
Nighttime Lighting Influences on the Plankton Feeding and Growth of Juvenile Pacific Bluefin Tuna, Thunnus orientalis

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
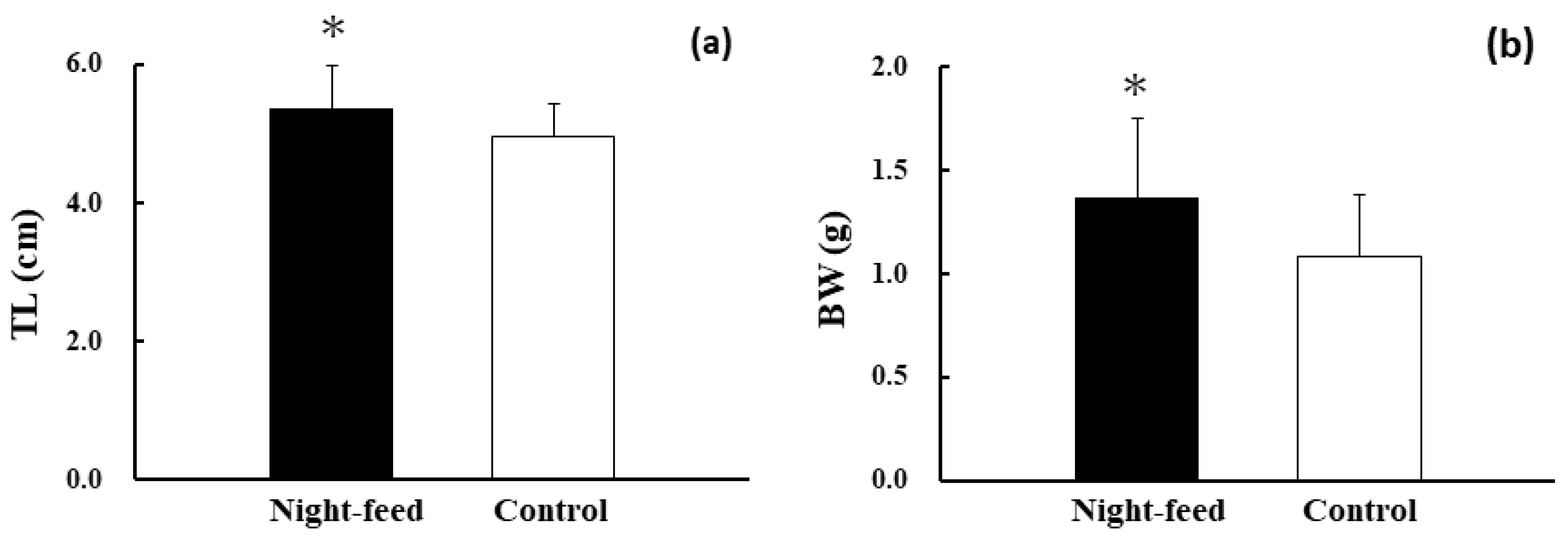
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Harada, T.; Kumai, H.; Mizuno, K.; Murata, O.; Nakamura, M.; Miyashita, S.; Furutani, H. On the rearing of young bluefin tuna. Mem. Fac. Agric. Kinki Univ. 1971, 4, 153–157. [Google Scholar]
- Sawada, Y.; Okada, T.; Miyashita, S.; Murata, O.; Kumai, H. Completion of the Pacific bluefin tuna Thunnus orientalis (Temminck et Schlegel) life cycle. Aquac. Res. 2005, 36, 413–421. [Google Scholar] [CrossRef]
- Benetti, D.D.; Partridge, G.J.; Stieglitz, J. Overview status and technological advances in Tuna aquaculture around the world. In Advances in Tuna Aquaculture from Hatchery to Market; Benetti, D.D., Partridge, G.J., Buentello, A., Eds.; Academic Press: Oxford, UK, 2016; pp. 1–19. [Google Scholar]
- Miyashita, S. Studies on the seedling production of the Pacific bluefin tuna, Thunnus thynnus orientalis. Bull. Fish. Lab. Kinki Univ. 2002, 8, 1–171. [Google Scholar]
- Okada, T.; Honryo, T.; Kawahara, M.; Takahashi, I.; Murayama, K.; Ishibashi, Y. Appropriate size for transportation to sea cages for juvenile Pacific bluefin tuna Thunnus orientalis (Temminck and Schlegel). Aquac. Res. 2021, 52, 1282–1290. [Google Scholar] [CrossRef]
- Higuchi, K.; Tanaka, Y.; Eba, T.; Nishi, A.; Kumon, K.; Nikaido, H.; Shiozawa, S. Causes of heavy mortality of hatchery-reared Pacific bluefin tuna Thunnus orientalis juveniles in sea cages. Aquaculture 2014, 424–425, 140–145. [Google Scholar] [CrossRef]
- Okada, T.; Honryo, T.; Sawada, Y.; Agawa, Y.; Miyashita, S.; Ishibashi, Y. The cause of death of juvenile Pacific bluefin tuna (Thunnus orientalis) reared in sea net cages. Aquac. Eng. 2014, 59, 23–25. [Google Scholar] [CrossRef]
- Okada, T.; Sakurai, Y.; Honryo, T.; Kawahara, M.; Ishibashi, Y. Improving rearing performance in sea-cage culture of Pacific bluefin tuna Thunnus orientalis juveniles (Temminck and Schlegel) using small sea cages. Aquac. Res. 2020, 51, 3017–3024. [Google Scholar] [CrossRef]
- Ishibashi, Y.; Honryo, T.; Saida, K.; Hagiwara, A.; Miyashita, S.; Sawada, Y.; Okada, T.; Kurata, M. Artificial lighting prevents high night-time mortality of juvenile Pacific bluefin tuna, Thunnus orientalis, caused by poor scotopic vision. Aquaculture 2009, 293, 157–163. [Google Scholar] [CrossRef]
- Honryo, T.; Kurata, M.; Okada, T.; Sawada, Y.; Ishibashi, Y. Effect of illuminance transition rate on survival of juvenile Pacific bluefin tuna, Thunnus orientalis. J. World Aquac. Soc. 2014, 45, 523–531. [Google Scholar] [CrossRef]
- Honryo, T.; Okada, T.; Kawahara, M.; Sawada, Y.; Kurata, M.; Ishibashi, Y. Effects of night-time light intensity and area of illumination in the sea cage culture of Pacific bluefin tuna (Thunnus orientalis) juveniles. Aquaculture 2020, 521, 735046. [Google Scholar] [CrossRef]
- Tacon, A.; Rausin, N.; Kadari, M.; Cornelis, P. Use of night-lights to attract food organisms into tropical marine fish cages. Aquac. Res. 1991, 22, 183–192. [Google Scholar] [CrossRef]
