossRef]
- Smolak, D.; Salamunova, S.; Jackova, A.; Harsanyova, M.; Budis, J.; Szemes, T.; Vilcek, S. Analysis of RNA virome in rectal swabs of healthy and diarrheic pigs of different age. Comp. Immunol. Microbiol. Infect. Dis. 2022, 90–91, 101892. [Google Scholar]
- Qiu, M.; Li, S.; Xiao, Y.; Li, J.; Zhang, Y.; Li, X.; Feng, B.; Li, C.; Lin, H.; Zhu, J.; et al. Pathogenic and metagenomic evaluations reveal the correlations of porcine epidemic diarrhea virus, porcine kobuvirus and porcine astroviruses with neonatal piglet diarrhea. Microb. Pathog. 2022, 170, 105703. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.; Xiao, Y.; Ye, M.; Li, X.; Li, S.; Xie, N.; Wei, Y.; Wang, J.; Zhu, J. High genetic diversity of Chinese porcine reproductive and respiratory syndrome viruses from 2016 to 2019. Res. Vet. Sci. 2020, 131, 38–42. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.; Ye, M.; Li, S.; Huang, Y.; Zhou, R.; Yu, X.; Tian, K.; Zhu, J. Emergence of a novel highly pathogenic recombinant virus from three lineages of porcine reproductive and respiratory syndrome virus 2 in China 2017. Transbound. Emerg. Dis. 2018, 65, 1775–1785. [Google Scholar] [CrossRef]
- Feng, B.; Li, C.; Qiu, Y.; Qi, W.; Qiu, M.; Li, J.; Lin, H.; Zheng, W.; Zhu, J.; Chen, N. Genomic Characterizations of Porcine Epidemic Diarrhea Viruses (PEDV) in Diarrheic Piglets and Clinically Healthy Adult Pigs from 2019 to 2022 in China. Animals 2023, 13, 1562. [Google Scholar] [CrossRef]
- Chen, N.H.; Trible, B.R.; Kerrigan, M.A.; Tian, K.G.; Rowland, R.R.R. ORF5 of porcine reproductive and respiratory syndrome virus (PRRSV) is a target of diversifying selection as infection progresses from acute infection to virus rebound. Infect. Genet. Evol. 2016, 40, 167–175. [Google Scholar] [CrossRef]
- Pond, S.L.; Frost, S.D. Datamonkey: Rapid detection of selective pressure on individual sites of codon alignments. Bioinformatics 2005, 21, 2531–2533. [Google Scholar] [CrossRef]
- Li, J.X.; Xiao, Y.Z.; Qiu, M.; Li, X.S.; Li, S.B.; Lin, H.; Li, X.D.; Zhu, J.Z.; Chen, N.H. A Systematic Investigation Unveils High Coinfection Status of Porcine Parvovirus Types 1 through 7 in China from 2016 to 2020. Microbiol. Spectr. 2021, 9, e01294-21. [Google Scholar] [CrossRef]
- Yang, F.; Liu, X.; Zhou, Y.; Lyu, W.; Xu, S.; Xu, Z.; Zhu, L. Histopathology of Porcine kobuvirus in Chinese piglets. Virol. Sin. 2015, 30, 396–399. [Google Scholar] [CrossRef]
- Eriksen, E.O. A Systematic Review: Is Porcine Kobuvirus Causing Gastrointestinal Disease in Young Pigs? Vet. Sci. 2023, 10, 286. [Google Scholar] [CrossRef]
- Chen, L.; Zhu, L.; Zhou, Y.C.; Xu, Z.W.; Guo, W.Z.; Yang, W.Y. Molecular and phylogenetic analysis of the porcine kobuvirus VP1 region using infected pigs from Sichuan Province, China. Virol. J. 2013, 10, 281. [Google Scholar] [CrossRef]
- Liu, P.; Li, P.; Lyu, W.; Li, X.; Li, S.; Yang, F.; Huang, J.; Xu, Z.; Zhu, L. Epidemiological study and variation analysis of the porcine kobuvirus 3D gene in Sichuan province, China. Virol. Sin. 2015, 30, 460–463. [Google Scholar] [CrossRef]
- Wang, C.; Lan, X.; Yang, B. Molecular Epidemiological Investigation of Porcine kobuvirus and Its Coinfection Rate with PEDV and SaV in Northwest China. BioMed Res. Int. 2016, 2016, 7590569. [Google Scholar] [CrossRef]