- Honryo, T.; Kurata, M.; Okada, T.; Ishibashi, Y. Effects of night-time light intensity on the survival rate and stress responses in juvenile Pacific bluefin tuna Thunnus orientalis (Temminck and Schlegel). Aquac. Res. 2013, 44, 1058–1065. [Google Scholar] [CrossRef]
- Ishibashi, Y.; Honryo, T.; Miyashita, S.; Oda, S. Suitable photoperiod and light intensity of laboratory-reared juvenile Pacific bluefin tuna Thunnus orientalis. Aquac. Sci. 2013, 61, 399–402. [Google Scholar]
- Ishibashi, Y. Fingerling production-II Flexion larvae to juveniles. In Full-Life Cycled Aquaculture of the Pacific Bluefin Tuna; Kumai, H., Miyashita, S., Sakamoto, W., Ono, S., Eds.; Agriculture and Forestry Statistics Publishing: Tokyo, Japan, 2012; pp. 39–59. [Google Scholar]
- Miyashita, S.; Sawada, Y.; Hattori, N.; Nakatsukasa, H.; Okada, T.; Murata, O.; Kumai, H. Mortality of northern bluefin tuna Thunnus thynnus due to trauma caused by collision during growout culture. J. World Aquac. Soc. 2000, 31, 632–639. [Google Scholar] [CrossRef]
- Honryo, T.; Okada, T.; Kurata, M.; Tamura, T.; Ishibashi, Y. Optimal periods of night-time lighting in the sea cage culture of Pacific bluefin tuna, Thunnus orientalis, juvenile (Temminck and Schlegel). Aquac. Res. 2014, 45, 1109–1115. [Google Scholar] [CrossRef]
- Honryo, T.; Okada, T.; Kurata, M.; Ishibashi, Y.; Agawa, Y.; Sawada, Y. Blood chemistry of Pacific bluefin tuna (Thunnus orientalis) juveniles showing abnormal swimming behavior. Aquaculture 2019, 506, 355–358. [Google Scholar] [CrossRef]
- Kawamura, G.; Nishimura, W.; Ueda, S.; Nishi, T. Vision in tunas and marlins. Mem. Kagoshima Univ. Res. Center 1981, 1, 3–26, (In Japanese with English Abstract). [Google Scholar]
- Masuma, S.; Kawamura, G.; Tezuka, N.; Koiso, M.; Namba, K. Retinomotor responses of juvenile bluefin tuna Thunnus thynnus. Fish. Sci. 2001, 67, 228–231. [Google Scholar] [CrossRef]
- Thorson, G. Light as an ecological factor in the dispersal and settlement of larvae of marine bottom invertebrates. Ophelia 1964, 1, 167–208. [Google Scholar] [CrossRef]
- Forward, R.B., Jr. Diel vertical migration: Zooplankton photobiology and behaviour. Oceanogr. Mar. Biol. Annu. Rev. 1988, 26, 361–393. [Google Scholar]



{kind=link}
{kind=link}
| Treatment | Replications | Temperature (°C) | Dissolved Oxygen (%) | pH | Salinity (mg L−1) |
|---|---|---|---|---|---|
| Night-feed | 4 | 27.0 ± 0.1 | 114.8 ± 6.8 | 8.16 ± 0.04 | 32.2 ± 0.2 |
| Control | 4 | 27.0 ± 0.1 | 118.9 ± 5.5 | 8.20 ± 0.02 | 32.1 ± 0.3 |
| Treatment | Day 1 | Day 3 | Day 5 |
|---|---|---|---|
| Night-feed | 77.8% | 69.2% | 75.0% |
| Control | 0% | 0% | 0% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Honryo, T.; Sawada, Y.; Ishibashi, Y. Nighttime Lighting Influences on the Plankton Feeding and Growth of Juvenile Pacific Bluefin Tuna, Thunnus orientalis. Animals 2023, 13, 3128. https://doi.org/10.3390/ani13193128
Honryo T, Sawada Y, Ishibashi Y. Nighttime Lighting Influences on the Plankton Feeding and Growth of Juvenile Pacific Bluefin Tuna, Thunnus orientalis. Animals. 2023; 13(19):3128. https://doi.org/10.3390/ani13193128
Chicago/Turabian StyleHonryo, Tomoki, Yoshifumi Sawada, and Yasunori Ishibashi. 2023. "Nighttime Lighting Influences on the Plankton Feeding and Growth of Juvenile Pacific Bluefin Tuna, Thunnus orientalis" Animals 13, no. 19: 3128. https://doi.org/10.3390/ani13193128