- Werid, G.M.; Ibrahim, Y.M.; Chen, H.; Fu, L.; Wang, Y. Molecular Detection and Genetic Characterization of Potential Zoonotic Swine Enteric Viruses in Northern China. Pathogens 2022, 11, 417. [Google Scholar] [CrossRef]
- Zhou, P.; Fan, H.; Lan, T.; Yang, X.L.; Shi, W.F.; Zhang, W.; Zhu, Y.; Zhang, Y.W.; Xie, Q.M.; Mani, S.; et al. Fatal swine acute diarrhoea syndrome caused by an HKU2-related coronavirus of bat origin. Nature 2018, 556, 255–258. [Google Scholar] [CrossRef]
- Ding, G.; Fu, Y.; Li, B.; Chen, J.; Wang, J.; Yin, B.; Sha, W.; Liu, G. Development of a multiplex RT-PCR for the detection of major diarrhoeal viruses in pig herds in China. Transbound. Emerg. Dis. 2020, 67, 678–685. [Google Scholar] [CrossRef]
- Zhang, Q.; Hu, R.; Tang, X.; Wu, C.; He, Q.; Zhao, Z.; Chen, H.; Wu, B. Occurrence and investigation of enteric viral infections in pigs with diarrhea in China. Arch. Virol. 2013, 158, 1631–1636. [Google Scholar] [CrossRef]
- Cui, Y.; Li, J.; Guo, J.; Pan, Y.; Tong, X.; Liu, C.; Wang, D.; Xu, W.; Shi, Y.; Ji, Y.; et al. Evolutionary Origin, Genetic Recombination, and Phylogeography of Porcine Kobuvirus. Viruses 2023, 15, 240. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence (5′-3′) | Location 1 | Amplicon (bp) |
|---|---|---|---|
| PKV-F1 | TCGGCCCTCTCACCCTCTTTTC | 24-45 2 | 1368 |
| PKV-R1 | GCAGCAGCAGGTTCCCACCA | 1372-1391 | |
| PKV-F2 | AATGGTTGGACTCCTACGGTGAACA | 1222-1246 | 1608 |
| PKV-R2 | TCCAGTGAGTGGATTCATCACCCA | 2806-2829 | |
| PKV-F3 | CGCGGTACTTACACCGTGTGGGA | 2665-2687 | 1472 |
| PKV-R3 | TGGACAACTTCAGGAGGAGCTTCAA | 4112-4136 | |
| PKV-F4 | TACTCTGCTACTAACAACTGCACCCACTTT | 3943-3972 | 1426 |
| PKV-R4 | TGGAAGTGACCACAACTACCTGGGA | 5344-5368 | |
| PKV-F5 | CCGCCCAGAACCTGTTGTGATCTA | 5043-5066 | 1458 |
| PKV-R5 | GCACCACAGAGACCCTGGAAGGT | 6478-6500 | |
| PKV-F6 | TTGACTGGGCGACCCTCCAA | 6236-6255 | 1562 |
| PKV-R6 | TCGAATGTCATCAGGCACAAACCA | 7774-7797 | |
| PKV-F7 | TTTGGCAACGAGACGTATGAGATGATT | 7465-7491 | 744 |
| PKV-R7 | AATACAGAATAGAAAGTAAAGGACAGTCAGGGA | 8176-8208 |
| Year/Location | Sample No. | PKV-Positive No. 1 | PKV-Positive Percentages |
|---|---|---|---|
| Year | |||
| 2018 | 11 | 3 | 27.27% |
| 2019 | 37 | 26 | 70.27% |
| 2020 | 142 | 76 | 53.52% |
| 2021 | 16 | 4 | 25.00% |
| 2022 | 118 | 103 | 87.29% |
| Total | 324 | 212 | 65.43% |
| Location | |||
| Jiangsu | 67 | 43 | 64.18% |
| Xinjiang | 37 | 26 | 70.27% |
| Guangdong | 32 | 19 | 59.38% |
| Henan | 84 | 69 | 82.14% |
| Shandong | 62 | 46 | 74.19% |
| Fujian | 42 | 9 | 21.43% |
| Total | 324 | 212 | 65.43% |
| Infection Status 1 | Diarrheic Piglets <10 Days Old | Pigs without Diarrhea >1 Month Old | Total |
|---|---|---|---|
| PEDV+ | 29 (90.63%) | 3 (9.37%) | 32 |
| PEDV+ PKV+ | 17 (89.47%) | 2 (10.53%) | 19 |
| PEDV− | 35 (11.98%) | 257 (88.01%) | 292 |
| PEDV− PKV+ | 16 (8.29%) | 177 (91.71%) | 193 |
| No. | Name | Region (City, Province) | Date | Sample | Age | Symptom | PEDV |
|---|---|---|---|---|---|---|---|
| 1 | JSYZ1806-158 | Yangzhou, Jiangsu | June 2018 | Intestine | 2 months old | Diarrhea | Negative |
| 2 | XJ1904-34 | Kashi, Xinjiang | April 2019 | Intestine | 5 days old | Diarrhea, death | Positive |
| 3 | GDCZ2202-1606 | Chaozhou, Guangdong | February 2022 | Intestine | Adult | Clinically healthy | Negative |
| Region | Length (nt) | Nucleotide/Amino Acid Identities (%) | |||||
|---|---|---|---|---|---|---|---|
| JSYZ1806-158 vs. SH-W-CHN | XJ1904-34 vs. SH-W-CHN | GDCZ2202-1606 vs. SH-W-CHN | JSYZ1806-158 vs. XJ1904-34 | XJ1904-34 vs. GDCZ2202-1606 | JSYZ1806-158 vs. GDCZ2202-1606 | ||
| L | 585 | 89.23/96.41 | 88.55/94.36 | 89.06/93.85 | 87.35/95.90 | 86.84/94.36 | 89.23/95.90 |
| VP0 | 1098 | 87.81/92.90 | 86.99/92.90 | 86.72/92.08 | 87.80/97.81 | 87.80/97.54 | 88.07/98.63 |
| VP3 | 669 | 85.05/93.72 | 84.16/93.27 | 84.01/92.38 | 86.40/96.86 | 86.55/95.96 | 91.03/98.65 |
| VP1 | 765 | 82.61/90.98 | 86.14/91.76 | 82.09/90.59 | 83.01/92.16 | 84.58/92.55 | 88.76/98.82 |
| Structural (VP0-VP3) | 2532 | 85.51/92.54 | 85.99/92.65 | 84.60/91.71 | 85.98/95.85 | 86.49/95.62 | 89.06/98.70 |
| 2A | 408 | 87.99/91.91 | 89.71/94.12 | 89.71/96.32 | 89.95/90.44 | 89.22/92.65 | 91.42/92.65 |
| 2B | 495/585/495 1 | 74.36/83.08 | 85.30/92.82 | 75.90/83.08 | 76.24/83.59 | 76.41/83.59 | 93.33/100 |
| 2C | 1005 | 91.74/99.10 | 90.45/99.40 | 91.34/97.61 | 91.54/99.40 | 91.24/97.91 | 91.74/97.91 |
| 3A | 270 | 90.37/98.89 | 86.30/90.00 | 92.59/98.89 | 88.52/91.11 | 87.78/91.11 | 93.33/100 |
| 3B | 102 | 91.18/97.06 | 83.33/97.06 | 86.27/97.06 | 86.27/100 | 88.24/100 | 90.20/100 |
| 3C | 576 | 85.94/94.79 | 85.07/94.27 | 90.28/98.44 | 89.06/98.96 | 86.46/94.27 | 87.67/94.79 |
| 3D | 1407 | 93.46/98.72 | 93.11/98.29 | 94.39/98.72 | 92.89/98.50 | 93.46/98.93 | 94.39/99.15 |
| Nonstructural (2A-3D) | 4263/4353/4263 1 | 88.74/95.52 | 89.41/96.34 | 89.92/96.07 | 89.13/95.59 | 88.86/94.97 | 92.28/97.82 |
| Polyprotein | 7380/7470/7380 | 87.69/94.58 | 88.18/94.94 | 88.05/94.42 | 87.93/95.70 | 87.90/95.14 | 90.93/97.97 |
| Test Method | Gene | No. of Codons | dN/dS | Positive Selection | Negative Selection | Criteria |
|---|---|---|---|---|---|---|
| SLAC | Polyprotein 1 | 2489 | 0.182 | 1 | 1134 | p-value threshold of 0.05 |
| FEL | Polyprotein | 2489 | 5 | 1641 | p-value threshold of 0.05 | |
| FUBAR | Polyprotein | 2489 | 1 | 1856 | Posterior probability of 0.95 |
| Recombinant Virus | Parental Virus | Breakpoint 1 | Score for the Seven Detection Methods Embedded in RDP4 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Major | Minor | Region | Begin | End | RDP | GENECONV | BootScan | MaxChi | Chimaera | SiScan | 3Seq | |
| JSYZ1806-158 | JX-2 (MT125683) | AH-42 (OM274026) | 2A | 3175 | 3517 | 2.714 × 10−3 | - 2 | 1.696 × 10−3 | 8.341 × 10−6 | 9.6 × 10−6 | 1.324 × 10−2 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zang, Y.; Feng, B.; Huang, Z.; Zhao, D.; Qi, W.; Qiu, Y.; Qiu, M.; Li, C.; Lin, H.; Zheng, W.; et al. Epidemiologic and Genomic Characterizations of Porcine Kobuviruses in Diarrheic and Healthy Pigs. Animals 2023, 13, 3129. https://doi.org/10.3390/ani13193129
Zang Y, Feng B, Huang Z, Zhao D, Qi W, Qiu Y, Qiu M, Li C, Lin H, Zheng W, et al. Epidemiologic and Genomic Characterizations of Porcine Kobuviruses in Diarrheic and Healthy Pigs. Animals. 2023; 13(19):3129. https://doi.org/10.3390/ani13193129
Chicago/Turabian StyleZang, Yu, Binghui Feng, Zitao Huang, Dashi Zhao, Wenhao Qi, Yuejia Qiu, Ming Qiu, Chen Li, Hong Lin, Wanglong Zheng, and et al. 2023. "Epidemiologic and Genomic Characterizations of Porcine Kobuviruses in Diarrheic and Healthy Pigs" Animals 13, no. 19: 3129. https://doi.org/10.3390/ani13193129